
Physiological and Biochemical Traits of Needles Imply That Understory Light Conditions in the Growing Season May Be Favorable to Pinus koraiensis Trees


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
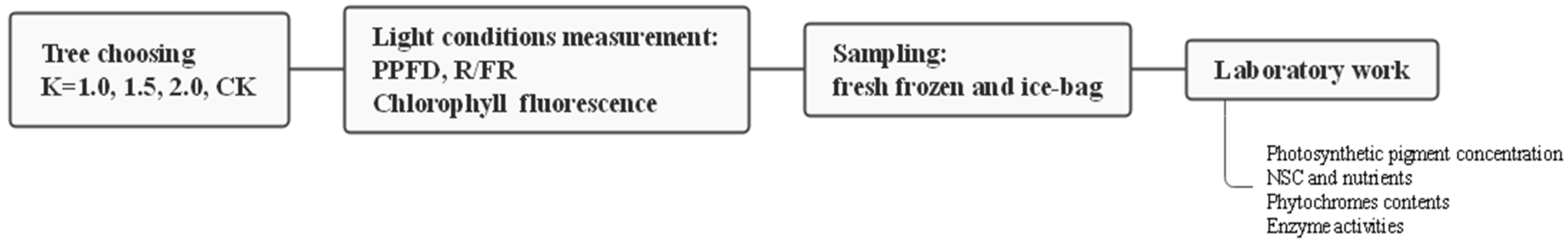
2.2. Experimental Design and Sampling
2.3. Light Conditions and Chlorophyll Fluorescence Measurement
2.4. Photosynthetic Pigment Concentration
2.5. Non-Structural Carbohydrate and Nutrient Concentrations
2.6. Enzyme Activity, DPPH Scavenging Capacity, Proline, and ABA Concentration
2.7. Data Analysis
3. Results
3.1. Dynamics of PPFD and R/FR under Different Light Conditions throughout the Research Period
3.2. Effects of Light on Photosynthetic Pigment Concentration, Chlorophyll Fluorescence, and Rubisco Activity
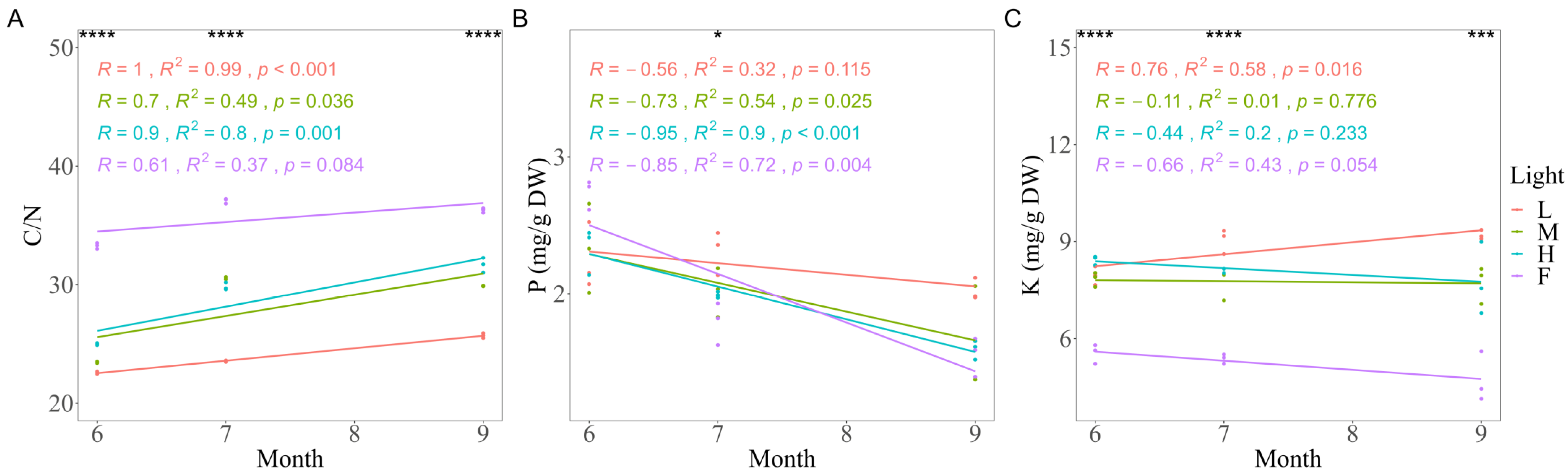
3.3. Effects of Light on NSC and Nutrient Variation
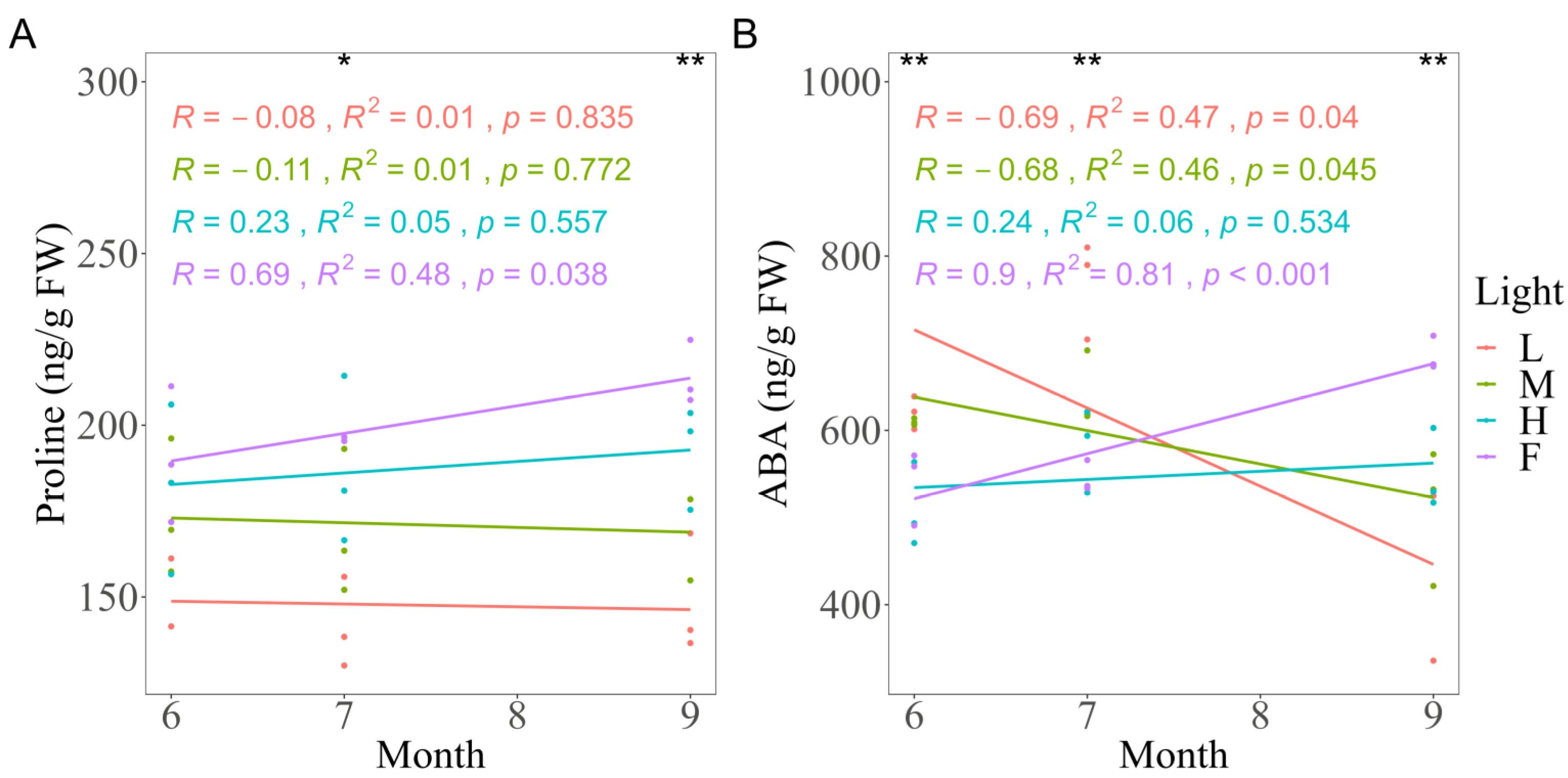
3.4. Effects of Light on Proline and ABA Concentration, Enzyme Activity, and DPPH Scavenging Capacity
4. Discussion
4.1. Dynamics of Light Affect Plant Photosynthetic Responses
4.2. NSC Concentrations in Korean Pine Needles Varied under Different Light Conditions
4.3. Anti-Stress Responses in Korean Pine Needles under Different Light Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, J.M.; Gu, L.H.; Warren, J.M.; Guha, A.; Mclennan, D.A.; Zhang, W.F.; Zhang, Y.L. The roles of photochemical and non-photochemical quenching in regulating photosynthesis depend on the phases of fluctuating light conditions. Tree Physiol. 2022, 42, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Niyogi, K.K. Manipulation of photoprotection to improve plant photosynthesis. Plant Physiol. 2011, 155, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifuentes, L.; Moreno, F. Trait coordination at leaf level explains the resistance to excess light stress in shade-tolerant tropical tree species. Tree Physiol. 2022, 42, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Chazdon, R.L. Fetcher N Photosynthetic light environments in a lowland tropical rain forest in Costa Rica. J. Ecol. 1984, 72, 553–564. [Google Scholar] [CrossRef]
- Way, D.A. Pearcy RW Sunflecks in trees and forests: From photosynthetic physiology to global change biology. Tree Physiol. 2012, 32, 1066–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K. Vegetation stress: An introduction to the stress concept in plants. J. Plant Physiol. 1996, 148, 4–14. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef] [Green Version]
- Song, X.H.; Li, H. Effects of building shade on photosynthesis and chlorophyll fluorescence of Euonymus fortunei. Acta Ecol. Sin. 2016, 36, 350–355. [Google Scholar] [CrossRef]
- Hitsuma, G.; Han, Q.; Chiba, Y. Photosynthesis and growth of Thujopsis dolabrata var. hondai seedlings in the understory of trees with various phenologies. J. For. Res. 2017, 17, 156–163. [Google Scholar] [CrossRef]
- Allen, J.F. Cyclic. pseudocyclic and noncyclic photophosphorylation: New links in the chain. Trends Plant Sci. 2003, 8, 15–19. [Google Scholar] [CrossRef]
