
Induced Drought Stress Response of European Beech Seedlings Treated with Hydrogel and Ectomycorrhizal Inoculum


Abstract
:1. Introduction
2. Materials and Methods
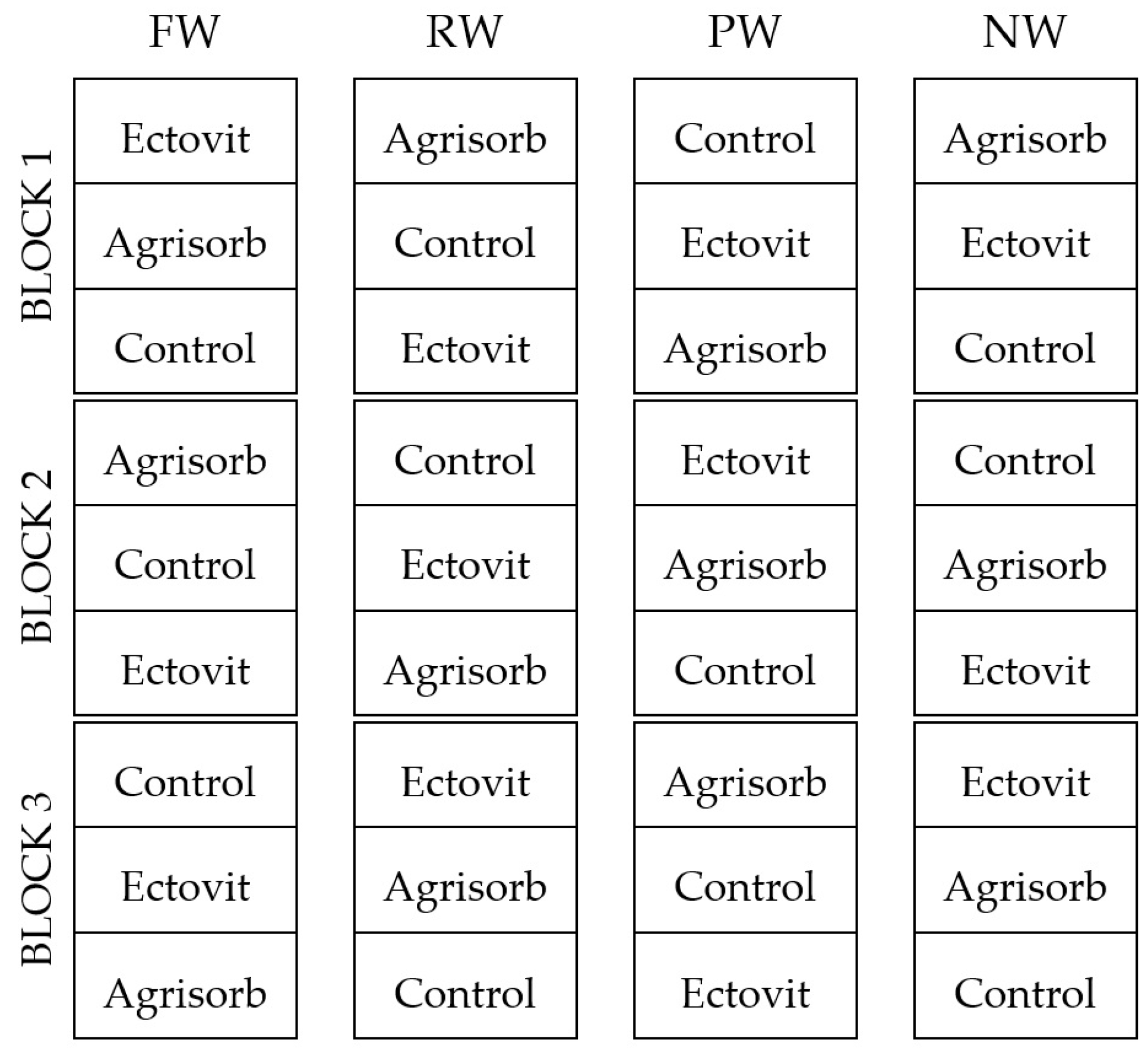
2.1. Experimental Design
- (i).
- Full watering (FW) (volumetric water content of the substrate 70%, easily available water).
- (ii).
- Reduced watering (RW) (volumetric water content 40%, poorly available water).
- (iii).
- Periodic watering (PW) (drought periods; substrate rewetted to 70% when volumetric water content decreased to wilting point (21%)).
- (iv).
- No watering (NW).
