
Adaptation Strategies of Populus euphratica to Arid Environments Based on Leaf Trait Network Analysis in the Mainstream of the Tarim River
Abstract
:1. Introduction
2. Materials and Methods
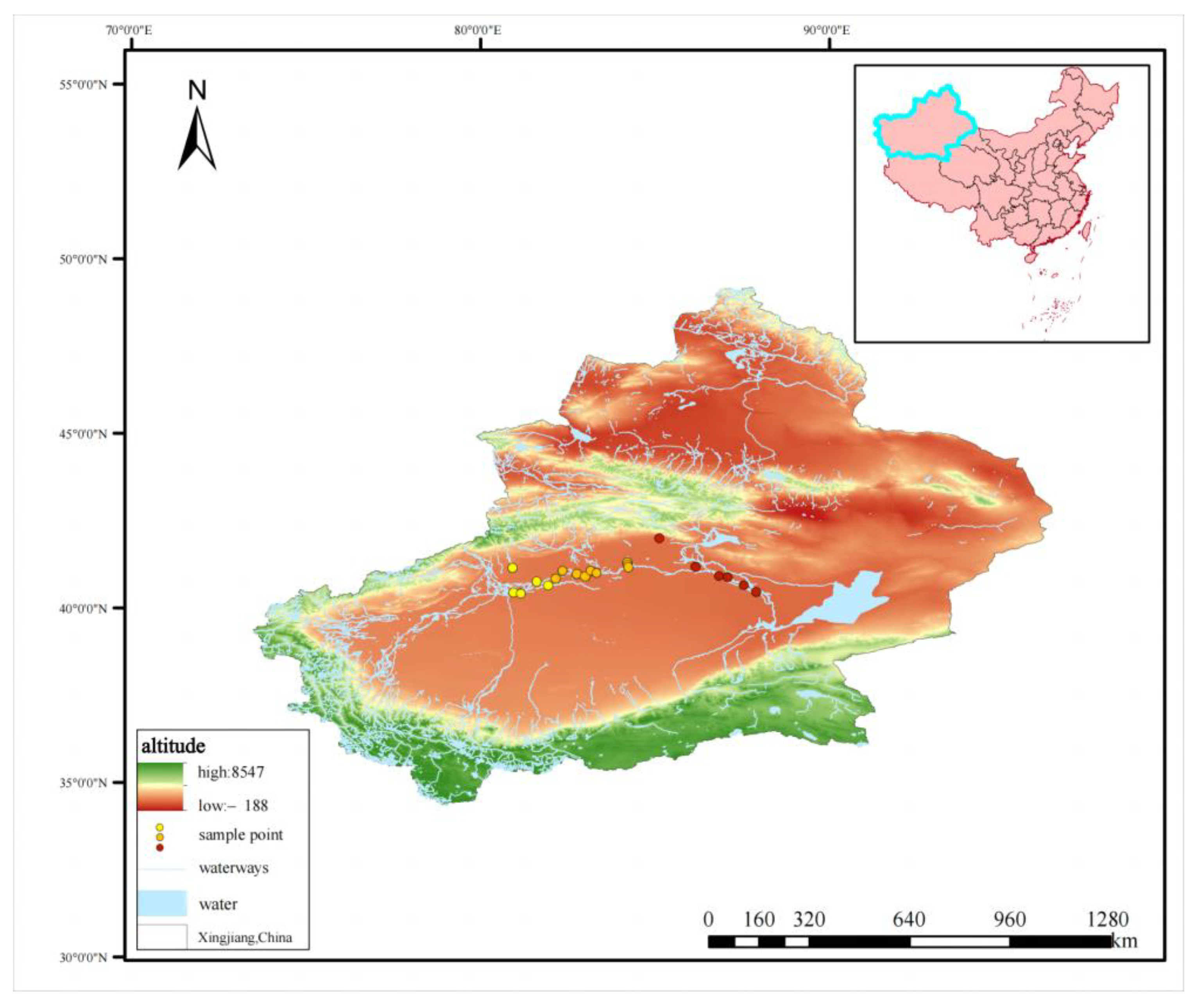
2.1. Study and Sampling Area
2.2. Leaf Sample Collection
2.3. Leaf Measurement and Data Collection
2.4. Establishment of Leaf Trait Networks
2.5. Parameters of Leaf Trait Networks
2.6. Statistical Analyses
3. Results
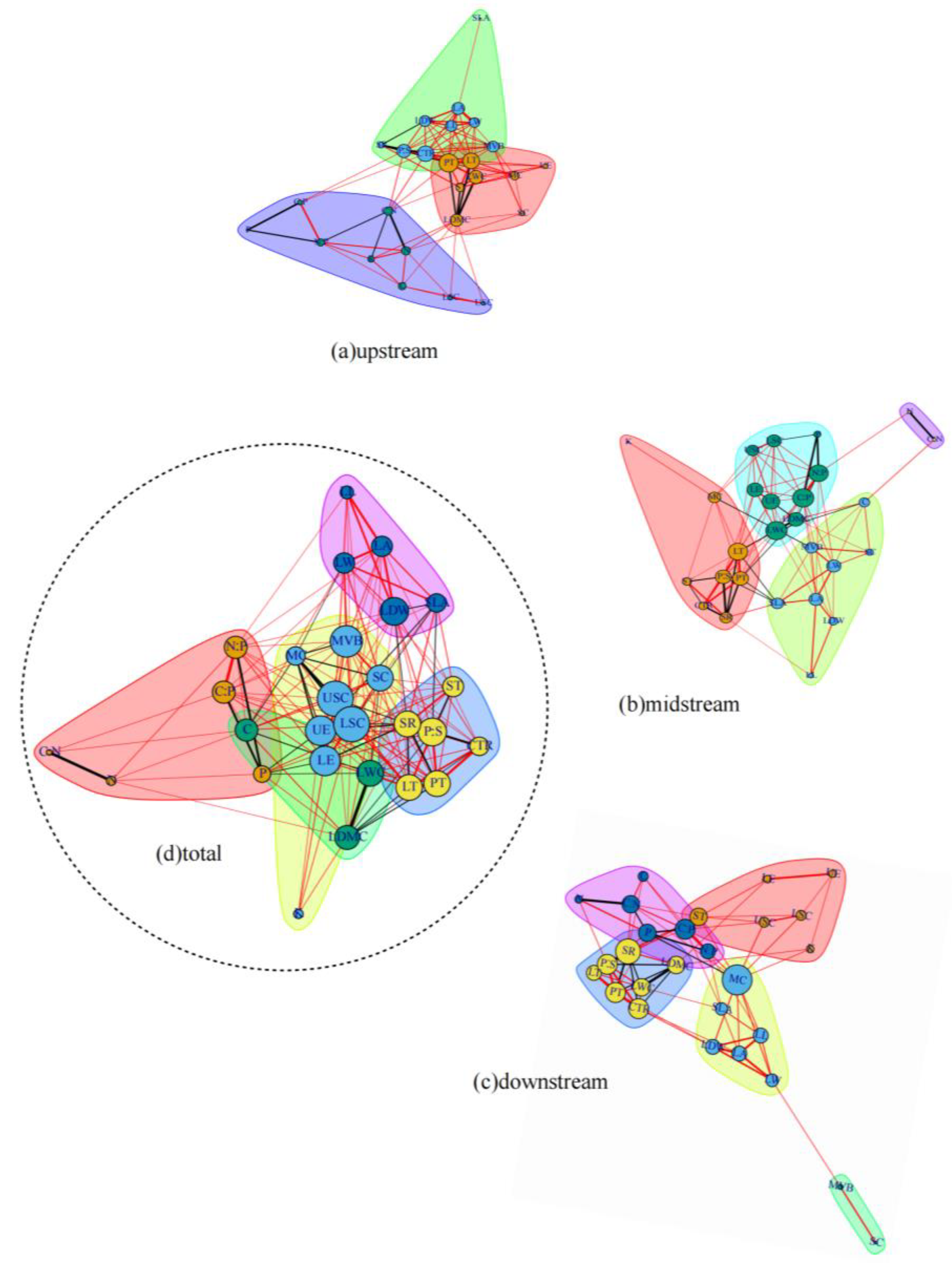
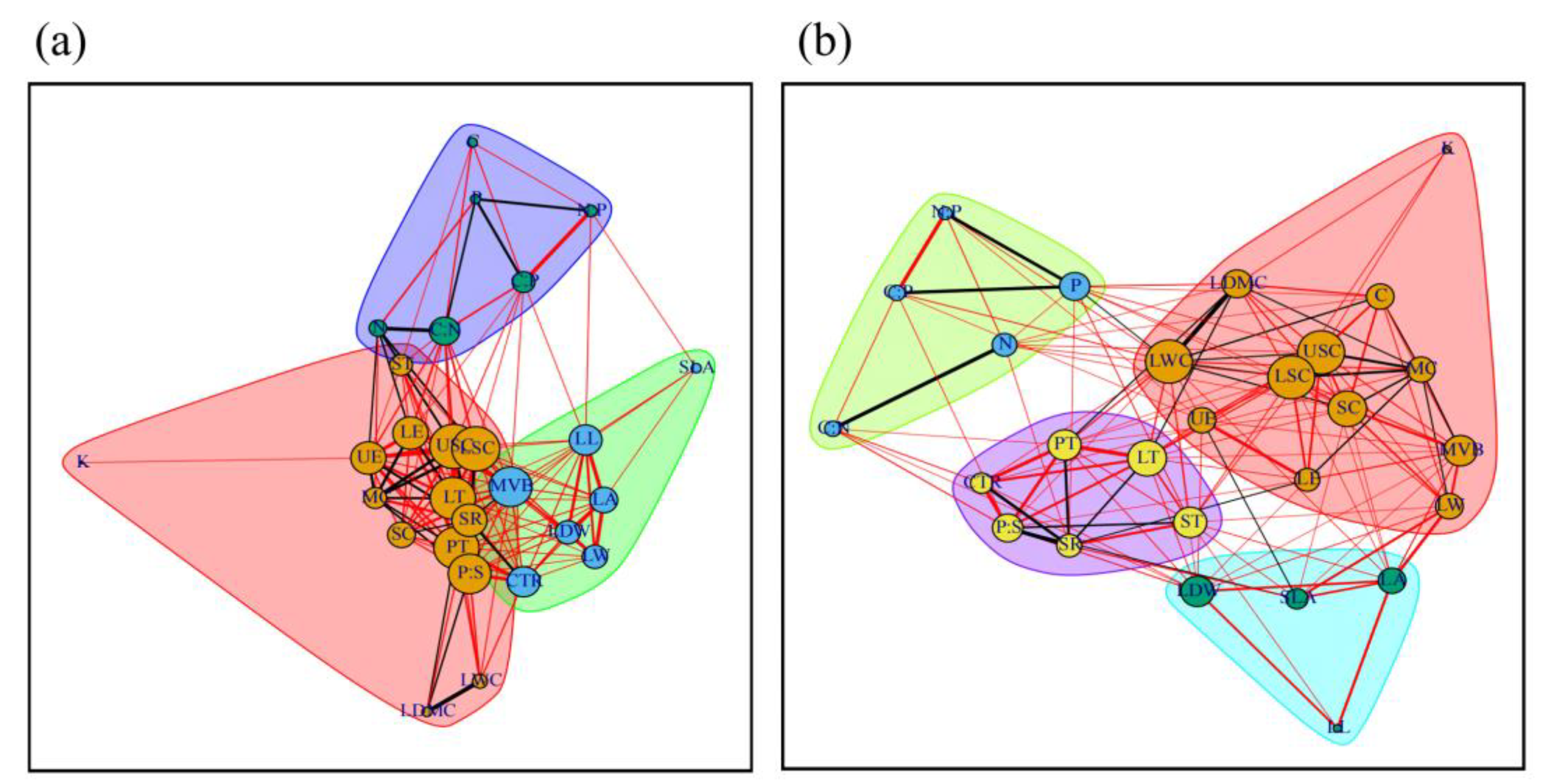
3.1. Leaf Trait Network of Populus euphratica in the Mainstream of Tarim River
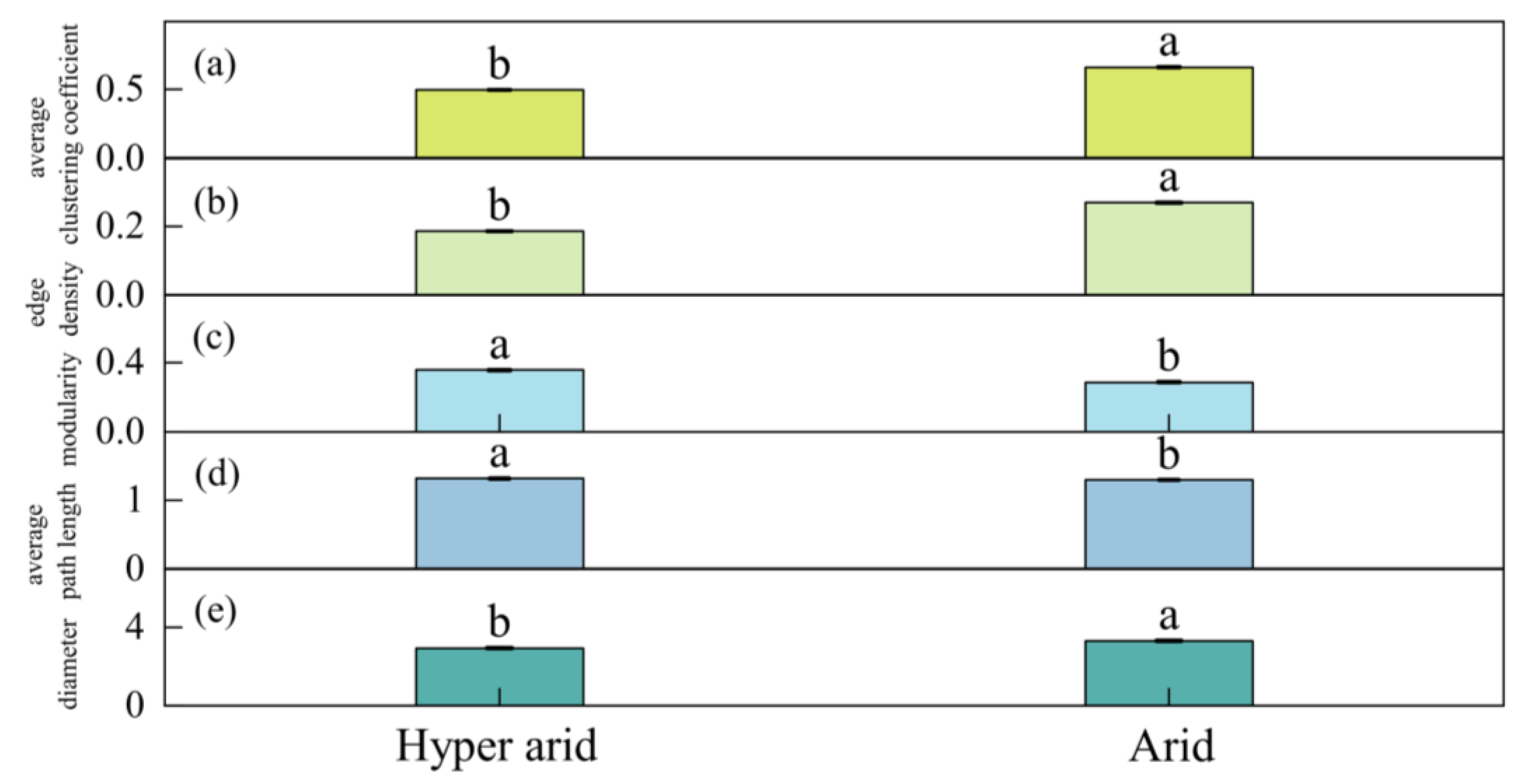
3.2. Differences in Leaf Trait Networks of Populus euphratica in Different Arid Regions along the Mainstream of the Tarim River
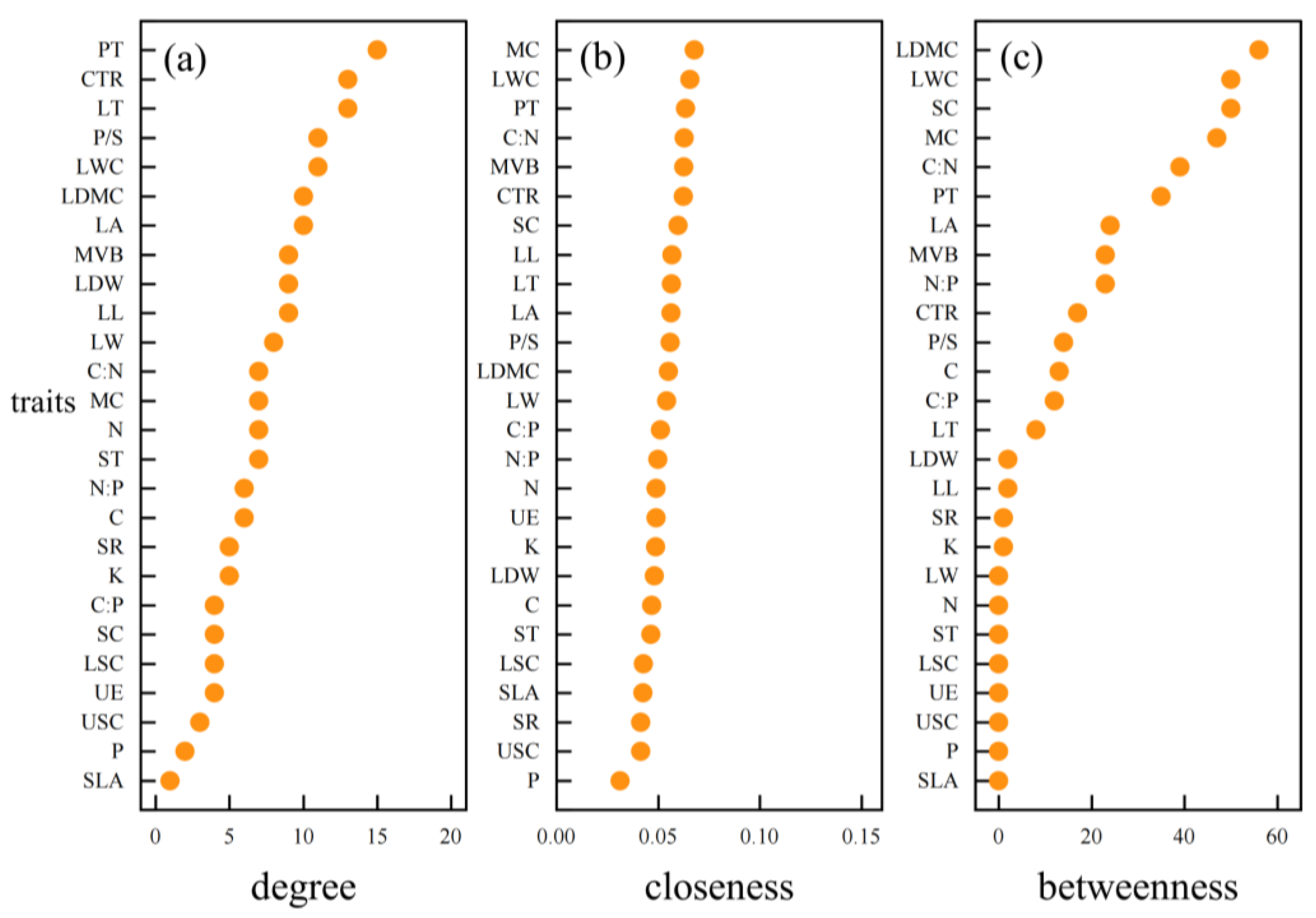
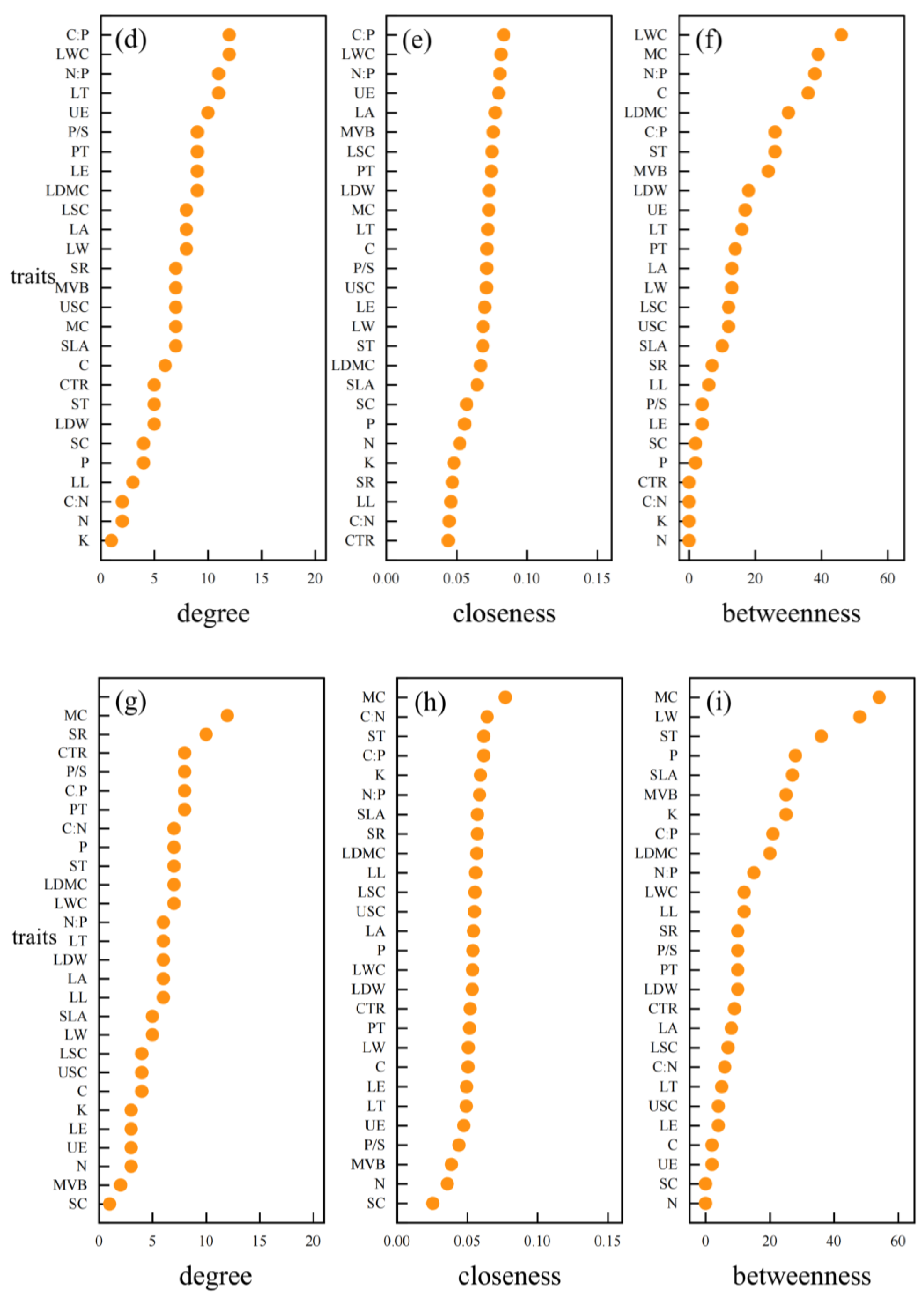
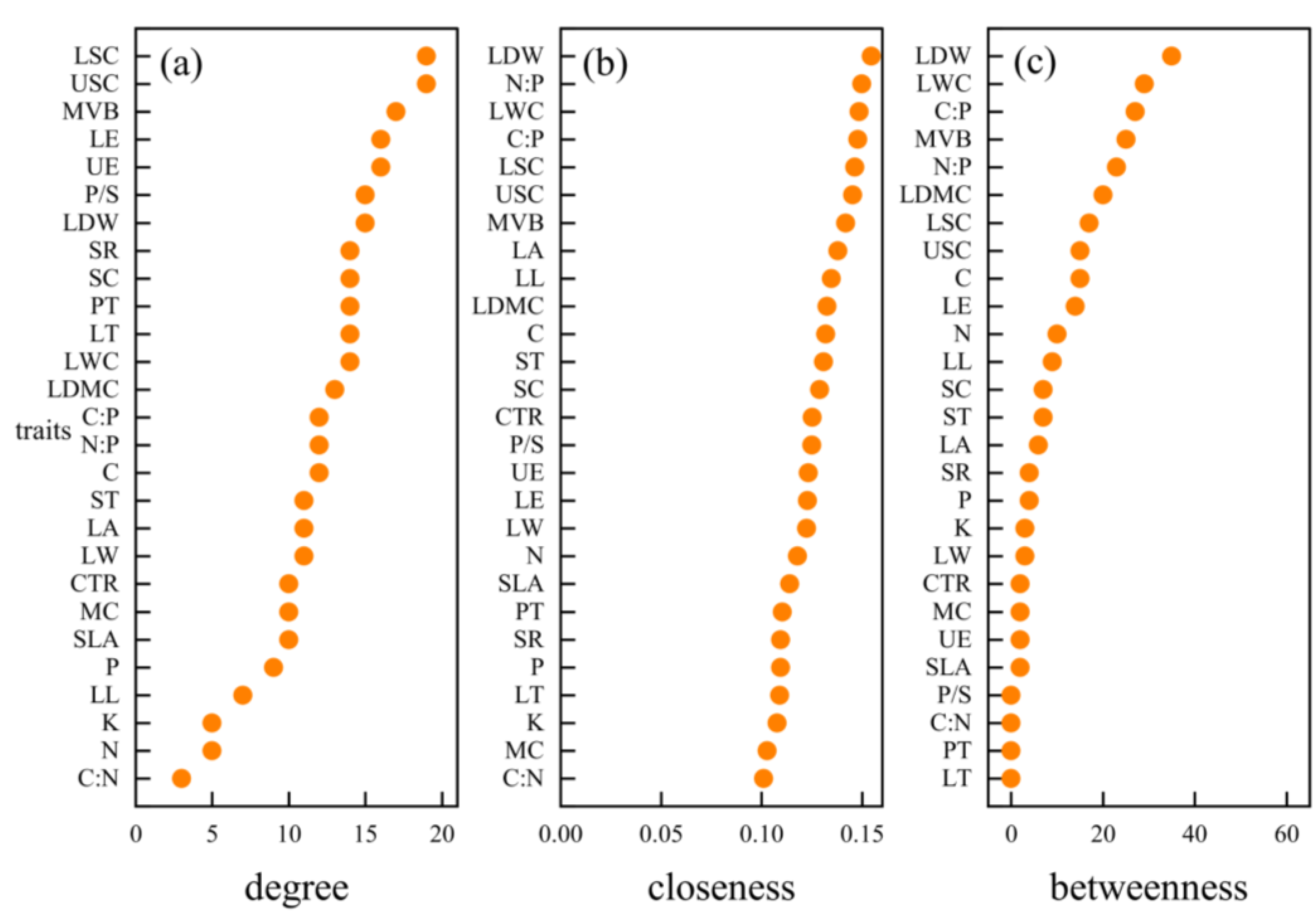
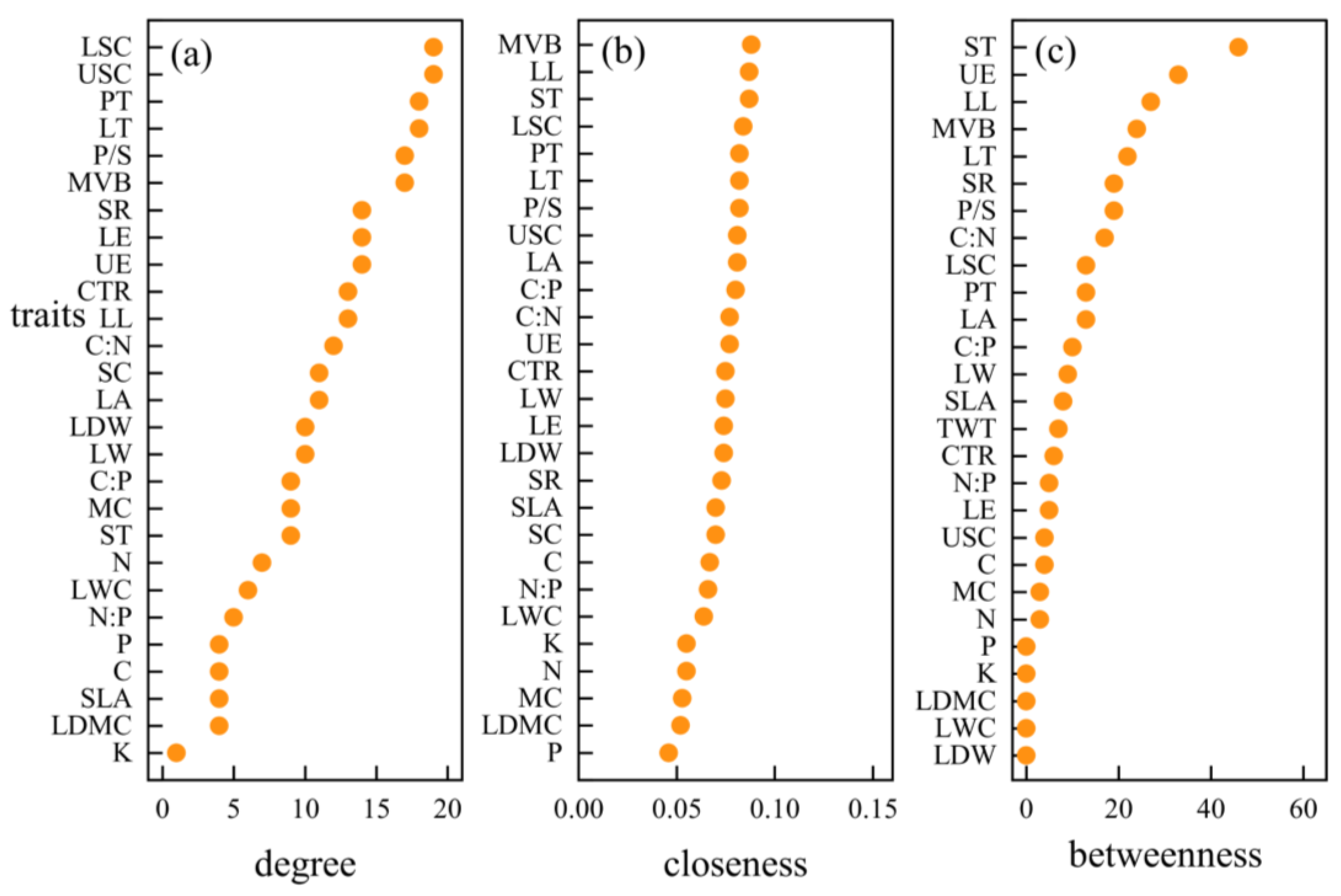
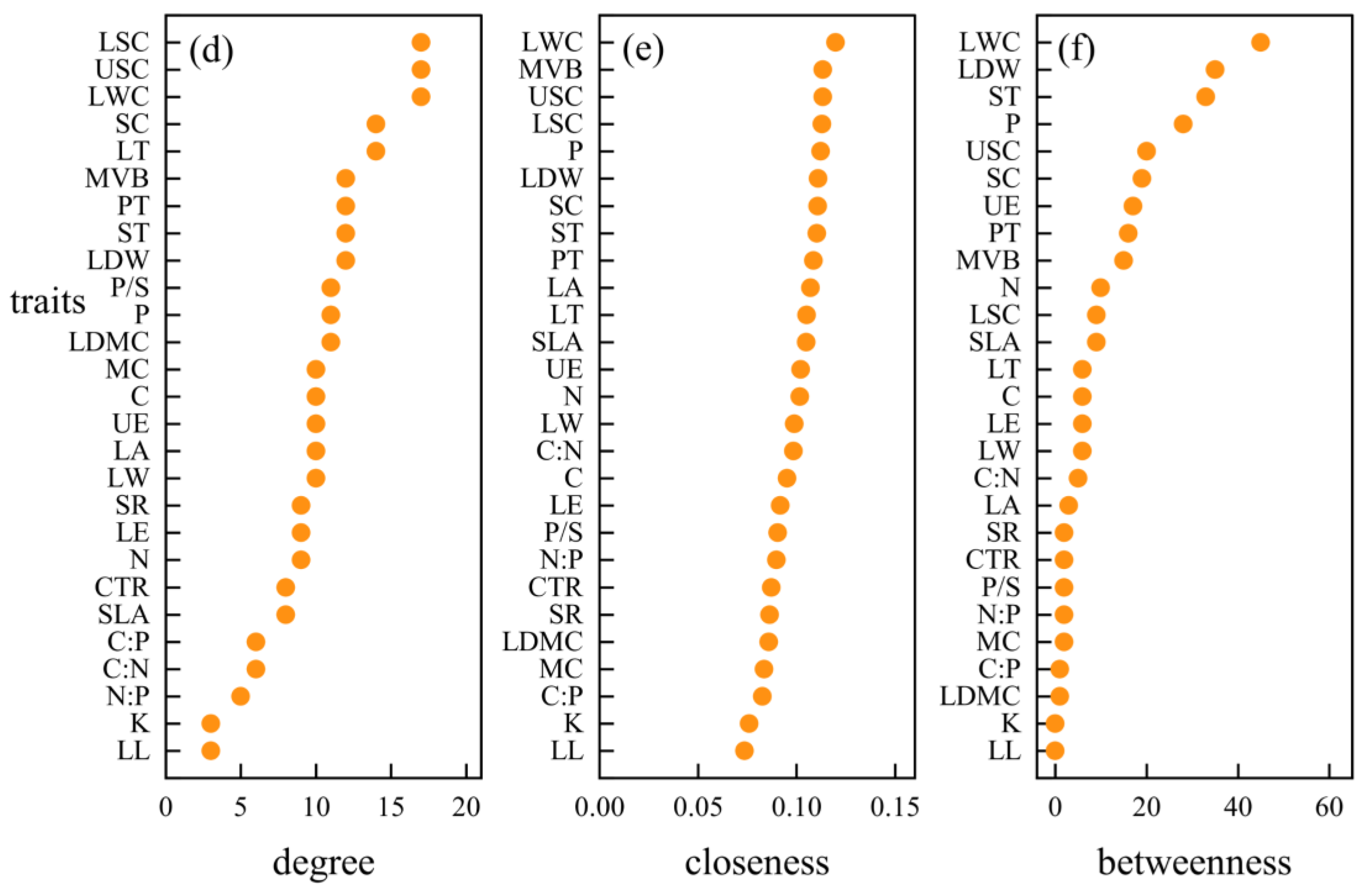
3.3. Hub Traits of Leaf Trait Networks in Different arid Regions
4. Discussion
4.1. Differences and Connections between Leaf Trait Networks of Populus euphratica in the Mainstream of the Tarim River
4.2. Highly Correlated Traits in the Leaf Trait Networks of the Mainstream of the Tarim River
4.3. The Connections and Differences between Leaf Trait Networks in Different Arid Regions
4.4. Important Node Parameters of Leaf Trait Networks in Different arid Regions