- Dietzel, L.; Bräutigam, K.; Pfannschmidt, T. Photosynthetic acclimation: State transitions and adjustment of photosystem stoichiometry—Functional relationships between short-term and long-term light quality acclimation in plants. FEBS J. 2008, 275, 1080–1088. [Google Scholar] [CrossRef]
- Schreier, T.B.; Hibberd, J.M. Variations in the Calvin-Benson cycle: Selection pressures and optimization? J. Exp. Bot. 2019, 70, 1697–1701. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A. The cost of photoinhibition. Physiol. Plantarum. 2011, 142, 87–104. [Google Scholar] [CrossRef]
- Kitao, M.; Yoneda, R.; Tobita, H.; Matsumoto, Y.; Maruyama, Y.; Arifin, A.; Asan, A.; Muhamad, M. Susceptibility to photoinhibition in seedlings of six tropical fruit tree species native to Malasia following transplantation to a degraded land. Trees 2006, 20, 601–610. [Google Scholar] [CrossRef]
- Krause, G.H.; Koroleva, O.Y.; Dalling, W.; Winter, K. Acclimation of tropical tree seedlings to excessive light in simulated tree-fall gaps. Plant Cell Environ. 2001, 24, 1345–1352. [Google Scholar] [CrossRef] [Green Version]
- Fankhauser, C.; Batschauer, A. Shadow on the Plant: A Strategy to Exit. Cell 2016, 164, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Zu, Y.G.; Wei, X.X.; Yu, J.H.; Li, D.W.; Pang, H.H.; Tong, L. Responses in the physiology and biochemistry of Korean pine (Pinus koraiensis) under supplementary UV-B radiation. Photosynthetica 2011, 49, 448. [Google Scholar] [CrossRef]
- Li, X.N.; Wang, Y.H.; Yang, Z.H.; Liu, T.; Mu, C.C. Photosynthesis adaption in Korean pine to gap size and position within Populus davidiana forests in Xiaoxing’anling, China. J. For. Res. 2022, 33, 1517–1527. [Google Scholar] [CrossRef]
- Wang, P.; Grimm, B. Connecting chlorophyll metabolism with accumulation of the photosynthetic apparatus. Trends Plant Sci. 2021, 26, 484–495. [Google Scholar] [CrossRef]
- Osmond, C.B. What is photoinhibition? Some insights from comparison of shade and sun plants. In Photoinhibition of Photosynthesis: From Molecular Mechanisms to the Field; Baker, N.R., Bowyer, J.R., Eds.; BIOS Scientific Publishers: Oxford, UK, 1994; pp. 1–24. [Google Scholar]
- Favaretto, V.F.; Martinez, C.A.; Soriani, H.H.; Furriel, R.P. Differential responses of antioxidant enzymes in pioneer and late-successional tropical tree species grown under sun and shade conditions. Environ. Exp. Bot. 2011, 70, 20–28. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Potters, G.; Horemans, N.; Jansen, M.A.K. The cellular redox state in plant stress biology—A charging concept. Plant Physiol. Bioch. 2010, 48, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Fen, Y.; Hai, L. Effects of oxidative stress on growth and development of suspension cells of Populus tomentosa by transcriptome analysis. J. Beijing For. Univ. 2019, 41, 90–98. [Google Scholar]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Stirbet, A.; Govindjee. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photoch. Photobio. B 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Aranda, I.; Castro, L.; AlÍa, R.; Pardos, J.A.; Gil, L. Low temperature during winter elicits differential responses among populations of the Mediterranean evergreen cork oak (Quercus suber). Tree Physiol. 2005, 25, 1085–1090. [Google Scholar] [CrossRef] [Green Version]
