
Is the Fine Root Tensile Strength Predictable from Structural and Morphological Traits across Mycorrhizal Types in Cool-Temperate Woody Species?

Abstract
:1. Introduction
2. Materials and Methods
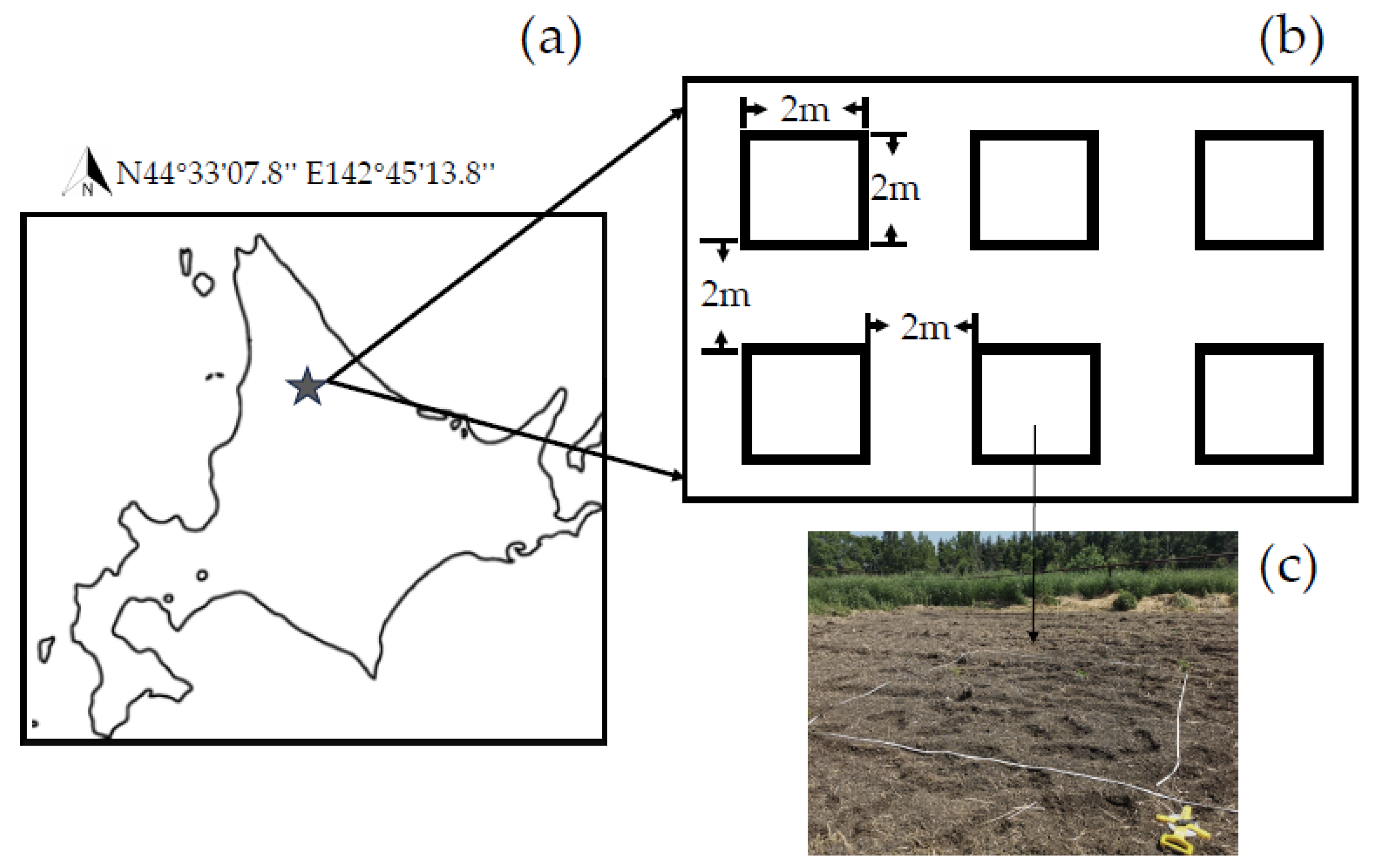
2.1. Species Selection and Experimental Design
2.2. Root Traits
2.3. Root Tensile Strength
2.4. Statistical Analysis
3. Results
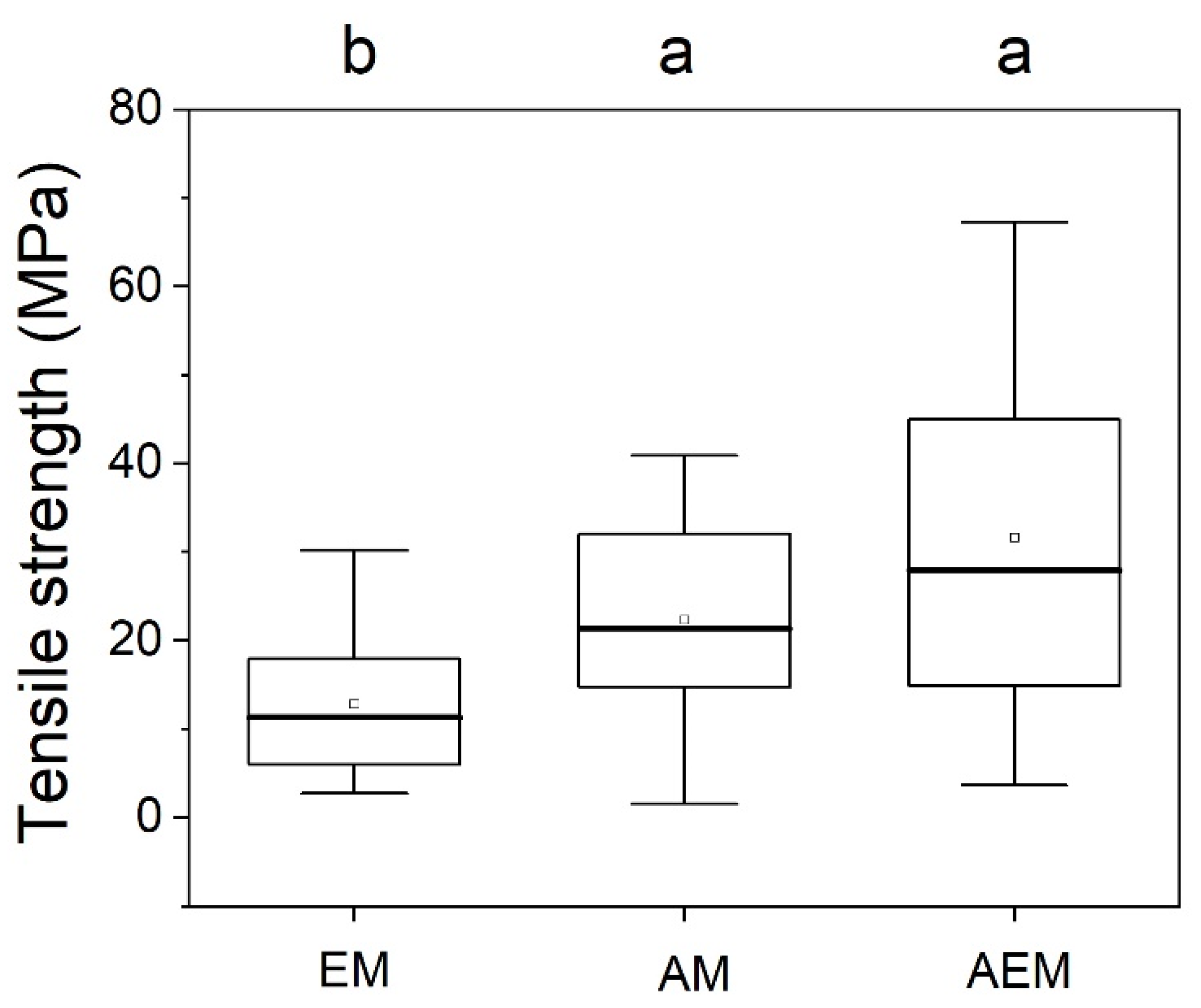
3.1. Fine Root Tr and 5 Root Traits
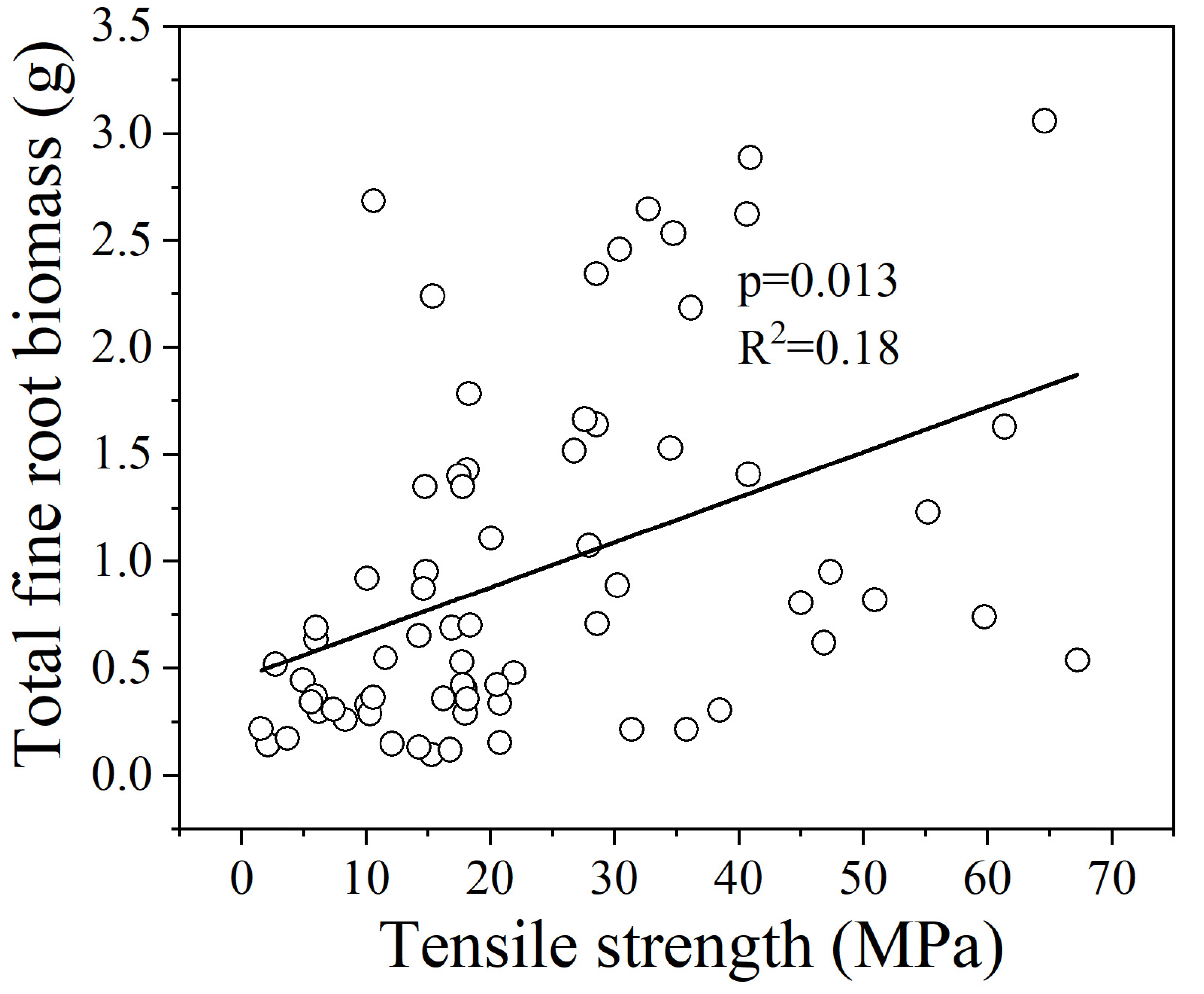
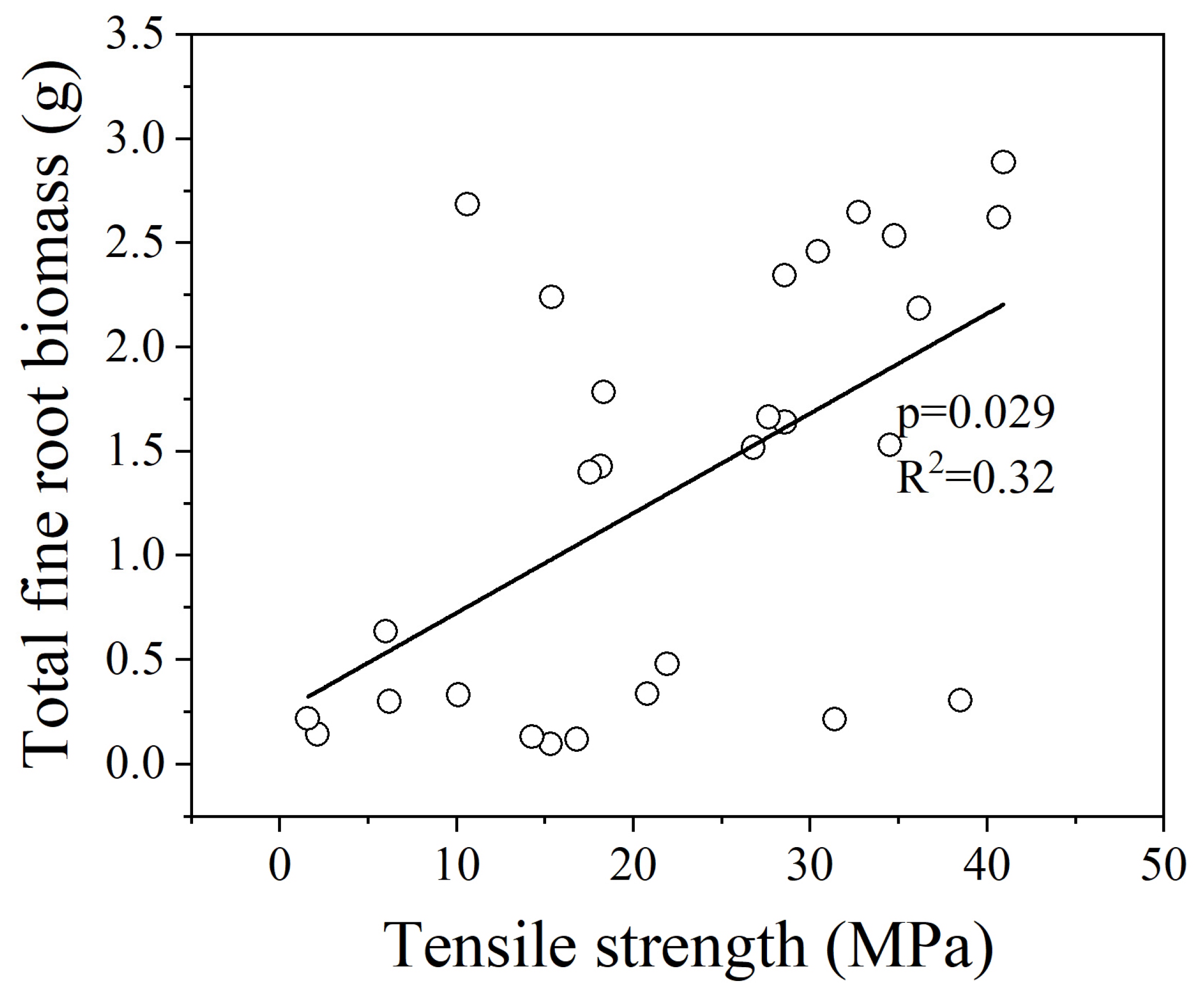
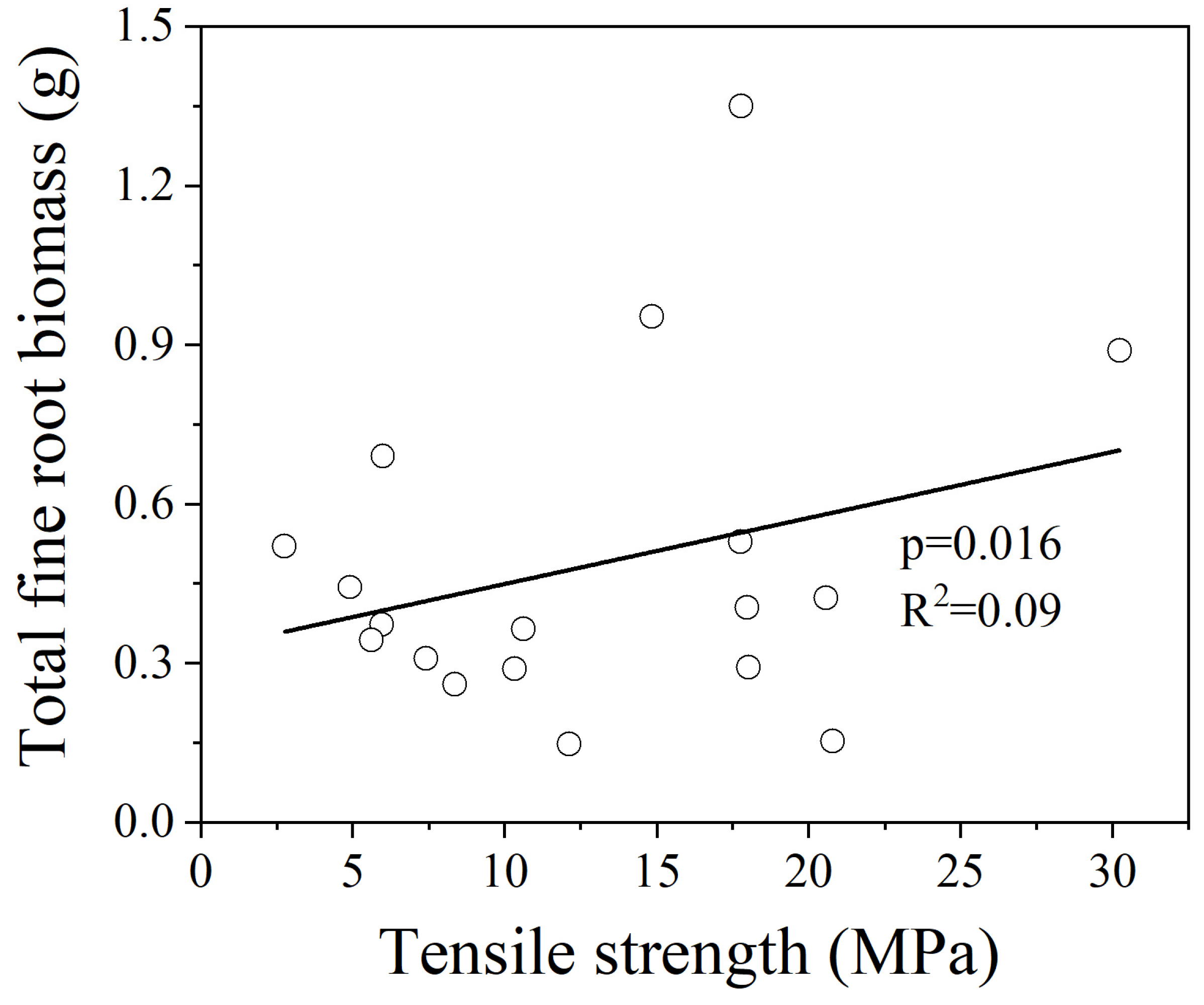
3.2. Relationships between Fine Root Tr and 5 Root Traits
4. Discussion
4.1. Determinants among Mycorrhizal Groups
4.2. Variation among Mycorrhizal Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stokes, A.; Atger, C.; Bengough, A.G.; Fourcaud, T.; Sidle, R.C. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil 2009, 324, 1–30. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, H.; He, B.H.; Yan, H.; Liu, X.H.; Qiang, J.J. Root tensile strength of terrace hedgerow plants in the karst trough valleys of SW China: Relation with root morphology and fiber content. Int. Soil. Water Consev. 2022, 10, 677–686. [Google Scholar] [CrossRef]
- Abdi, E.; Majnounian, B.; Rahimi, H.; Zobeiri, M. Distribution and tensile strength of Hornbeam (Carpinus betulus) roots growing on slopes of Caspian Forests, Iran. J. For. Res. 2009, 20, 105–110. [Google Scholar] [CrossRef]
- De Baets, S.; Poesen, J.; Reubens, B.; Wemans, K.; De Baerdemaeker, J.; Muys, B. Root tensile strength and root distribution of typical Mediterranean plant species and their contribution to soil shear strength. Plant Soil 2008, 305, 207–226. [Google Scholar] [CrossRef]