2.2. Seeds, Seedlings, and Cultural Practices
2.3. Pots, Growth Substrate, and Additives
2.4. Setting and Maintenance of Watering Regimes
2.5. Sampling and Measurements
2.5.1. Survival and Growth
2.5.2. Frequency of Short Roots and Ectomycorrhizas
2.5.3. Foliar Nutrient Concentration
2.5.4. Chlorophyll a Fluorescence
2.6. Data Analysis
3. Results
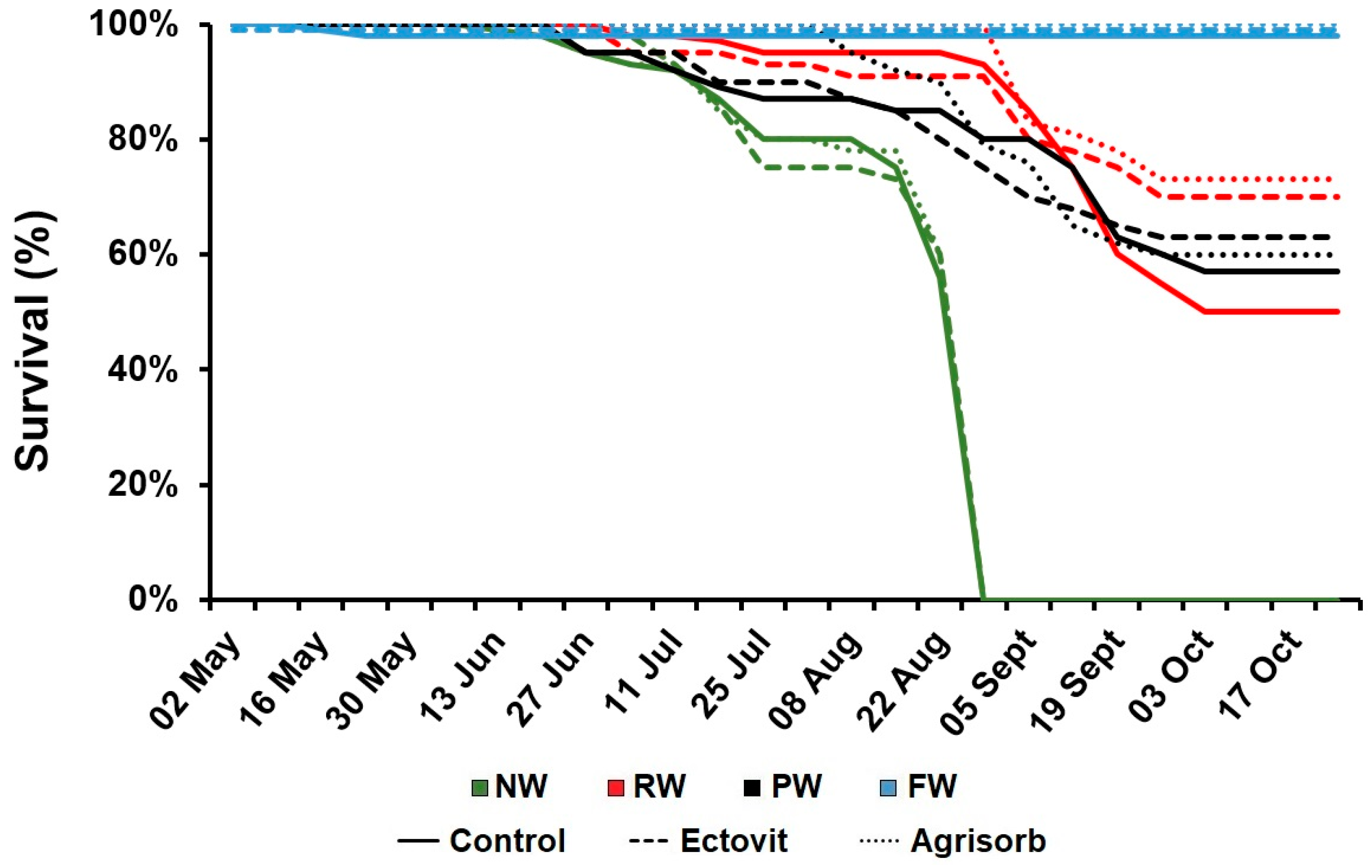
3.1. Survival
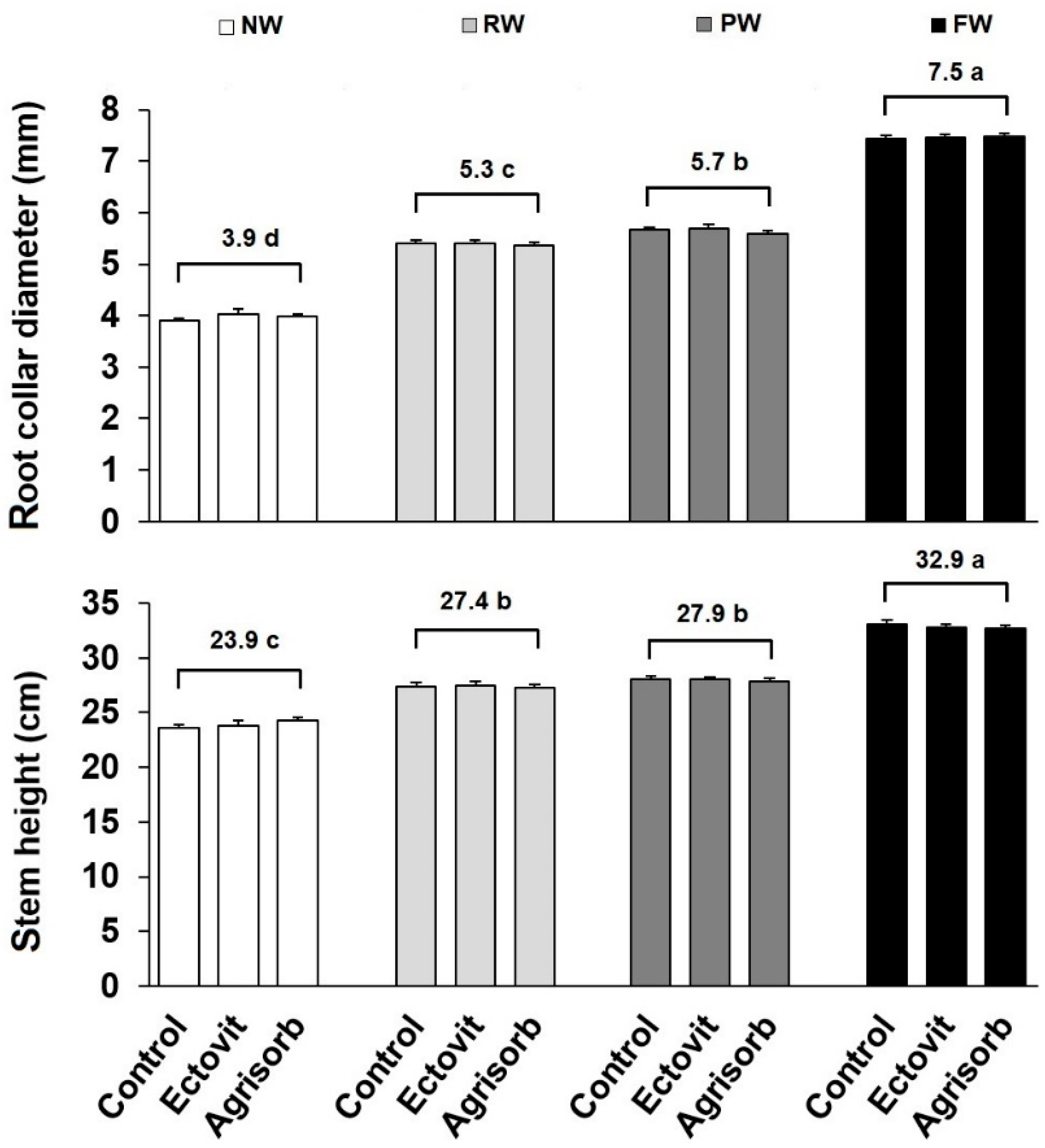
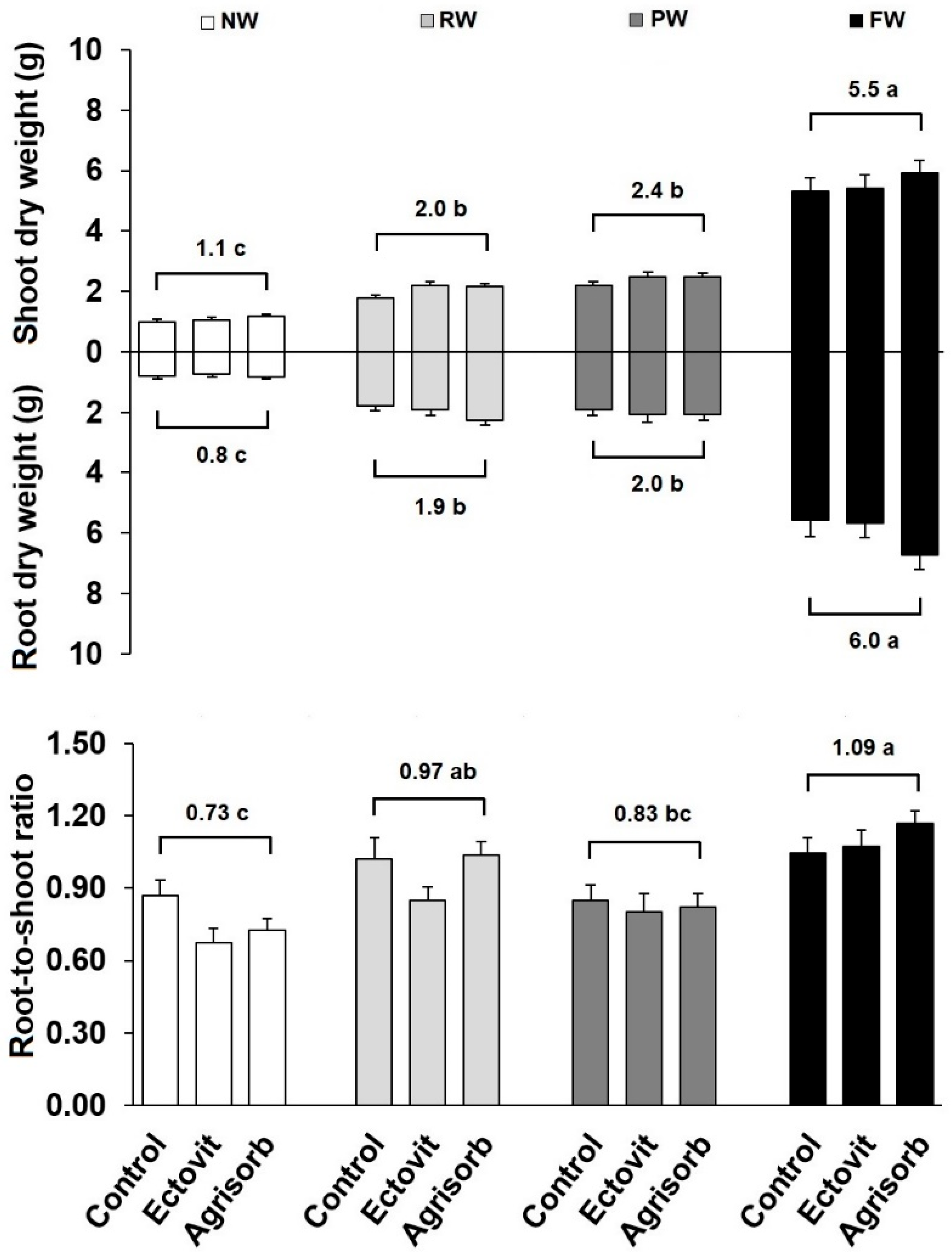
3.2. Growth
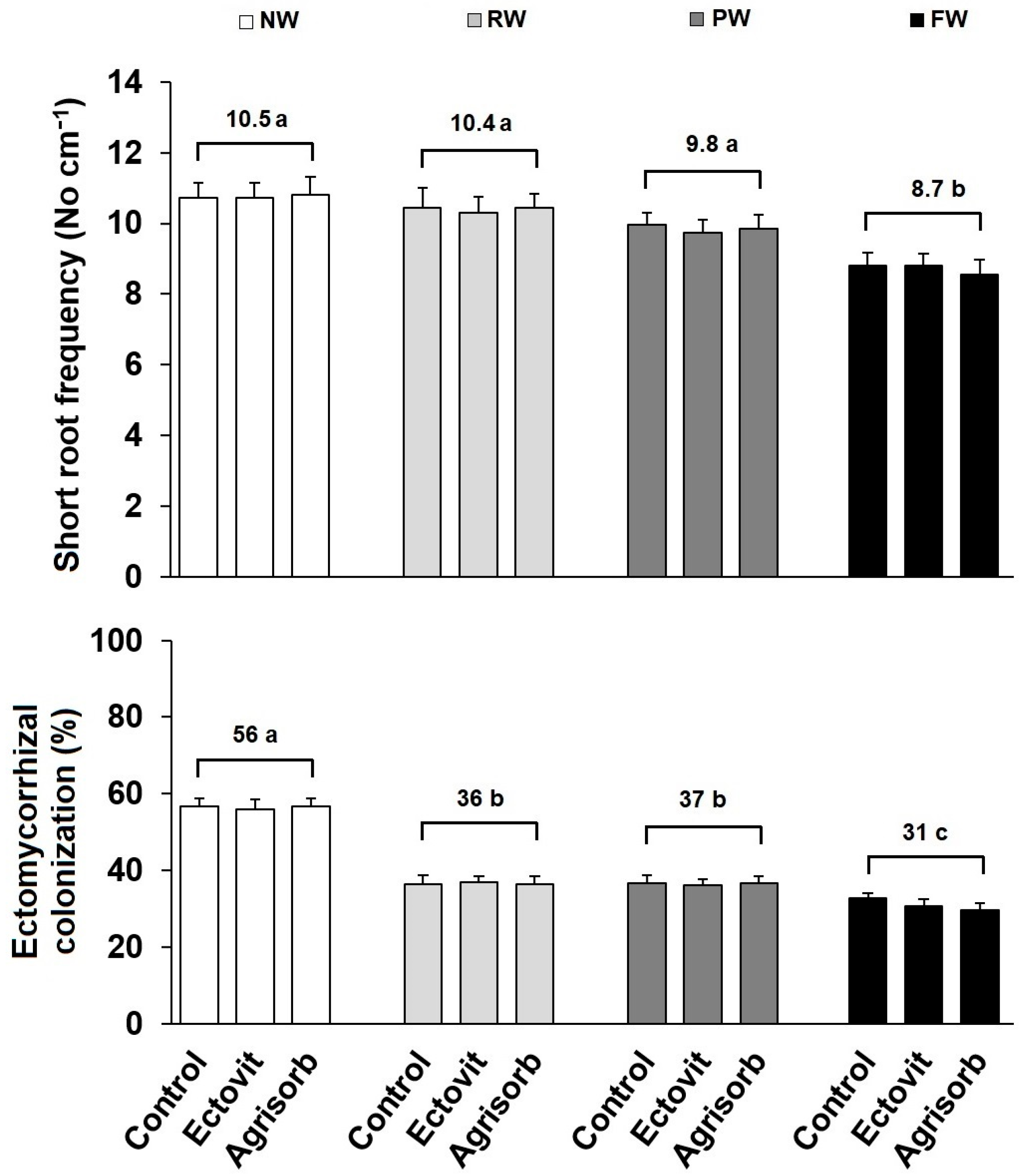
3.3. Short Roots and Ectomycorrhizas
3.4. Foliar Nutrient Concentration
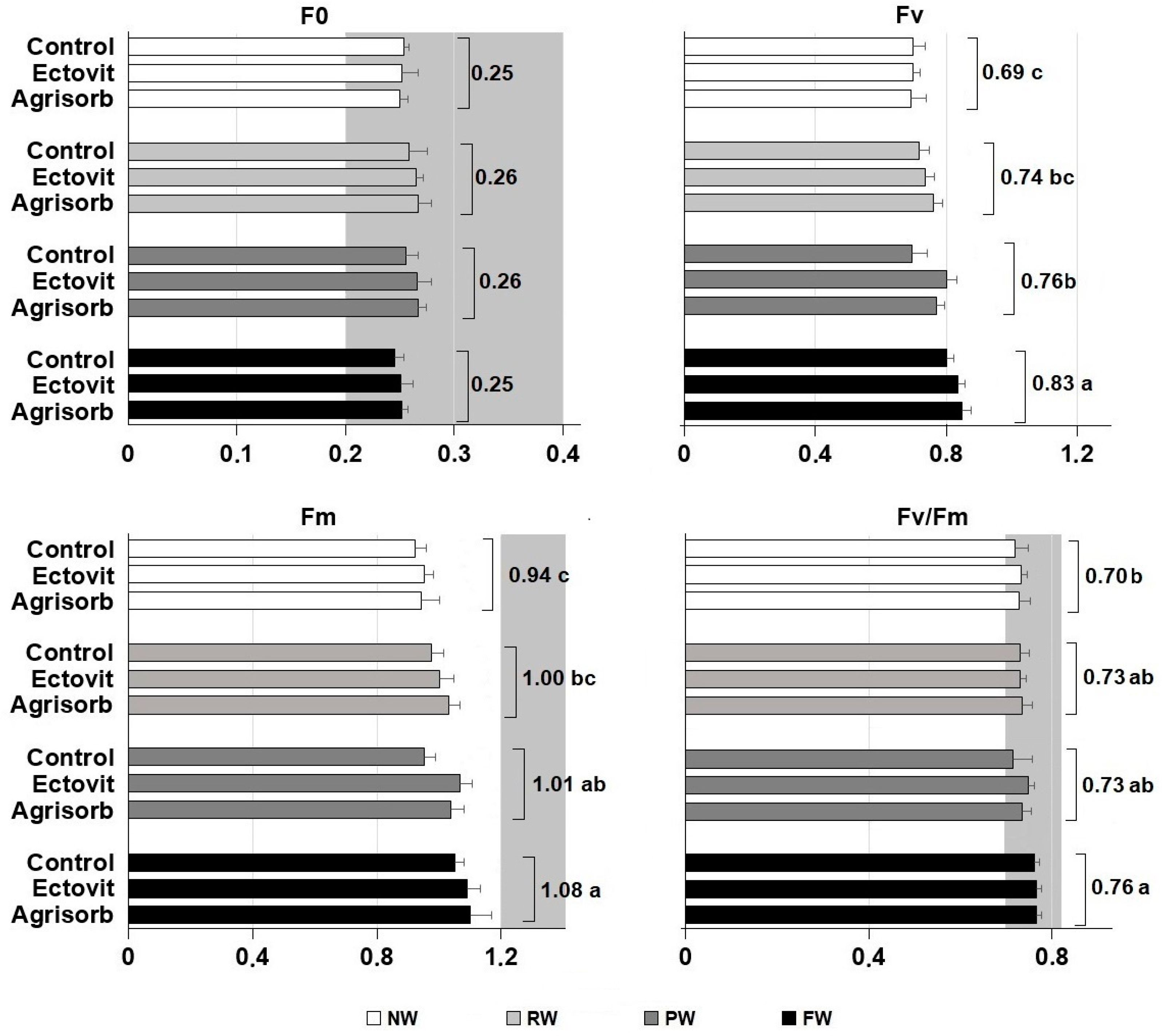
3.5. Chlorophyll a Fluorescence
4. Discussion
4.1. Effect of Water Availability on Seedling Survival and Growth
4.2. Effect of Hydrogel on Seedling Water Supply
4.3. Effect of EMF Inoculation on Seedling Development
4.4. Effect of Water Availability and Additives on Foliar Nutrient Concentration
4.5. Effect of Water Availability and Additives on Chlorophyll a Fluorescence
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ammer, C.; Kölling, C. Converting Norway spruce stands with beech—A review of arguments and techniques. Austrian J. For. Sci. 2008, 1, 3–26. [Google Scholar]
- Janda, P.; Trotsiuk, V.; Mikoláš, M.; Bače, R.; Nagel, T.A.; Seidl, R.; Seedre, M.; Morrissey, R.C.; Kucbel, S.; Jaloviar, P.; et al. The historical disturbance regime of mountain Norway spruce forests in the Western Carpathians and its influence on current forest structure and composition. For. Ecol. Manag. 2017, 388, 67–78. [Google Scholar] [CrossRef]
- Aussenac, G. Interactions between forest stands and microclimate: Ecophysiological aspects and consequences for silviculture. Ann. For. Sci. 2000, 57, 287–301. [Google Scholar] [CrossRef]
- Repáč, I.; Belko, M.; Krajmerová, D.; Paule, L. Planting time, stocktype and additive effects on the development of spruce and pine plantations in Western Carpathian Mts. New For. 2021, 52, 449–472. [Google Scholar] [CrossRef]
- Beniwal, R.S.; Hooda, S.M.; Polle, A. Amelioration of planting stress by soil amendment with a hydrogel–mycorrhiza mixture for early establishment of beech (Fagus sylvatica L.) seedlings. Ann. For. Sci. 2011, 68, 803–810. [Google Scholar] [CrossRef]
- Repáč, I.; Kmeť, J.; Vencurik, J.; Balanda, M. Effects of commercial products application on survival, growth and physiological parameters of Norway spruce and European beech plantations. Rep. For. Res.-Zpravy Lesn. Vyzk. 2013, 58, 167–175. [Google Scholar]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests, 6th ed.; Springer Nature: Cham, Switzerland, 2017; pp. 31–116. [Google Scholar]
- Vacek, Z.; Vacek, S.; Slanař, J.; Bílek, L.; Bulušek, D.; Štefančík, I.; Králíček, I.; Vančura, K. Adaption of Norway spruce and European beech forests under climate change: From resistance to close-to-nature silviculture. Cent. Eur. For. J. 2019, 65, 129–144. [Google Scholar] [CrossRef]
- Anonymous. Report on Forestry in the Slovak Republic 2022—Green Report; Ministry of Agriculture and Rural Development of the Slovak Republic: Bratislava, Slovakia, 2022; pp. 7–21.