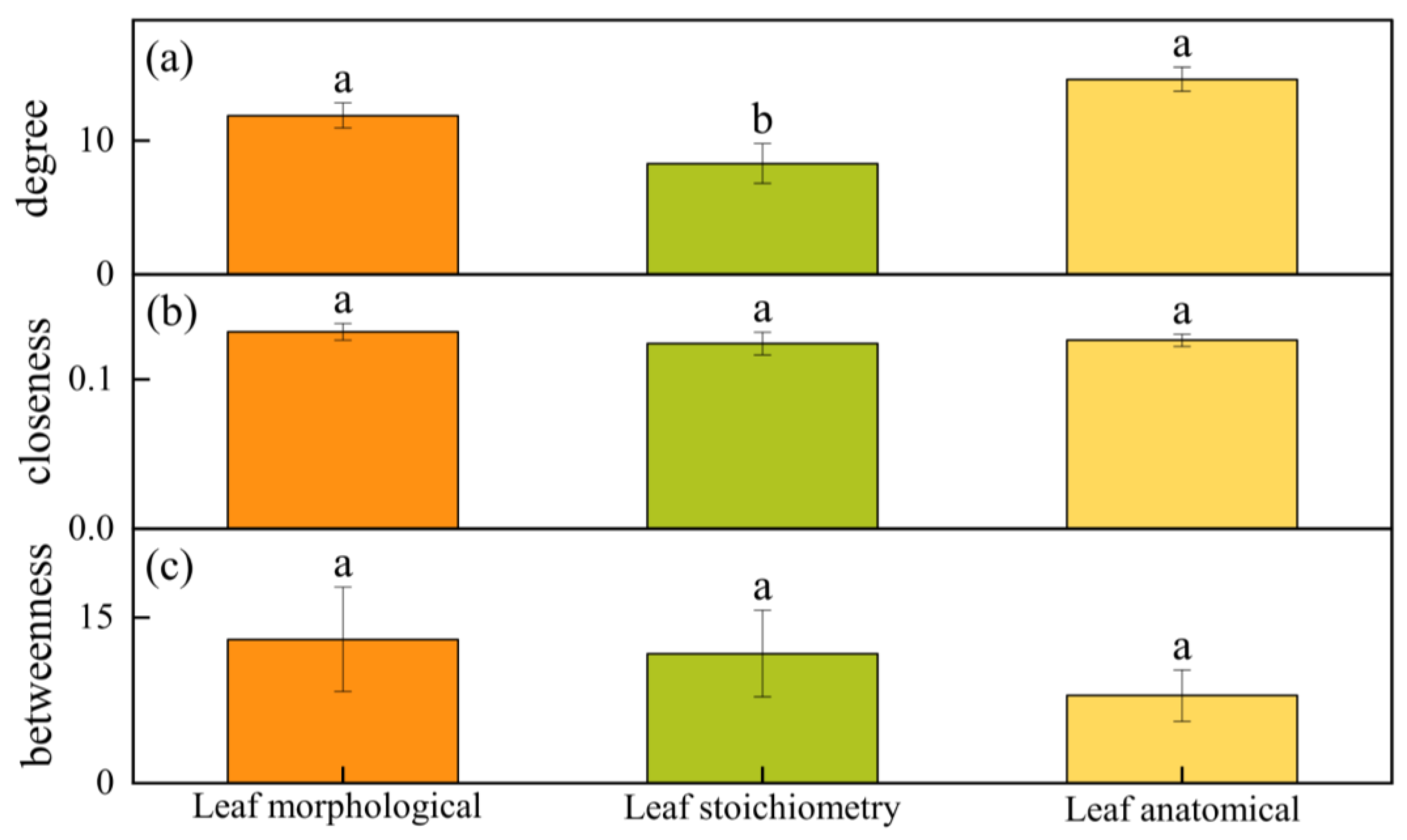
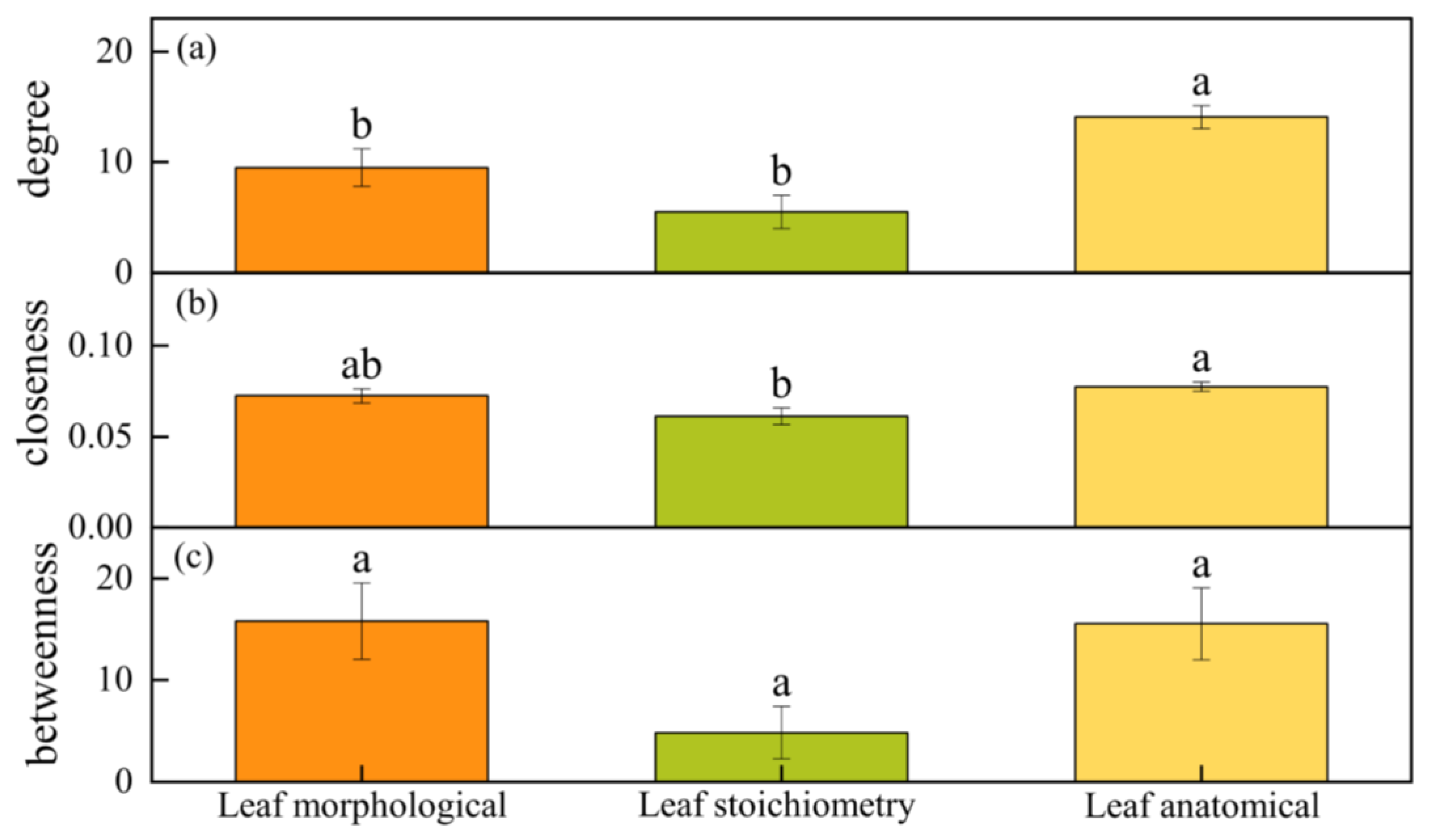
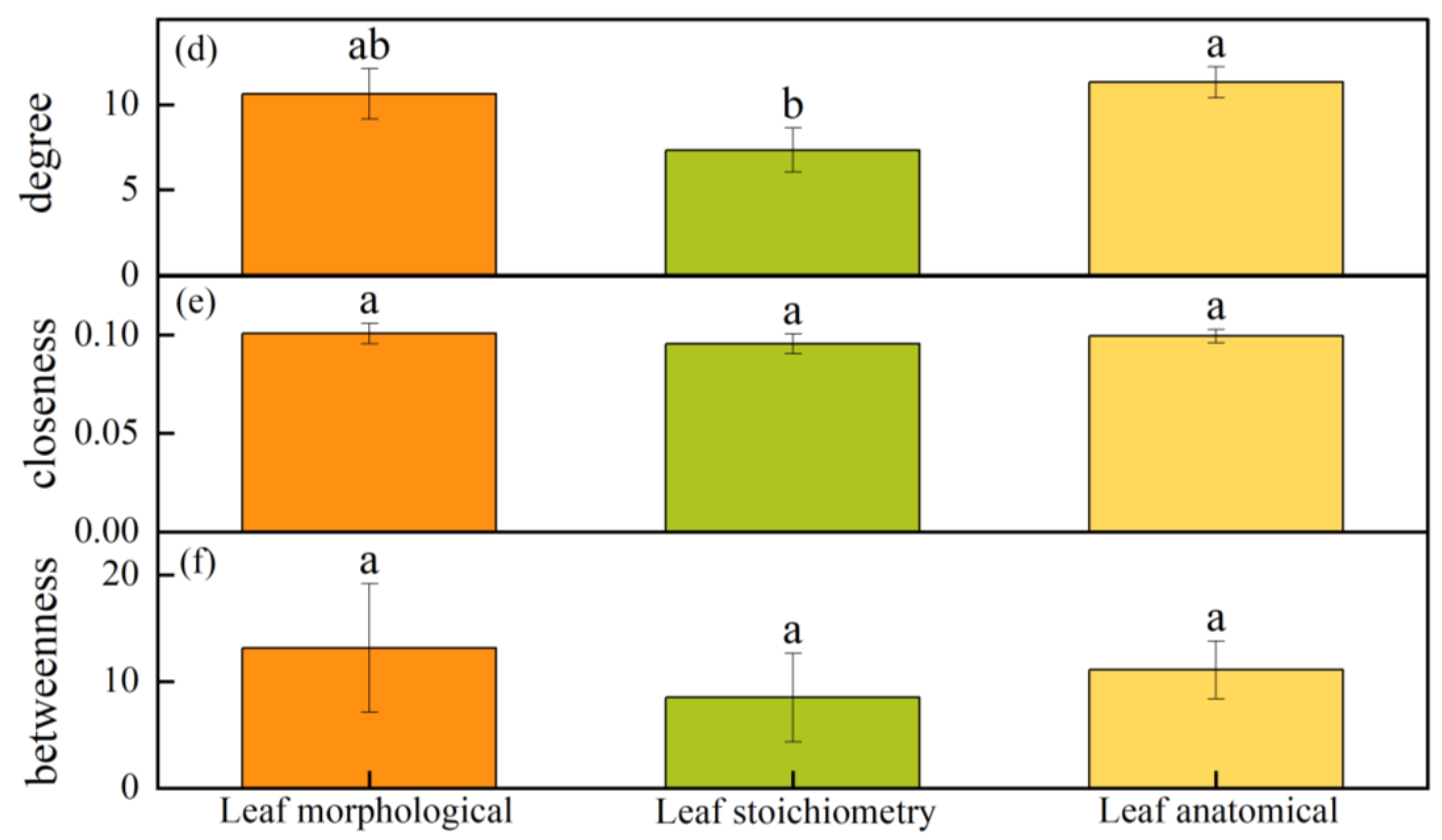
4.5. Comparison of Morphological, Anatomical, and Stoichiometry Traits in the Network Characteristics of Populus euphratica Leaves
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaches | Regions | Sample Point | Longitude/°E | Latitude/°N | Altitude/m |
|---|---|---|---|---|---|
| upstream | arid | U1 | 80.916 | 41.137 | 1084 |
| hyper arid | U2 | 80.944 | 40.437 | 1025 | |
| hyper arid | U3 | 81.155 | 40.425 | 1018 | |
| arid | U4 | 81.6 | 40.757 | 1001 | |
| arid | U5 | 81.929 | 40.654 | 994 | |
| midstream | arid | M1 | 82.152 | 40.845 | 988 |
| arid | M2 | 82.343 | 41.053 | 978 | |
| arid | M3 | 82.743 | 40.963 | 970 | |
| arid | M4 | 83.015 | 40.904 | 962 | |
| arid | M5 | 83.162 | 41.052 | 957 | |
| arid | M6 | 83.316 | 41.006 | 955 | |
| arid | M7 | 84.202 | 41.3 | 932 | |
| arid | M8 | 84.213 | 41.245 | 934 | |
| arid | M9 | 84.238 | 41.172 | 933 | |
| downstream | arid | D1 | 85.127 | 41.994 | 974 |
| hyper arid | D2 | 86.163 | 41.19 | 891 | |
| hyper arid | D3 | 86.843 | 40.924 | 874 | |
| hyper arid | D4 | 87.076 | 40.894 | 868 | |
| hyper arid | D5 | 87.541 | 40.645 | 856 | |
| hyper arid | D6 | 87.904 | 40.454 | 845 |
| Category | Traits | Hyper Arid | Arid | Total |
|---|---|---|---|---|
| Leaf morphology | LL | |||
| LW | ||||
| LA | ||||
| LDW | ||||
| LWC | ||||