- Genet, M.; Stokes, A.; Salin, F.; Mickovski, S.B.; Fourcaud, T.; Dumail, J.F.; Beek, R.V. The influence of cellulose content on tensile strength in tree roots. Plant Soil 2005, 278, 1–9. [Google Scholar] [CrossRef]
- Ghestem, M.; Cao, K.F.; Ma, W.Z.; Rowe, N.; Leclerc, R.; Gadenne, C.; Stokes, A. A framework for identifying plant species to be used as ‘ecological engineers’ for fixing soil on unstable slopes. PLoS ONE 2014, 9, e95876. [Google Scholar] [CrossRef]
- Mahannopkul, K.; Jotisankasa, A. Influence of root suction on tensile strength of Chrysopogon zizanioides roots and its implication on bioslope stabilization. J. Mt. Sci. 2019, 16, 275–284. [Google Scholar] [CrossRef]
- Giadrossich, F.; Schwarz, M.; Cohen, D.; Cislaghi, A.; Vergani, C.; Hubble, T.; Phillips, C.; Stokes, A. Methods to measure the mechanical behaviour of tree roots: A review. Ecol. Eng. 2017, 109, 256–271. [Google Scholar] [CrossRef]
- Bischetti, G.B.; Chiaradia, E.A.; Simonato, T.; Speziali, B.; Vitali, B.; Vullo, P.; Zocco, A. Root strength and root area of forest species in Lombardy (Northern Italy). Plant Soil 2005, 278, 11–22. [Google Scholar] [CrossRef]
- Zhang, C.B.; Chen, L.H.; Jiang, J. Why fine tree roots are stronger than thicker roots: The role of cellulose and lignin in relation to slope stability. Geomorphology 2014, 206, 196–202. [Google Scholar] [CrossRef]
- Burylo, M.; Rey, F.; Mathys, N.; Dutoit, T. Plant root traits affecting the resistance of soils to concentrated flow erosion. Earth Surf. Process Landf. 2012, 37, 1463–1470. [Google Scholar] [CrossRef]