- Grossnickle, S.C.; El-Kassaby, A.Y. Bareroot versus container stocktypes: A performance comparison. New For. 2016, 47, 1–51. [Google Scholar] [CrossRef]
- Wagner, S.; Collet, C.; Madsen, P.; Nakashizuka, T.; Nyland, R.D.; Sagheb-Talebi, K. Beech regeneration research: From ecological to silvicultural aspects. For. Ecol. Manag. 2010, 259, 2172–2182. [Google Scholar] [CrossRef]
- Bolte, A.; Czajkowski, T.; Cocozza, C.; Tognetti, R.; Miguel, M.; Pšidová, E.; Ditmarová, Ľ.; Dinca, L.; Delzon, S.; Cochard, H.; et al. Desiccation and mortality dynamics in seedlings of different European beech (Fagus sylvatica L.) populations under extreme drought conditions. Front. Plant. Sci. 2016, 14, 751. [Google Scholar] [CrossRef]
- Manzanedo, R.D.; Schanz, F.R.; Fischer, M.; Allan, E. Fagus sylvatica seedlings show provenance differentiation rather than adaptation to soil in a transplant experiment. BMC Ecol. 2018, 18, 42. [Google Scholar] [CrossRef]
- Robakowski, P.; Wyka, T.P.; Kowalkowski, W.; Barzdajn, W.; Pers-Kamczyc, E.; Jankowski, A.; Politycka, B. Practical implications of different phenotypic and molecular responses of evergreen conifer and broadleaf deciduous forest tree species to regulated water deficit in a container nursery. Forests 2020, 11, 1011. [Google Scholar] [CrossRef]
- Bhardwaj, A.K.; Shainberg, I.; Goldstein, D.; Warrington, D.N.; Levy, G.J. Water retention and hydraulic conductivity of cross-linked polyacrylamides in sandy soil. Soil Sci. Soc. Am. J. 2007, 71, 406–412. [Google Scholar] [CrossRef]
- Thomas, D.S. Hydrogel applied to the root plug of subtropical eucalypt seedlings halves transplant death following planting. For. Ecol. Manag. 2008, 255, 1305–1314. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Vallejo, V.R. Using hydrogel and clay to improve the water status of seedlings for dryland restoration. Plant Soil 2011, 344, 99–110. [Google Scholar] [CrossRef]
- Jamnická, G.; Ditmarová, Ľ.; Kmeť, J.; Pšidová, E.; Macková, M.; Gömöry, D.; Střelcová, K. The soil hydrogel improved photosynthetic performance of beech seedlings treated under drought. Plant Soil Environ. 2013, 59, 446–451. [Google Scholar] [CrossRef]
- Fonteno, W.C.; Bilderback, T.E. Impact of hydrogel on physical properties of coarse-structured horticultural substrates. J. Am. Soc. Hortic. Sci. 1993, 118, 217–222. [Google Scholar] [CrossRef]
- Rowe, E.C.; Williamson, J.C.; Jones, D.L.; Holliman, P.; Healey, J.R. Initial tree establishment on blocky quarry waste ameliorated with hydrogel or slate processing fines. J. Environ. Qual. 2005, 34, 994–1003. [Google Scholar] [CrossRef]
- Kropp, B.R.; Langlois, C.G. Ectomycorrhizae in reforestation. Can. J. For. Res. 1990, 20, 438–451. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; pp. 191–385. [Google Scholar]
- Marjanović, Ž.; Nehls, U. Ectomycorrhiza and water transport. In Mycorrhiza, State of the Art, Genetics and Molecular Biology, Eco-Function, Biotechnology, Eco-Physiology, Structure and Systematics; Varma, A., Ed.; Springer: Berlin, Germany, 2008; pp. 149–161. [Google Scholar]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenlid, J.; Finlay, R. Afforestation of abandoned farmland with conifer seedlings inoculated with three ectomyorrhizal fungi—Impact on plant performance and ectomycorrhizal community. Mycorrhiza 2007, 17, 337–348. [Google Scholar] [CrossRef]