| LDMC | ||||
| SLA | ||||
| LT | ||||
| Leaf stoichiometry | N | |||
| K | ||||
| C | ||||
| P | ||||
| N:P | ||||
| C:N | ||||
| C:P | ||||
| Leaf anatomy | ST | |||
| UE | ||||
| LE | ||||
| MC | ||||
| PT | ||||
| USC | ||||
| LSC | ||||
| MVB | ||||
| SC | ||||
| P/S | ||||
| CTR | ||||
| SR |
References
- Diaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional. Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Wang, H.; Harrison, S.P.; Li, M.; Prentice, I.C.; Qiao, S.; Wang, R.; Xu, H.; Mengoli, G.; Peng, Y.; Yang, Y. The China plant trait database version 2. Sci. Data 2022, 9, 769. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, H.; Harrison, S.P.; Prentice, I.C.; Wright, I.J.; Peng, C.; Lin, G. Quantifying leaf-trait covariation and its controls across climates and biomes. New Phytol. 2019, 221, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Cui, E.; Weng, E.; Yan, E.; Xia, J. Robust leaf trait relationships across species under global environmental changes. Nat. Commun. 2020, 11, 2999. [Google Scholar] [CrossRef] [PubMed]
- Rawat, M.; Arunachalam, K.; Arunachalam, A.; Alatalo, J.M.; Pandey, R. Assessment of leaf morphological, physiological, chemical and stoichiometry functional traits for understanding the functioning of Himalayan temperate forest ecosystem. Sci. Rep. 2021, 11, 23807. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Li, Y.; Liu, C.; Xu, L.; Li, M.; Zhang, J.; He, J.; Tang, Z.; Han, X.; Ye, Q.; et al. Plant trait networks: Improved resolution of the dimensionality of adaptation. Trends Ecol. Evol. 2020, 35, 908–918. [Google Scholar] [CrossRef] [PubMed]