- Lee, J.T.; Chu, M.Y.; Lin, Y.S.; Kung, K.N.; Lin, W.C.; Lee, M.J. Root Traits and Biomechanical Properties of Three Tropical Pioneer Tree Species for Forest Restoration in Landslide Areas. Forests 2020, 11, 179. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.B.; Chen, L.H.; Jiang, J.; Zhou, S. Effects of gauge length and strain rate on the tensile strength of tree roots. Trees 2012, 26, 1577–1584. [Google Scholar] [CrossRef]
- Mao, Z.; Wang, Y.; McCormack, M.L.; Rowe, N.; Deng, X.B.; Yang, X.D.; Xia, S.E.; Nespoulous, J.; Sidle, R.C.; Guo, D.L.; et al. Mechanical traits of fine roots as a function of topology and anatomy. Ann. Bot. 2018, 122, 1103–1116. [Google Scholar] [CrossRef] [Green Version]
- Pregitzer, K.S. Fine roots of trees-a new perspective. New Phytol. 2002, 154, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Q.; Liu, Z.K.; Chen, H.; Tang, M. Symbiosis of arbuscular mycorrhizal fungi and Robinia pseudoacacia L. improves root tensile strength and soil aggregate stability. PLoS ONE 2016, 11, e0153378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubisch, P.; Hertel, D.; Leuschner, C. Do ectomycorrhizal and arbuscular mycorrhizal temperate tree species systematically differ in root order-related fine root morphology and biomass? Front. Plant Sci. 2015, 6, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.L.; Koide, R.T.; Adams, T.S.; De Forest, J.L.; Cheng, L.; Eissenstat, D.M. Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees. Proc. Natl. Acad. Sci. USA 2016, 113, 8741–8746. [Google Scholar] [CrossRef]
- Ma, Z.Q.; Guo, D.L.; Xu, X.L.; Bardgett, R.D.; Eissenstat, D.M.; McCormack, M.L.; Hedin, L.O. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94–97. [Google Scholar] [CrossRef] [Green Version]