- Quoreshi, A.M.; Piché, Y.; Khasa, P.D. Field performance of conifer and hardwood species 5 years after nursery inoculation in the Canadian Prairie Provinces. New For. 2008, 35, 235–253. [Google Scholar] [CrossRef]
- Holuša, J.; Pešková, V.; Lorenc, F. The impact of artificial inoculation on the growth of common oak seedlings and development of mycorrhiza: Inoculation may not positively affect growth of seedlings. Period. Biol. 2016, 117, 519–526. [Google Scholar] [CrossRef]
- Ortega, U.; Duñabeitia, M.; Menendez, S.; Gonzalez-Murua, C.; Majada, J. Effectiveness of mycorrhizal inoculation in the nursery on growth and water relation of Pinus radiata in different water regimes. Tree Physiol. 2004, 24, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Parladé, J.; Luque, J.; Pera, J.; Rincón, A.M. Field performance of Pinus pinea and P. halepensis seedlings inoculated with Rhizopogon spp. and outplanted in formerly arable land. Ann. For. Sci. 2004, 61, 507–514. [Google Scholar] [CrossRef]
- Pešková, V.; Tuma, M. Infuence of artifcial mycorrhization on development of spruce seedlings at Jablunkov (forest district). Rep. For. Res.-Zpravy Lesn. Vyzk. 2010, 55, 211–220. [Google Scholar]
- Orikiriza, L.J.B.; Agaba, H.; Eilu, G.; Kabasa, D.J.; Worbes, M.; Hüttermann, A. Effects of hydrogels on tree seedling performance in temperate soils before and after water stress. J. Environ. Protect. 2013, 4, 713–721. [Google Scholar] [CrossRef]
- Pena, R.; Simon, J.; Rennenberg, H.; Polle, A. Ectomycorrhiza affect architecture and nitrogen partitioning of beech (Fagus sylvatica L.) under shade and drought. Environ. Exp. Bot. 2013, 87, 207–217. [Google Scholar] [CrossRef]
- Leberecht, M.; Tu, J.; Polle, A. Acid and calcareous soils affect nitrogen nutrition and organic nitrogen uptake by beech seedlings (Fagus sylvatica L.) under drought, and their ectomycorrhizal community structure. Plant Soil 2016, 409, 143–157. [Google Scholar] [CrossRef]
- Moser, B.; Kipfer, T.; Richter, S.; Egli, S.; Wohlgemuth, T. Drought resistance of Pinus sylvestris seedlings conferred by plastic root architecture rather than ectomycorrhizal colonisation. Ann. For. Sci. 2015, 72, 303–309. [Google Scholar] [CrossRef]
- Crous, J.W. Use of hydrogels in the planting of industrial wood plantations. South. For. J. For. Sci. 2016, 79, 197–213. [Google Scholar] [CrossRef]
- Slovak Hydrometeorological Institute. Available online: http://klimat.shmu.sk/kas (accessed on 2 March 2023).
- DeBoodt, M.; Verdonck, O.; Cappaert, I. Method for measuring water release curve of organic substrates. Acta Hortic. 1974, 37, 2054–2062. [Google Scholar] [CrossRef]
- Prasad, M.; O’Shea, J. Relative breakdown of peat and non-peat growing media. Acta Hortic. 1999, 481, 121–128. [Google Scholar] [CrossRef]