- Öquist, G.; Wass, R. A portable, microprocessor operated instrument for measuring chlorophyll fluorescence kinetics in stress physiology. Physiol. Plantarum. 1988, 73, 211–217. [Google Scholar] [CrossRef]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural carbon in woody plants. Annu. Rev. Plant Biol. 2014, 65, 667–687. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Carbone, M.S.; Huggett, B.A.; Furze, M.E.; Czimczik, C.I.; Walker, J.C.; Xu, X.M.; Schaberg, P.G.; Murakami, P. Distribution and mixing of old and new nonstructural carbon in two temperate trees. New Phytol. 2015, 206, 590–597. [Google Scholar] [CrossRef] [Green Version]
- Xing, K.X.; Zhao, M.F.; Niinemets, Ü.; Niu, S.L.; Tian, J.; Jiang, Y.; Chen, H.Y.H.; White, P.J.; Guo, D.L.; Ma, Z.Q. Relationships between leaf carbon and macronutrients across woody species and forest ecosystems highlight how carbon is allocated to leaf structural function. Front. Plant Sci. 2021, 12, 674932. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.F.; Wu, W.H. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2021, 63, 34–52. [Google Scholar] [CrossRef]
- Li, J.Q.; Li, J.W. Regeneration and restoration of broad-leaved Korean pine forests in Lesser Xing’an Mountains of Northeast China. Acta Ecol. Sin. 2003, 23, 1268–1277. [Google Scholar]
- Shen, H.L.; Zhang, Q.; Fan, S.H.; Zhao, K.Z.; Yang, W.H. Influence of community structural characteristics of natural secondary forest on the growth of the young trees of Pinus koraiensis. For. Res. 2004, 17, 610–615. (In Chinese) [Google Scholar]
- Shen, H.L.; Cong, J.; Zhang, P.; Zhang, Q.; Fang, S.H.; Yang, W.H.; Liu, S.R. Effect of opening degree regulation on diameter and height increment and aboveground biomass of Korean pine trees planted under secondary forest. Chin. J. Appl. Ecol. 2011, 22, 2781–2791. (In Chinese) [Google Scholar]
- Cong, J.; Shen, H.L.; Yang, W.H.; Fan, S.H.; Zhang, Q. Effect of microenvironmental quantitative regulation on growth of Korean pine trees planted under secondary forest. J. For. Res. 2011, 22, 175–181. [Google Scholar] [CrossRef]
- Chen, X.W.; Li, B.L.; Lin, Z.S. The acceleration of succession for the restoration of the mixed-broadleaved Korean pine forests in Northeast China. For. Ecol. Manag. 2003, 177, 503–514. [Google Scholar] [CrossRef]
- Shen, H.L.; Wang, L.; Lin, C.X.; Cong, J.; Yang, W.H.; Zhang, P.; Zhang, Q.; Fang, S.H. Effect of community structure regulation on upper-layer broad-leaved trees in a mixed forest of planted Korean pine and naturally-regenerated broad-leaved trees. Sci. Silvae Sin. 2014, 50, 22–30. (In Chinese) [Google Scholar]
- Zhang, M.; Zhu, J.J.; Li, M.C.; Zhang, G.Q.; Yan, Q.L. Different light acclimation strategies of two coexisting tree species seedlings in a temperate secondary forest along five natural light levels. For. Ecol. Manag. 2013, 306, 234–242. [Google Scholar] [CrossRef]
- Li, Y.X.; Zhang, X.X.; Cai, K.W.; Zhang, Q.H.; Jiang, L.P.; Li, H.X.; Lv, Y.Z.; Qu, G.Z.; Zhao, X. Y Comparative transcriptomic and metabolic analyses reveal the coordinated mechanisms in Pinus koraiensis under different light stress conditions. Int. J. Mol. Sci. 2022, 23, 9556. [Google Scholar] [CrossRef]