- Karst, J.; Franklin, J.; Simeon, A.; Light, A.; Bennett, J.A.; Erbilgin, N. Assessing the dual-mycorrhizal status of a widespread tree species as a model for studies on stand biogeochemistry. Mycorrhiza 2021, 31, 313–324. [Google Scholar] [CrossRef]
- Cheng, L.; Chen, W.L.; Adams, T.S.; Wei, X.; Li, L.; McCormack, M.L.; DeForest, J.L.; Koide, R.T.; Eissenstat, D.M. Mycorrhizal fungi and roots are complementary in foraging within nutrient patches. Ecology 2016, 97, 2815–2823. [Google Scholar] [CrossRef] [PubMed]
- Püschel, D.; Bitterlich, M.; Rydlová, J.; Jansa, J. Facilitation of plant water uptake by an arbuscular mycorrhizal fungus: A Gordian knot of roots and hyphae. Mycorrhiza 2020, 30, 299–313. [Google Scholar] [CrossRef]
- Chen, X.W.; Kang, Y.; So, P.S.; Ng, C.W.W.; Wong, M.H. Arbuscular mycorrhizal fungi increase the proportion of cellulose and hemicellulose in the root stele of vetiver grass. Plant Soil 2018, 425, 309–319. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; ISBN 978-0-12-370526-6. [Google Scholar]
- Makoto, K.; Wilson, S.D.; Sato, T.; Blume-Werry, G.; Cornelissen, J.H.C. Synchronous and asynchronous root and shoot phenology in temperate woody seedlings. Oikos 2020, 129, 643–650. [Google Scholar] [CrossRef]
- Nakamura, Y.; Fujikawa, T.; Fujita, M. Long-term changes in the soil properties and the soil macrofauna and mesofauna of an agricultural field in northern Japan during transition from chemical-intensive farming to nature farming. J. Crop Prod. 2001, 3, 63–75. [Google Scholar] [CrossRef]
- Freschet, G.T.; Pagès, L.; Iversen, C.M.; Comas, L.H.; Rewald, B.; Roumet, C.; Klimešová, J.; Zadworny, M.; Poorter, H.; Postma, J.A.; et al. A starting guide to root ecology: Strengthening ecological concepts and standardising root classification, sampling, processing and trait measurements. New Phytol. 