- Cocozza, C.; Miguel, M.; Pšidová, E.; Ditmarová, Ľ.; Marino, S.; Maiuro, L.; Alvino, A.; Czajkowski, T.; Bolte, A.; Tognetti, R. Variation in ecophysiological traits and drought tolerance of beech (Fagus sylvatica L.) seedlings from different populations. Front. Plant Sci. 2016, 7, 886. [Google Scholar] [CrossRef] [PubMed]
- Vaario, L.M.; Tervonen, A.; Haukioja, K.; Haukioja, M.; Pennanen, T.; Timonen, S. The effect of nursery substrate and fertilization on the growth and ectomycorrhizal status of containerized and outplanted seedlings of Picea abies. Can. J. For. Res. 2009, 39, 64–75. [Google Scholar] [CrossRef]
- DEEMY—An Information System for Characterization and Determination of Ectomycorrhizae. Available online: http://www.deemy.de (accessed on 30 March 2023).
- Ritchie, G.A. Chlorophyll fluorescence: What is it and what do the numbers mean? In National Proceedings: Forest and Conservation Nursery Associations. Proc. RMRS-P-43; Riley, E.L., Dumroese, R.K., Landis, T.D., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006; pp. 34–42. [Google Scholar]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef]
- Pflug, E.E.; Buchmann, N.; Siegwolf, R.T.W.; Schaub, M.; Rigling, A.; Arend, M. Resilient leaf physiological response of European beech (Fagus sylvatica L.) to summer drought and drought release. Front. Plant Sci. 2018, 9, 187. [Google Scholar] [CrossRef]
- Leuschner, C. Drought response of European beech (Fagus sylvatica L.)—A review. Perspect. Plant Ecol. Evol. Syst. 2020, 47, 125576. [Google Scholar] [CrossRef]
- Zang, U.; Goisser, M.; Häberle, K.H.; Matyssek, R.; Matzner, E.; Borken, W. Effects of drought stress on photosynthesis, rhizosphere respiration, and fine-root characteristics of beech saplings: A rhizotron field study. J. Plant Nutr. Soil Sci. 2014, 177, 168–177. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analysis of interspecific variation and environmental control. New Phytol. 2011, 193, 30–50. [Google Scholar] [CrossRef]
- Edwards, C.E.; Ewers, B.E.; Weinig, C. Genotypic variation in biomass allocation in response to field drought has a greater effect on yield than gas exchange or phenology. BMC Plant Biol. 2016, 16, 185. [Google Scholar] [CrossRef]
- Bengough, A.G.; Bransby, M.F.; Hans, J.; McKenna, S.J.; Roberts, T.J.; Valentine, T.A. Root responses to soil physical conditions; growth dynamics from field to cell. J. Exp. Bot. 2006, 57, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Sloan, J.P. The use of rootdips on North American conifer seedlings: A review of the literature. Tree Planter’s Notes 1994, 45, 26–31. [Google Scholar]
- Apostol, K.G.; Jacobs, D.F.; Dumroese, R.K. Root desiccation and drought stress responses of bareroot Quercus rubra seedlings treated with a hydrophilic polymer root dip. Plant Soil 2009, 315, 229–240. [Google Scholar] [CrossRef]
- Wang, Y.T.; Gregg, L.L. Hydrophilic polymers—Their response to soil amendments and effect on properties of a soilless potting mix. J. Am. Soc. Hortic. Sci. 1990, 115, 943–948. [Google Scholar] [CrossRef]
- Ghebru, M.G.; du Toit, E.S.; Steyn, J.M. Water and nutrient retention by AquaSoil® and Stockosorb® polymers. S. Afr. J. Plant Soil 2007, 24, 32–36. [Google Scholar] [CrossRef]