- Makoto, K.; Koike, T. Effects of nitrogen supply on photosynthetic and anatomical changes in current-year needles of Pinus koraiensis seedlings grown under two irradiances. Photosynthetica 2007, 45, 99–104. [Google Scholar] [CrossRef]
- Sun, Y.R.; Zhu, J.J.; Sun, O.J.X.; Yan, Q.L. Photosynthetic and growth responses of Pinus koraiensis seedlings to canopy openness: Implications for the restoration of mixed-broadleaved Korean pine forests. Environ. Exp. Bot. 2016, 129, 118–126. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, Q.J.; Xu, Z.Z.; Du, W.X.X.; Yu, J.; Meng, S.W.; Zhou, H.; Qin, L.H.; Shah, S. How can the shade intolerant Korean pine survive under dense deciduous canopy? Forest Ecol. Manag. 2020, 457, 117735. [Google Scholar] [CrossRef]
- Zhu, J.J.; Wang, K.; Sun, Y.R.; Yan, Q.L. Response of Pinus koraiensis seedling growth to different light conditions based on the assessment of photosynthesis in current and one-year-old needles. J. For. Res. 2014, 25, 53–62. [Google Scholar] [CrossRef]
- Li, J.; Ding, D.X.; Li, N.H.; Xie, J.M.; Yu, J.H.; Lyv, J.; Bakpa, E.P.; Zhang, J.; Wang, C.; Zhang, J.F. Melatonin enhances the low-temperature combined low-light tolerance of pepper (Capsicum annuum L.) seedlings by regulating photosynthesis, carotenoid, and hormone metabolism. Environ. Exp. Bot. 2022, 199, 104868. [Google Scholar] [CrossRef]
- Binkley, D.; Adams, M.; Fredericksen, T.; Laclau, J.P.; Mäkinen, H.H.; Prescott, C. Editors’ note. For. Ecol. Manag. 2015, 349, 1–3. [Google Scholar] [CrossRef]
- Yuan, D.Y.; Zhu, L.J.; Cherubini, P.; Li, Z.S.; Zhang, Y.D.; Wang, X.C. Species-specific indication of 13 tree species growth on climate warming in temperate forest community of northeast China. Ecol. Indic. 2021, 133, 108389. [Google Scholar] [CrossRef]
- Wu, H.B.; Yin, D.S.; Salomón, R.L.; Rodríguez-Calcerrada, J.; Zhang, J.Y.; Zhang, P.; Shen, H.L. Cone-bearing branches of Pinus koraiensis are not carbon autonomous during cone development. Forests 2021, 12, 1257. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.J.; Gu, J.C.; Yu, L.Z.; Wang, Z.Q. Changes in soil phosphorus fractions after 9 years of continuous nitrogen addition in a Larix gmelinii plantation. Ann. For. Sci. 2014, 72, 435–442. [Google Scholar] [CrossRef]
- He, J.; Xü, X.; Li, S.H.; Mi, H.L.; Zhang, Y.P.; Zhao, T.C.; Ma, Y.M. Effects of water stress on photosynthetic pigment in leaves and chlorophyll fluorescence of Cynanchum komarovii. Acta Bot. Boreali-Occident. Sin. 2004, 24, 1594–1598. (In Chinese) [Google Scholar]
- Hansen, J.; Møller, I.B. Percolation of starch and soluble carbohydrates from plant tissue for quantitative determination with anthrone. Anal. Biochem. 1975, 68, 87–94. [Google Scholar] [CrossRef]
- Xu, S.; Chen, M.; Feng, T.; Zhan, L.; Zhou, L.; Yu, G. Use ggbreak to effectively utilize plotting space to deal with large datasets and outliers. Front. Genet. 2021, 12, 774–846. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, U. The Architecture of Plant Crowns: From Design Rules to Light Capture and Performance. In Handbook of Functional Plant Ecology, 2nd ed.; Pugnaire, F.I., Valladares, F., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 101–149. [Google Scholar]