- Freschet, G.T.; Kichenin, E.; Wardle, D.A. Explaining within-community variation in plant biomass allocation: A balance between organ biomass and morphology above vs below ground? J. Veg. Sci. 2015, 26, 431–440. [Google Scholar] [CrossRef]
- Niklas, K.J.; Shi, P.; Gielis, J.; Schrader, J.; Niinemets, Ü. Editorial: Leaf functional traits: Ecological and evolutionary implications. Front. Plant Sci. 2023, 14, 1169558. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Lourens, P.; Frans, B. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology 2006, 87, 1733–1743. [Google Scholar]
- Domínguez, M.T.; Aponte, C.; Pérez-Ramos, I.M.; García, L.V.; Villar, R.; Marañón, T. Relationships between leaf morphological traits, nutrient concentrations and isotopic signatures for Mediterranean woody plant species and communities. Plant Soil 2012, 357, 407–424. [Google Scholar] [CrossRef]
- Li, X.; Zhai, J.; Li, Z. Morphological and physiological differences in heteromorphic leaves of male and female Populus euphratica Oliv. J. Arid Land 2022, 14, 1456–1469. [Google Scholar] [CrossRef]
- Zhai, J.; Li, Z.; Si, J.; Zhang, S.; Han, X.; Chen, X. Structural and Functional Responses of the Heteromorphic Leaves of Different Tree Heights on Populus euphratica Oliv. to Different Soil Moisture Conditions. Plants 2022, 11, 2376. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, C.; Xu, L.; Li, M.; Zhang, J.; He, N. Leaf trait networks based on global data: Representing variation and adaptation in plants. Front. Plant Sci. 2021, 12, 710530. [Google Scholar] [CrossRef] [PubMed]
- Kleyer, M.; Trinogga, J.; Cebrianpiqueras, M.A.; Trenkamp, A.; Flojgaard, C.; Ejrnæs, R.; Bouma, T.J.; Minden, V.; Maier, M.; Mantilla-Contreras, J.; et al. Trait correlation network analysis identifies biomass allocation traits and stem specific length as hub traits in herbaceous perennial plants. J. Ecol. 2019, 107, 828–842. [Google Scholar] [CrossRef]
- Rao, Q.; Su, H.; Ruan, L.; Deng, X.; Wang, L.; Rao, X.; Liu, J.; Xia, W.; Xu, P.; Shen, H.; et al. Stoichiometric and physiological mechanisms that link hub traits of submerged macrophytes with ecosystem structure and functioning. Water Res. 2021, 202, 117392. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Rao, Q.; Su, H.; Ruan, L.; Deng, X.; Liu, J.; Chen, J.; Xie, P. Linking the network topology of plant traits with community structure, functioning, and adaptive strategies of submerged macrophytes. Sci. Total Environ. 2022, 850, 158092. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yan, X.X.; Kong, M.; Tong, Q.; Li, Y.; Huang, G.; Li, Y. Industry-environment system management based on an uncertain Gaussian diffusion optimization model for coal-dependent cities in ecologically fragile areas. J. Clean. Prod. 2022, 234, 832–857. [Google Scholar] [CrossRef]