2021, 232, 973–1122. [Google Scholar] [CrossRef] [PubMed]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.L.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Hales, T.C.; Cole-Hawthorne, C.; Lovell, L.; Evans, S. Assessing the accuracy of simple field based root strength measurements. Plant Soil 2013, 372, 553–565. [Google Scholar] [CrossRef]
- Böhm, W. Methods of Studying Root Systems; Durham, V.D.B., Athens, F.G., Wtirzburg, O.L.L., Oak Ridge, J.S.O., Eds.; Springer: Berlin/Heidelberg, Germany, 1979; ISBN 978-3-642-67284-2. [Google Scholar]
- Mattia, C.; Bischetti, G.B.; Gentile, F. Biotechnical characteristics of root systems of typical Mediterranean species. Plant Soil 2005, 278, 23–32. [Google Scholar] [CrossRef]
- Docker, B.B.; Hubble, T.C.T. Quantifying root-reinforcement of river bank soils by four Australian tree species. Geomorphology 2008, 100, 401–418. [Google Scholar] [CrossRef]
- Ji, J.N.; Kokutse, N.; Genet, M.; Fourcaud, T.; Zhang, Z.Q. Effect of spatial variation of tree root characteristics on slope stability. A case study on Black Locust (Robinia pseudoacacia) and Arborvitae (Platycladus orientalis) stands on the Loess Plateau, China. Catena 2012, 92, 139–154. [Google Scholar] [CrossRef]
- Gao, J.; Zhou, M.Y.; Shao, J.J.; Zhou, G.Y.; Liu, R.Q.; Zhou, L.G.; Liu, H.T.; He, Y.H.; Chen, Y.; Zhou, X.H.; et al. Fine root trait-function relationships affected by mycorrhizal type and climate. Geoderma 2021, 394, 115011. [Google Scholar] [CrossRef]
- Ma, X.M.; Zhu, B.; Nie, Y.X.; Liu, Y.; Kuzyakov, Y. Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen deposition: A meta-analysis. Soil. Biol. Biochem. 2021, 163, 108418. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Z.C.; Ma, X.H.; Jin, G.Q. Foraging ability and growth performance of four subtropical tree species in response to heterogeneous nutrient environments. J. For. Res. 2010, 15, 91–98. [Google Scholar] [CrossRef]