- Castellano, M.A. Outplanting performance of mycorrhizal inoculated seedlings. In Concepts in Mycorrhizal Research; Mukerji, K.G., Ed.; Kluwer Academic Publisher: Dordrecht, The Netherlands, 1996; pp. 223–301. [Google Scholar]
- Sanchez-Zabala, J.; Majada, J.; Martín-Rodrigues, N.; Gonzalez-Murua, C.; Ortega, U.; Alonso-Graña, M.; Arana, O.; Duñabeitia, K.M. Physiological aspects underlying the improved outplanting performance of Pinus pinaster Ait. seedlings associated with ectomycorrhizal inoculation. Mycorrhiza 2013, 23, 627–640. [Google Scholar] [CrossRef]
- Repáč, I.; Parobeková, Z.; Belko, M. Ectomycorrhiza-hydrogel additive enhanced growth of Norway spruce seedlings in a nutrient-poor peat substrate. J. For. Sci. 2022, 68, 170–181. [Google Scholar] [CrossRef]
- Socha, J.; Orzel, S.; Ochal, W.; Pietrzykowski, M. Effect of seedling production method on the growth of Pinus sylvestris L. on reclamation post-industrial sites in Poland. Dendrobiology 2022, 88, 124–137. [Google Scholar] [CrossRef]
- Duponnois, R.; Kisa, M.; Prin, Y.; Ducousso, M.; Plenchette, C.; Lepage, M.; Galiana, A. Soil factors influencing the growth response of Acacia holosericea A. Cunn. ex G. don to ectomycorrhizal inoculation. New For. 2008, 35, 105–117. [Google Scholar] [CrossRef]
- Marx, D.H.; Ruehle, J.L.; Kenney, D.S.; Cordell, C.E.; Riffle, J.W.; Molina, R.J.; Pawuk, W.H.; Navratil, S.; Tinus, R.W.; Goodwin, O.C. Commercial vegetative inoculum of Pisolithus tinctorius and inoculation techniques for development of ectomycorrhizae on container-grown tree seedlings. For. Sci. 1982, 28, 373–400. [Google Scholar]
- Repáč, I. Inoculation of Picea abies [L.] Karst. seedlings with vegetative inocula of ectomycorrhizal fungi Suillus bovinus (L.:Fr.) O. Kuntze and Inocybe lacera (Fr.) Kumm. New For. 1996, 12, 41–54. [Google Scholar] [CrossRef]
- Wiemken, V.; Ineichen, K.; Boller, T. Development of ectomycorrhizas in model beech-spruce ecosystems on siliceous and calcareous soil: A 4-year experiment with atmospheric CO2 enrichment and nitrogen fertilization. Plant Soil 2001, 234, 99–108. [Google Scholar] [CrossRef]
- Lonergan, E.R.; Cripps, C.L. Use of low nitrogen fertilizer as a strategy for maintaining mycorrhizal colonization on white bark pine seedlings inoculated with native fungi in the greenhouse. Native Plants J. 2013, 14, 213–224. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. Fungal reactions to plants prior to mycorrhizal formation. In Mycorrhizal Functioning: An Integrative Plant-Fungal Process, 1st ed.; Allen, M.F., Ed.; Chapman and Hall: London, UK, 1992; pp. 3–36. [Google Scholar]
- Shi, L.; Guttenberger, M.; Kottke, I.; Hampp, R. The effect of drought on mycorrhizas of beech (Fagus sylvatica L.): Changes in community structure, and the content of carbohydrates and nitrogen storage bodies of the fungi. Mycorrhiza 2002, 12, 303–311. [Google Scholar] [CrossRef]