- Consrabel, A.J.; Lieffers, V.J. Seasonal patterns of light transmission through boreal mixedwood canopies. Can. J. For. Res 1996, 26, 1008–1014. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.F.; Chen, C.; Jin, Z.X.; Yang, Z.N.; Li, Y.L. Leaf anatomy, photosynthesis, and chloroplast ultrastructure of Heptacodium miconioides seedlings reveal adaptation to light environment. Environ. Exp. Bot. 2022, 195, 104780. [Google Scholar] [CrossRef]
- Valladares, F.; Arrieta, S.; Aranda, I.; Lorenzo, D.; Sánchez-Gómez, D.; Tena, D.; Suárez, F.; Pardos, J.A. Shade tolerance, photoinhibition sensitivity and phenotypic plasticity of Ilex aquifolium in continental Mediterranean sites. Tree Physiol. 2005, 25, 1041–1052. [Google Scholar] [CrossRef] [Green Version]
- Griffin, J.J.; Ranney, T.G.; Pharr, D.M. Photosynthesis, chlorophyll fluorescence, and carbohydrate concentration of Illicium taxa grown under varied irradiance. J. Amer. Soc. Hort. Sci. 2004, 129, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Fanciullino, A.L.; Bidel, L.P.; Urban, L. Carotenoid responses to environmental stimuli: Integrating redox and carbon controls into a fruit model. Plant Cell Environ. 2014, 37, 273–289. [Google Scholar] [CrossRef]
- Carmo-Silva, E.; Scales, J.C.; Madgwick, P.J.; Parry, M.A. Optimizing Rubisco and its regulation for greater resource use efficiency. Plant Cell Environ. 2015, 38, 1817–1832. [Google Scholar] [CrossRef]
- Banks, J.M. Continuous excitation chlorophyll fluorescence parameters: A review for practitioners. Tree Physiol. 2017, 37, 1128–1136. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Linkosalo, T.; Heikkinen, J.; Pulkkinen, P.; Mäkipää, R. Fluorescence measurements show stronger cold inhibition of photosynthetic light reactions in Scots pine compared to Norway spruce as well as during spring compared to autumn. Front. Plant Sci. 2014, 5, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruban, A.V. Nonphotochemical chlorophyll fluorescence quenching: Mechanism and effectiveness in protecting plants from photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyka, T.; Robakowski, P.; Żytkowiak, R. Acclimation of leaves to contrasting irradiance in juvenile trees differing in shade tolerance. Tree Physiol. 2007, 27, 1293–1306. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Y.; Zhang, Q.H.; Shuang, S.P.; Cun, Z.; Wu, H.M.; Chen, J.W. The responses of light reaction of photosynthesis to dynamic sunflecks in a typically shade-tolerant species Panax notoginseng. Front. Plant Sci. 2021, 12, 718981. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees—From what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Zhang, Z.; Jin, G.; Liu, Z. Variations of leaf nonstructural carbohydrates in an evergreen coniferous species: Needle age and phenology dominate over life history. Ecol. Indic. 2022, 136, 108685. [Google Scholar] [CrossRef]