- Zhao, J.J.; Gong, L. Response of fine root carbohydrate content to soil nitrogen addition and its relationship with soil factors in a Schrenk (Picea schrenkiana) forest. J. Plant Growth Regul. 2021, 40, 1210–1221. [Google Scholar] [CrossRef]
- Teste, F.P.; Jones, M.D.; Dickie, I.A. Dual-mycorrhizal plants: Their ecology and relevance. New Phytol. 2020, 225, 1835–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, M.E.; Turner, B.L.; Liang, C.; Clay, K.; Johnson, D.J.; Phillips, R.P. Tree mycorrhizal type predicts within-site variability in the storage and distribution of soil organic matter. Glob. Chang. Biol. 2018, 24, 3317–3330. [Google Scholar] [CrossRef]
- Graf, F.; Frei, M. Soil aggregate stability related to soil density, root length, and mycorrhiza using site-specific Alnus incana and Melanogaster variegatus s.l. Ecol. Eng. 2013, 57, 314–323. [Google Scholar] [CrossRef]
- Bast, A.; Wilcke, W.; Graf, F.; Lüscher, P.; Gärtner, H. Does mycorrhizal inoculation improve plant survival, aggregate stability, and fine root development on a coarse-grained soil in an alpine eco-engineering field experiment? J. Geophys. Res.-Biogeo. 2016, 121, 2158–2171. [Google Scholar] [CrossRef] [Green Version]
- Berta, G.; Trotta, A.; Fusconi, A.; Hooker, J.E.; Munro, M.; Atkinson, D.; Giovannetti, M.; Morini, S.; Fortuna, P.; Tisserant, B.; et al. Arbuscular mycorrhizal induced changes to plant growth and root system morphology in Prunus cerasifera. Tree Phys. 1995, 15, 281–293. [Google Scholar] [CrossRef]