- Nehls, U.; Göhringer, F.; Wittulsky, S.; Dietz, S. Fungal carbohydrate support in the ectomycorrhizal symbiosis: A review. Plant Biol. 2010, 12, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, M.; Churin, J.L.; Garbaye, J. Differential ability of ectomycorrhizas to survive drying. Mycorrhiza 2007, 17, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Pena, R.; Polle, A. Attributing functions to ectomycorrhizal fungal identities in assemblages for nitrogen acquisition under stress. ISME J. 2014, 8, 321–330. [Google Scholar] [CrossRef]
- Brunner, I.; Scheidegger, C. Effects of high nitrogen concentrations on ectomycorrhizal structure and growth of seedlings of Picea abies (L.) Karst. New Phytol. 1995, 129, 83–95. [Google Scholar] [CrossRef]
- Morin, C.; Samson, J.; Dessureault, M. Protection of black spruce seedlings against Cylindrocladium root rot with ectomycorrhizal fungi. Can. J. Bot. 1999, 77, 169–174. [Google Scholar] [CrossRef]
- Sebastiana, M.; Bernardes da Silva, A.; Matos, A.R.; Alcântara, A.; Silvestre, S.; Malhó, R. Ectomycorrhizal inoculation with Pisolithus tinctorius reduces stress induced by drought in cork oak. Mycorrhiza 2018, 28, 247–258. [Google Scholar] [CrossRef]
- Wallin, G.; Karlsson, P.E.; Selldén, G.; Ottosson, S.; Medin, E.L.; Pleijel, H.; Skärby, L. Impact of four years exposure to different levels of ozone, phosphorus and drought on chlorophyll, mineral nutrients, and stem volume of Norway spruce, Picea abies. Physiol. Plant. 2002, 114, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Lamhamedi, M.S.; Lambany, G.; Margolis, H.A.; Renaud, M.; Veilleux, I.; Bernier, P.Y. Growth, physiology and leachate losses in Picea glauca seedlings (1 + 0) grown in air-slit container under different irrigation regimes. Can. J. For. Res. 2001, 31, 1968–1980. [Google Scholar] [CrossRef]
- Mellert, H.K.; Göttlein, A. Comparison of new foliar nutrient thresholds derived from van den Burg’s literature compilation with established central European references. Eur. J. For. Res. 2012, 131, 1461–1472. [Google Scholar] [CrossRef]
- Peuke, A.D.; Rennenberg, H. Carbon, nitrogen, phosphorus, and sulphur concentration and partitioning in beech ecotypes (Fagus sylvatica L.): Phosphorus most affected by drought. Trees 2004, 18, 639–648. [Google Scholar] [CrossRef]
- Bowman, D.C.; Evans, R.Y.; Paul, J.L. Fertilizer salts reduce hydration of polyacrylamide gels and affect physical properties of gel-amended container media. J. Am. Soc. Hortic. Sci. 1990, 115, 382–386. [Google Scholar] [CrossRef]
- Mikkelsen, R.L. Using hydrophilic polymers to control nutrient release. Fertil. Res. 1994, 38, 53–59. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Watering | Additive | Watering × Additive | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| Survival | 311.4 | 0.001 | 6.4 | 0.056 | 4.1 | 0.017 |
| Root collar diameter | 2276.1 | 0.001 | 1.2 | 0.368 | 0.4 | 0.853 |
| Stem height | 207.4 | 0.001 | 0.1 | 0.999 | 0.5 | 0.776 |
| Shoot dry weight | 164.3 | 0.001 | 0.9 | 0.455 | 0.2 | 0.973 |
| Root dry weight | 656.1 | 0.001 | 3.4 | 0.135 | 0.7 | 0.634 |
| Root-to-shoot ratio | 14.5 | 0.004 | 1.2 | 0.386 | 1.1 | 0.428 |
| Short root frequency | 12.9 | 0.005 | 0.1 | 0.861 | 0.1 | 0.991 |