- Xu, M.L.; Hu, T.Q.; Poethig, R.S.S. Low light intensity delays vegetative phase change. Plant Physiol. 2021, 187, 1177–1188. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2019, 251, 3. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.L.; Pearcy, R.W. The importance of sunflecks for forest understory plants. BioScience 1991, 41, 760–766. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Breusegem, F.V. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Marler, T.E.; Krishnapillai, M.V. Incident light and leaf age influence leaflet element concentrations of Cycas micronesica trees. Horticulturae 2019, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Patra, J.K.; Thatoi, H. Antioxidative response to abiotic and biotic stresses in mangrove plants: A review. Int. Rev. Hydrobiol. 2016, 101, 3–19. [Google Scholar] [CrossRef]
- Shafi, A.; Zahoor, I.; Mushtaq, U. Proline Accumulation and Oxidative Stress: Diverse roles and Mechanism of Tolerance and Adaptation under Salinity Stress. Salt Stress, Microbes, and Plant Interactions: Mechanisms and Molecular Approaches; Springer: Berlin/Heidelberg, Germany, 2019; Volume 2, pp. 269–300. [Google Scholar]
- Forlani, G.; Trovato, M.; Funck, D.; Signorelli, S. Regulation of proline accumulation and its molecular and physiological functions in stress defence. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants: Recent Advances and Future Perspectives; Springer: Berlin/Heidelberg, Germany, 2019; pp. 73–97. [Google Scholar]
- Rajendrakumar, C.S.V.; Reddy, B.V.B.; Reddy, A.R. Proline-protein interactions: Protection of structural and functional integrity of M4 lactate dehydrogenase. Biochem. Biophys. Res. Commun. 1994, 201, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Dubey, R.S. Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water stress: Role of osmolytes as enzyme protectant. J. Plant Physiol. 2005, 162, 854–864. [Google Scholar] [CrossRef]
- Mishra, S.; Dubey, R.S. Inhibition of ribonuclease and protease activities in arsenic exposed rice seedlings: Role of proline as enzyme protectant. J. Plant Physiol. 2006, 163, 927–936. [Google Scholar] [CrossRef]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Matysik, J.; Bhalu, A.B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 82, 525–532. [Google Scholar]
- Wang, F.J.; Zeng, B.; Sun, Z.X.; Zhu, C. Relationship between proline and Hg2+-induced oxidative stress in a tolerant rice mutant. Arch. Environ. Contam. Toxicol. 2009, 56, 723–731. [Google Scholar] [CrossRef]
- Alia, P.S.; Mohanty, P. Involvement of proline in protecting thylakoid membranes against free radical-induced photodamage. J. Photochem. Photobiol. 1997, 38, 253–257. [Google Scholar] [CrossRef]
- Yang, C.W.; Li, L. Hormonal regulation in shade avoidance. Front. Plant Sci. 2017, 8, 1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagnola, J.I.; Ploschuk, E.; Benech-Arnold, T.; Finlayson, S.A.; Casal, J.J. Stem transcriptome reveals mechanisms to reduce the energetic cost of shade-avoidance responses in tomato. Plant Physiol. 