- Caesar-Ton That, T.C.; Shelver, W.L.; Thorn, R.G.; Cochran, V.L. Generation of antibodies for soil aggregating basidiomycete detection as an early indicator of trends in soil quality. Appl. Soil. Ecol. 2001, 18, 99–116. [Google Scholar] [CrossRef]
- Mankel, A.; Krause, K.; Kothe, E. Identification of a hydrophobin gene that is developmentally regulated in the ectomycorrhizal fungus Tricholoma terreum. Appl. Environ. Microbiol. 2002, 68, 1408–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagu, D.; De Bellis, R.; Balestrini, R.; De Vries, O.M.H.; Piccoli, G.; Stocchi, V.; Bonfante, P.; Martin, F. Immunolocalization of hydrophobin HYDPt-1 from the ectomycorrhizal basidiomycete Pisolithus tinctorius during colonization of Eucalyptus globulus roots. New Phytol. 2008, 149, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Caravaca, F.; Barea, J.M.; Palenzuela, J.; Figueroa, D.; Alguacil, M.M.; Roldán, A. Establishment of shrub species in a degraded semiarid site after inoculation with native or allochthonous arbuscular mycorrhizal fungi. Appl. Soil. Ecol. 2003, 22, 103–111. [Google Scholar] [CrossRef]
- Chaudhary, V.B.; Bowker, M.A.; O’Dell, T.E.; Grace, J.B.; Redman, A.E.; Rillig, M.C.; Johnson, N.C. Untangling the biological contributions to soil stability in semiarid shrublands. Ecol. Appl. 2009, 19, 110–122. [Google Scholar] [CrossRef] [Green Version]
- King, E.G.; Hobbs, R.J. Identifying linkages among conceptual models of ecosystem degradation and restoration: Towards an integrative framework. Restor. Ecol. 2006, 14, 369–378. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Mycorrhizal |
|---|---|---|
| Abies sachalinensis | Pinaceae | EM |
| Picea genii | Pinaceae | EM |