| EMF colonization | 516.0 | 0.001 | 0.3 | 0.789 | 0.2 | 0.971 |
| F0 | 2.8 | 0.128 | 0.7 | 0.561 | 0.5 | 0.799 |
| Fm | 14.8 | 0.003 | 5.3 | 0.075 | 0.7 | 0.636 |
| Fv | 17.9 | 0.002 | 5.9 | 0.063 | 0.6 | 0.748 |
| Fv/Fm | 14.5 | 0.004 | 3.3 | 0.143 | 0.4 | 0.882 |
| EMF Morphotypes (25× Magnification) | Description of EMF Morphotypes and Identification of Ectomycorrhizal Fungi via DNA Sequencing |
|---|---|
 | White color, ramification absent, short and straight shape with inflated distal end, shiny mantle, white tips, and whitish hyphae. Ectomycorrhizal fungus identified: Hebeloma sacchariolens Quél. NCBI database record, Accession No: MT929352, similarity 99%. |
 | Ochre to yellowish-brown color, monopodial pinnate ramification, straight or bent shape, loosely cottony mantle covered with soil particles, brown tips, yellowish hyphae. Ectomycorrhizal fungus identified: Hymenogaster vulgaris Tul. and C. Tul. NCBI database record, Accession No: MT929774, similarity 99%. |
 | Yellowish-brown color, ramification absent, sinuous shape, smooth densely wooly mantle, yellowish-brown tips, and whitish hyphae. Unsuccessful DNA sequencing. |
 | Yellowish-brown color, monopodial-pinnate ramification, straight or bent shape with inflated distal end, shiny loosely cottony mantle, ochre tips, no emanating hyphae. Unsuccessful DNA sequencing. |
 | Brown color, ramification absent, straight or bent shape, smooth, loosely cottony mantle, brownish tips, no emanating hyphae. Unsuccessful DNA sequencing. |
| Treatment | N (%) | P | K | Ca | Mg |
|---|---|---|---|---|---|
| (mg kg−1) | (mg kg−1) | (mg kg−1) | (mg kg−1) | ||
| No watering | |||||
| Control | 2.9 | 4650 | 10,200 | 11,500 | 2640 |
| Ectovit | 2.9 | 5630 | 10,100 | 12,100 | 2420 |
| Agrisorb | 3.3 | 5820 | 10,500 | 11,700 | 2820 |
| Reduced watering | |||||
| Control | 3.2 | 6730 | 9050 | 12,300 | 2120 |
| Ectovit | 3.2 | 6540 | 8900 | 13,300 | 2170 |
| Agrisorb | 3.2 | 7180 | 8910 | 14,800 | 2560 |
| Periodic watering | |||||
| Control | 2.9 | 5910 | 8380 | 9160 | 1990 |
| Ectovit | 3.2 | 7060 | 9510 | 12,200 | 2590 |
| Agrisorb | 3.2 | 7080 | 9460 | 10,700 | 2630 |
| Full watering | |||||
| Control | 2.9 | 6010 | 9990 | 8830 | 1910 |
| Ectovit | 2.9 | 4500 | 8380 | 8170 | 1640 |
| Agrisorb | 2.8 | 5970 | 9070 | 8740 | 1800 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Repáč, I.; Belko, M.; Krajmerová, D.; Kmet’, J.; Pavlík, M. Induced Drought Stress Response of European Beech Seedlings Treated with Hydrogel and Ectomycorrhizal Inoculum. Forests 2023, 14, 1749. https://doi.org/10.3390/f14091749
Repáč I, Belko M, Krajmerová D, Kmet’ J, Pavlík M. Induced Drought Stress Response of European Beech Seedlings Treated with Hydrogel and Ectomycorrhizal Inoculum. Forests. 2023; 14(9):1749. https://doi.org/10.3390/f14091749
Chicago/Turabian StyleRepáč, Ivan, Martin Belko, Diana Krajmerová, Jaroslav Kmet’, and Martin Pavlík. 2023. "Induced Drought Stress Response of European Beech Seedlings Treated with Hydrogel and Ectomycorrhizal Inoculum" Forests 14, no. 9: 1749. https://doi.org/10.3390/f14091749