2012, 160, 1110–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klem, K.; Gargallo-Garriga, A.; Rattanapichai, W.; Oravec, M.; Holub, P.; Vesela, B.; Sardans, J.; Penuelas, J.; Urban, O. Distinct morphological, physiological, and biochemical responses to light quality in barley leaves and roots. Front. Plant Sci. 2019, 10, 1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Dai, J.; Ge, Q. Responses of autumn phenology to climate change and the correlations of plant hormone regulation. Sci. Rep. 2020, 10, 9039. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2009, 15, 89–97. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Light Conditions | Sampling Time | ||
|---|---|---|---|---|
| June | July | September | ||
| Chl a (mg g−1 FW) | Low | 0.796 ± 0.041 Ab | 1.154 ± 0.157 Aa | 1.006 ± 0.018 Aab |
| Moderate | 0.485 ± 0.028 Bb | 0.860 ± 0.095 Aa | 0.961 ± 0.043 Aa | |
| High | 0.533 ± 0.044 Bb | 1.062 ± 0.077 Aa | 0.932 ± 0.135 Aa | |
| Full | 0.302 ± 0.015 Cc | 0.498 ± 0.006 Bb | 0.859 ± 0.045 Aa | |
| Chl b (mg g−1 FW) | Low | 0.210 ± 0.009 Ab | 0.347 ± 0.038 Aa | 0.271 ± 0.007 Aab |
| Moderate | 0.131 ± 0.003 Bb | 0.253 ± 0.027 Ba | 0.257 ± 0.014 Aa | |
| High | 0.144 ± 0.020 Ba | 0.292 ± 0.027 ABa | 0.288 ± 0.072 Aa | |
| Full | 0.067 ± 0.002 Cc | 0.122 ± 0.006 Cb | 0.186 ± 0.010 Aa | |
| Car (mg g−1 FW) | Low | 0.242 ± 0.008 Ab | 0.415 ± 0.062 Aa | 0.398 ± 0.006 Aa |
| Moderate | 0.170 ± 0.012 Bc | 0.310 ± 0.026 ABb | 0.382 ± 0.012 Aa | |
| High | 0.190 ± 0.016 Bb | 0.381 ± 0.030 Aa | 0.389 ± 0.054 Aa | |
| Full | 0.108 ± 0.005 Cc | 0.196 ± 0.003 Bb | 0.345 ± 0.010 Aa | |
| Car/Chl | Low | 0.241 ± 0.003 Bc | 0.275 ± 0.011 Bb | 0.312 ± 0.013 Aa |
| Moderate | 0.276 ± 0.006 Ab | 0.280 ± 0.012 Bb | 0.315 ± 0.025 Aa | |
| High | 0.282 ± 0.005 Ab | 0.281 ± 0.005 Bb | 0.322 ± 0.019 Aa | |
| Full | 0.293 ± 0.006 Aa | 0.316 ± 0.002 Aa | 0.332 ± 0.036 Aa | |
| Fv/Fm | Low | 0.865 ± 0.004 Aa | 0.826 ± 0.004 Cb | 0.870 ± 0.008 Aa |
| Moderate | 0.863 ± 0.003 Aa | 0.866 ± 0.002 Aa | 0.873 ± 0.005 Aa | |
| High | 0.870 ± 0.003 Aa | 0.869 ± 0.002 Aa | 0.850 ± 0.025 Aa | |
| Full | 0.833 ± 0.009 Ba | 0.840 ± 0.006 Ba | 0.856 ± 0.011 Aa | |
| NPQ | Low | 2.663 ± 0.114 Bb | 2.333 ± 0.073 Bc | 3.672 ± 0.050 aB |
| Moderate | 3.131 ± 0.231 ABa | 3.408 ± 0.125 Aa | 3.452 ± 0.326 Ba | |
| High | 3.055 ± 0.205 ABa | 3.023 ± 0.298 Aa | 3.237 ± 0.495 Ba | |
| Full | 3.428 ± 0.277 Ab | 3.501 ± 0.062 Ab | 4.827 ± 0.199 Aa | |
| Rubisco activity (U g−1 FW) | Low | 0.575 ± 0.023 Ba | 0.497 ± 0.005 Ba | 0.534 ± 0.048 Ba |
| Moderate | 0.663 ± 0.049 ABa | 0.545 ± 0.029 Ba | 0.543 ± 0.059 Ba | |
| High | 0.688 ± 0.031 ABa | 0.697 ± 0.019 Aa | 0.635 ± 0.051 ABa | |
| Full | 0.716 ± 0.044 Aa | 0.702 ± 0.010 Aa | 0.733 ± 0.013 Aa | |
| Parameters | Light Conditions | Sampling Stages | ||
|---|---|---|---|---|
| June | July | September | ||
| SOD (U g−1 FW) | Low | 44 ± 18 Ba | 49 ± 0 Bb | 77 ± 1 Ab |
| Moderate | 66 ± 19 Ba | 49 ± 1 Bb | 46 ± 2 Bb | |
| High | 67 ± 40 Aa | 55 ± 6 bB | 69 ± 2 Ab | |
| Full | 100 ± 39 Aa | 247 ± 13 Aa | 76 ± 5 Ab | |
| POD (U g−1 FW) | Low | 353 ± 18 Ac | 1300 ± 76 ABb | 2527 ± 137 Ba |
| Moderate | 287 ± 18 Ac | 1078 ± 118 Bb | 2567 ± 188 Ba | |
| High | 380 ± 80 Ac | 1787 ± 264 Ab | 3573 ± 77 Aa | |
| Full | 370 ± 38 Ac | 832 ± 65 Bb | 2907 ± 29 Ba | |