- Cui, C.; Guo, Y.; Shen, Y.J. Spatio-temporal variation in and the driving factors of desert vegetation in Xinjiang. Chin. J. Eco-Agric. 2021, 29, 1668–1678. [Google Scholar]
- Cao, D.; Li, J.; Huang, Z.; Baskin, C.C.; Baskin, J.M.; Hao, P.; Zhou, W.; Li, J. Reproductive characteristics of a Populus euphratica population and prospects for its restoration in China. PLoS ONE 2012, 7, e39121. [Google Scholar] [CrossRef]
- Yang, L.W.; He, B.Y.; Huang, P.Y. Assessment of ecological service values for native Populus euphratica forest in Khotan watershed. Acta Ecol. Sin. 2006, 26, 681–689. [Google Scholar]
- Huang, W.J.; Li, Z.J.; Yang, Z.P.; Bai, G.Z. The structural traits of Populus euphratica heteromorphic leaves and their correlations. Acta Ecol. Sin. 2010, 30, 4636–4642. [Google Scholar]
- Zhai, J.; Zhang, X.; Li, Z.; Han, X.; Zhang, S. Differences in the Functional Traits of Populus pruinosa Leaves in Different Developmental Stages. Plants 2023, 12, 2262. [Google Scholar] [CrossRef] [PubMed]
- Fu, A.; Chen, Y.; Li, W. Assessment on ecosystem health in the Tarim River Basin. Acta Ecol. Sin. 2009, 29, 2418–2426. [Google Scholar]
- Chen, Y.N.; Li, W.H.; Chen, Y.P.; Xu, C. Water conveyance in dried-up river way and ecological restoration in the lower reaches of Tarim river, China. Shengtai Xuebao Acta Ecol. Sin. 2007, 27, 0538–0554. [Google Scholar]