| Quercus crispula | Fagaceae | EM |
| Tilia japonica | Malvaceae | EM |
| Aralia elata | Araliaceae | AM |
| Cercidiphyllum japonicum | Cercidiphyllaceae | AM |
| Fraxinus mandshurica var. Japonica | Oleaceae | AM |
| Ulmus davidiana var. japonica | Ulmaceae | AM |
| Lespedeza bicolor | Fabaceae | AM |
| Magnolia obovata | Magnoliaceae | AM |
| Alnus hirsuta | Betulaceae | AEM |
| Cerasus sargentii | Rosaceae | AEM |
| Morus australis | Moraceae | AEM |
| Salix caprea | Salicaceae | AEM |
| Sorbus commixta | Rosaceae | AEM |
| Tensile Strength (MPa) | Average Fine Root Diameter (mm) | Root Tissue Density (g/cm3) | Total Fine Root Biomass(g) | Maximum Root Depth (cm) | Specific Root Length (cm/g) | |
|---|---|---|---|---|---|---|
| ANOVA between species | F = 11.779 *** | F = 14.31 *** | F = 3.66 *** | F = 11.21 *** | F = 3.58 *** | F = 2.27 * |
| ANOVA between groups | F = 7.088 ** | F = 6.97 ** | F = 6.32 ** | F = 4.94 ** | F = 0.095 ns | F = 0.067 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, R.; Makoto, K. Is the Fine Root Tensile Strength Predictable from Structural and Morphological Traits across Mycorrhizal Types in Cool-Temperate Woody Species? Forests 2023, 14, 1542. https://doi.org/10.3390/f14081542
Zeng R, Makoto K. Is the Fine Root Tensile Strength Predictable from Structural and Morphological Traits across Mycorrhizal Types in Cool-Temperate Woody Species? Forests. 2023; 14(8):1542. https://doi.org/10.3390/f14081542
Chicago/Turabian StyleZeng, Ruiqi, and Kobayashi Makoto. 2023. "Is the Fine Root Tensile Strength Predictable from Structural and Morphological Traits across Mycorrhizal Types in Cool-Temperate Woody Species?" Forests 14, no. 8: 1542. https://doi.org/10.3390/f14081542