| APX (U g−1 FW) | Low | 592 ± 33 Ba | 686 ± 34 Ba | 440 ± 53 Bb |
| Moderate | 682 ± 38 Bab | 796 ± 39 ABa | 516 ± 92 Bb | |
| High | 690 ± 32 Bb | 870 ± 70 Aa | 620 ± 35 ABb | |
| Full | 802 ± 25 Ab | 913 ± 22 Aa | 785 ± 34 Ab | |
| GSH-PX (U g−1 FW) | Low | 1.29 ± 0.07 ABb | 1.68 ± 0.04 Aa | 0.61 ± 0.08 Bc |
| Moderate | 1.23 ± 0.09 ABb | 1.63 ± 0.04 Aa | 0.67 ± 0.09 Bc | |
| High | 1.42 ± 0.05 Aa | 1.37 ± 0.07 Ba | 1.01 ± 0.07 Ab | |
| Full | 1.05 ± 0.11 Bab | 1.32 ± 0.03 Ba | 0.98 ± 0.08 Ab | |
| DPPH (%) | Low | 27.16 ± 3.32 Cb | 45.39 ± 2.30 Ba | 50.70 ± 5.51 Ba |
| Moderate | 44.35 ± 0.57 Bb | 49.84 ± 6.22 Bab | 58.27 ± 1.09 Ba | |
| High | 44.63 ± 3.73 Bb | 55.02 ± 4.67 Bab | 61.95 ± 4.96 Ba | |
| Full | 85.59 ± 1.95 Aa | 88.71 ± 0.20 Aa | 87.86 ± 1.71 Aa | |
| Variables | Different Light Factors at Three Sampling Stages | |||||
|---|---|---|---|---|---|---|
| PPFD | R/FR | |||||
| June | July | September | June | July | September | |
| Chl a | −0.723 ** | −0.791 ** | −0.382 | −0.813 ** | −0.708 * | −0.358 |
| Chlb | −0.777 ** | −0.833 ** | −0.558 | −0.842 ** | −0.781 ** | −0.205 |
| Car | −0.798 ** | −0.756 ** | −0.436 | −0.839 ** | −0.673 * | −0.247 |
| Car/Chl | 0.576 * | 0.911 ** | 0.276 | 0.740 ** | 0.887 ** | 0.305 |
| Fv/Fm | −0.878 ** | −0.302 | −0.177 | −0.801 ** | −0.117 | −0.310 |
| NPQ | 0.516 | 0.492 | 0.780 ** | 0.483 | 0.501 | 0.187 |
| Rubisco | 0.424 | 0.569 | 0.690 * | 0.562 | 0.753 ** | 0.684 * |
| Glucose | 0.937 ** | 0.977 ** | 0.794 ** | 0.880 ** | 0.917 ** | 0.219 |
| Starch | 0.845 ** | 0.178 | −0.692 * | 0.823 ** | 0.105 | −0.466 |
| Proline | 0.440 | 0.524 | 0.718 ** | 0.481 | 0.651 * | 0.850 ** |
| ABA | −0.309 | −0.566 | 0.749 ** | −0.324 | −0.682 * | 0.792 ** |
| SOD | 0.78 ** | 0.987 ** | 0.375 | 0.81 ** | 0.959 ** | −0.007 |
| POD | 0.181 | −0.580 * | 0.045 | 0.162 | −0.350 | 0.655 * |
| APX | 0.737 ** | 0.523 | 0.747 ** | 0.761 ** | 0.662 * | 0.718 ** |
| GSH-PX | −0.638 * | −0.625 * | 0.475 | −0.538 | −0.786 ** | 0.863 ** |
| DPPH | 0.869 ** | 0.931 ** | 0.897 ** | 0.865 ** | 0.930 ** | 0.700 * |
| C/N | 0.982 ** | 0.828 ** | 0.840 ** | 0.980 ** | 0.845 ** | 0.846 ** |
| P | 0.710 ** | −0.629 * | −0.401 | 0.685 * | −0.712 ** | −0.800 ** |
| K | −0.962 ** | −0.920 ** | −0.894 ** | −0.821 ** | −0.893 ** | −0.697 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Li, B.; Ma, X.; Saha, S.; Wu, H.; Zhang, P.; Shen, H. Physiological and Biochemical Traits of Needles Imply That Understory Light Conditions in the Growing Season May Be Favorable to Pinus koraiensis Trees. Forests 2023, 14, 1333. https://doi.org/10.3390/f14071333
Li W, Li B, Ma X, Saha S, Wu H, Zhang P, Shen H. Physiological and Biochemical Traits of Needles Imply That Understory Light Conditions in the Growing Season May Be Favorable to Pinus koraiensis Trees. Forests. 2023; 14(7):1333. https://doi.org/10.3390/f14071333
Chicago/Turabian StyleLi, Wenkai, Bei Li, Xiao Ma, Sudipta Saha, Haibo Wu, Peng Zhang, and Hailong Shen. 2023. "Physiological and Biochemical Traits of Needles Imply That Understory Light Conditions in the Growing Season May Be Favorable to Pinus koraiensis Trees" Forests 14, no. 7: 1333. https://doi.org/10.3390/f14071333