- Nelson, P.W.; Sommers, C.; Page, A.L. Methods of Soil Analysis: Part 2. Chemical Methods; Soil Science Society of America: Madison, WI, USA, 1996. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.; Sumner, M. Methods of Soil Analysis: Part 3—Chemical Methods; Soil Science Society of America: Madison, WI, USA, 1996. [Google Scholar]
- Armbruster, W.S.; Pelabon, C.; Bolstad, G.H.; Hansen, T.F. Integrated phenotypes: Understanding trait covariation in plants and animals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130245. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, C.D.; Scoffoni, C.; John, G.P.; Bartlett, M.K.; Inman-Narahari, F.; Ostertag, R.; Cordell, S.; Giardina, C.; Sack, L. An extensive suite of functional traits distinguishes Hawaiian wet and dry forests and enables prediction of species vital rates. Funct. Ecol. 2019, 33, 712–734. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, Y.H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Zomer, R.; Trabucco, A. Source Code for: Global Aridity Index and Potential Evapotranspiration Database v3—(Python). 2022. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwiemtfhysCEAxU_m1YBHRRcBvsQFnoECA8QAQ&url=https%3A%2F%2Fwww.plantplus.cn%2Fcn%2Fdatasetexport%2Fexportxlxs%2F1512D26417FF6A38&usg=AOvVaw36jK8VFr09i1FdE3j_dEI9&opi=89978449 (accessed on 21 October 2023).
- Trabucco, A.; Zomer, R.J. Global Aridity Index and Potential Evapotranspiration (ET0) Climate Database v2 (Global_AI_PET_v2). 2019. Available online: https://figshare.com/articles/dataset/Global_Aridity_Index_and_Potential_Evapotranspiration_ET0_Climate_Database_v2/7504448/3 (accessed on 21 October 2023).
- Trabucco, A.; Zomer, R.J. Global Aridity Index and PET Database v1 (Global_AI_PET_v1). 2008. Available online: https://cgiarcsi.community/data/global-aridity-and-pet-database/ (accessed on 21 October 2023).
- Ling, H.; Xu, H.; Fu, J. Changes in intra-annual runoff and its response to climate change and human activities in the headstream areas of the Tarim River Basin China. Quat. Int. 2014, 336, 158–170. [Google Scholar] [CrossRef]
- Zhao, R.; Chen, Y.; Li, W.; Chen, Z.; Hao, X. Trends in Runoff Variations of the Mainstream of the Tarim River during the Last 50 Years. Resour. Sci. 2010, 32, 1196–1203. [Google Scholar]
- Yaning, C.; Weihong, L.; Changchun, X.; Zhaoxia, Y.; Yapeng, C. Desert riparian vegetation and groundwater in the lower reaches of the Tarim River basin. Environ. Earth Sci. 2015, 73, 547–558. [Google Scholar]
- Koschützki, D.; Schreiber, F. Centrality analysis methods for biological networks and their application to gene regulatory networks. Gene Regul. Syst. Biol. 2008, 2, 193–201. [Google Scholar]
- Yang, S.D.; Zheng, W.J.; Chen, G.C.; Zhang, C.L.; Chen, J.; Wang, X.C. Difference of Ultrastructure and Photosynthetic Characteristics between Lanceolate and Broad-ovate Leaves in Populus euphratica. Acta Bot. Boreali-Occident. Sin 2005, 25, 14–21. [Google Scholar]
- Zhang, X.; Zheng, Y. Variation in leaf carbon, nitrogen, and phosphorus stoichiometric characteristics of Populus euphratica along a vertical riverbank gradient and associated environmental explanation in a desert riparian area in China. Chin. J. Appl. Environ. Biol. 2023, 29, 1093–1099. [Google Scholar]
- Shi, J.-H.; Wang, X.-Y.; Liu, M.-X.; Ma, X.-X. Stoichiometric Characteristics of Leaves of Populus euphratica with Different Stand Ages and Soil. Arid Zone Res. 2017, 34, 815–822. [Google Scholar]
- Ji, R.-X.; Yu, X.; Chang, Y.; Shen, C.; Bai, X.-Q.; Xia, X.-L.; Yin, W.-L.; Liu, C. Geographical provenance variation of leaf anatomical structure of Caryopteris mongholica and its significance in response to environmental changes. Chin. J. Plant Ecol. 2020, 44, 277–286. [Google Scholar] [CrossRef]
- Kofidis, G.; Bosabalidis, A.M.; Chartzoulakis, K. Leaf anatomical alterations induced by drought stress in two avocado cultivars. J. Biol. Res. 2004, 1, 115–120. [Google Scholar]
- Huang, W.; Jiao, P.; Huang, J.; Zhang, D. Leaf Anatomical Structure of Populus euphratica in Tarim River Basin. Bull. Bot. Res. 2016, 36, 669–675. [Google Scholar]
- Dong, F.; Wang, W.; Cui, P.; Wang, J.; Zhang, T.; Li, J. Plasticity Response of Leaf Anatomical Characteristics of Populus euphratica in Different Soil Conditions. Acta Bot. Boreali-Occident. Sin. 2016, 36, 2047–2057. [Google Scholar]
- Haase, P.; Pugnaire, F.I.; Clark, S.C.; Incoll, L.D. Environmental control of canopy dynamics and photosynthetic rate in the evergreen tussock grass Stipa tenacissima. Plant Ecol. 1999, 145, 327–339. [Google Scholar] [CrossRef]
- Raich, J.W.; Rastetter, E.B.; Melillo, J.M.; Kicklighter, D.W.; Steudler, P.A.; Peterson, B.J.; Grace, A.L.; Moore, B.; Vorosmarty, C.J. Potential net primary productivity in South America: Application of a global model. Ecol. Appl. 1991, 1, 399–429. [Google Scholar] [CrossRef]
- Li, S.J.; Su, P.X.; Zhang, H.N.; Zhou, Z.J.; Xie, T.T. Characteristics and Relationships of Foliar Water and Leaf Functional Traits of Desert Plants. Plant Physiol. 2013, 49, 153–160. [Google Scholar]
- Li, Y.; Liu, C.; Sack, L.; Xu, L.; Li, M.; Zhang, J.; He, N. Leaf trait network architecture shifts with species-richness and climate across forests at continental scale. Ecol. Lett. 2022, 25, 1442–1457. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.J.; Zhang, X.S. Some observations of the adaptations of sandy shrubs to the arid environment in the Mu Us sandland: Leaf water relations and anatomic features. J. Arid Environ. 2001, 48, 41–48. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Li, L.; Wang, H.S.; Dong, J.G. Selecting of the plants constructing energy forest on loess plateau—Part I. Observation on the anatomic feature of 19 xero-morphic plants leaves through lens. Acta Conserv. Soli Aquae Sin. 1989, 3, 20–30. [Google Scholar]
- Yang, Y.N. Comparative study on anatomical structure of leaf of Magnolia sieboldii under different ecological habitats. J. Jilin Agric. Univ. 2010, 32, 476–482. [Google Scholar]
- Lambers, H.; Oliveira, R. Plant Physiological Ecology; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Castro-díez, P.; Puyravaud, J.P.; Cornelissen, J.H.C. Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types. Oecologia 2000, 124, 476–486. [Google Scholar] [CrossRef]
- Hsiao, T.C. Plant Responses to Water Stress. Annu. Rev. Plant Physiol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stress; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Pistón, N.; Bello, F.; Dias, A.T.C.; Götzenberger, L.; Rosado, B.H.P.; Mattos, E.A.; Salguero-Gómez, R.; Carmona, C.P. Multidimensional ecological analyses demonstrate how interactions between functional traits shape fitness and life history strategies. J. Ecol. 2019, 107, 2317–2328. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, S.; Wang, J.; Huang, W.; Jiao, P.; Peng, C.; Li, Y.; Song, S. Adaptation Strategies of Populus euphratica to Arid Environments Based on Leaf Trait Network Analysis in the Mainstream of the Tarim River. Forests 2024, 15, 437. https://doi.org/10.3390/f15030437
Yao S, Wang J, Huang W, Jiao P, Peng C, Li Y, Song S. Adaptation Strategies of Populus euphratica to Arid Environments Based on Leaf Trait Network Analysis in the Mainstream of the Tarim River. Forests. 2024; 15(3):437. https://doi.org/10.3390/f15030437
Chicago/Turabian StyleYao, Shiyu, Jie Wang, Wenjuan Huang, Peipei Jiao, Chengzhi Peng, Ying Li, and Shuangfei Song. 2024. "Adaptation Strategies of Populus euphratica to Arid Environments Based on Leaf Trait Network Analysis in the Mainstream of the Tarim River" Forests 15, no. 3: 437. https://doi.org/10.3390/f15030437