
The Effects of Korean Pine and Manchurian Walnut Monocultures and Mixed Plantations on Soil Fungal and Bacterial Communities

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Soil Sampling
2.3. Soil Physical and Chemical Analysis
2.4. DNA Extraction and PCR Amplification
2.5. Illumina Sequencing and Sequence Data Processing
2.6. Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
3.2. The Sequencing Characteristics and Community Composition of Soil Fungi and Bacteria
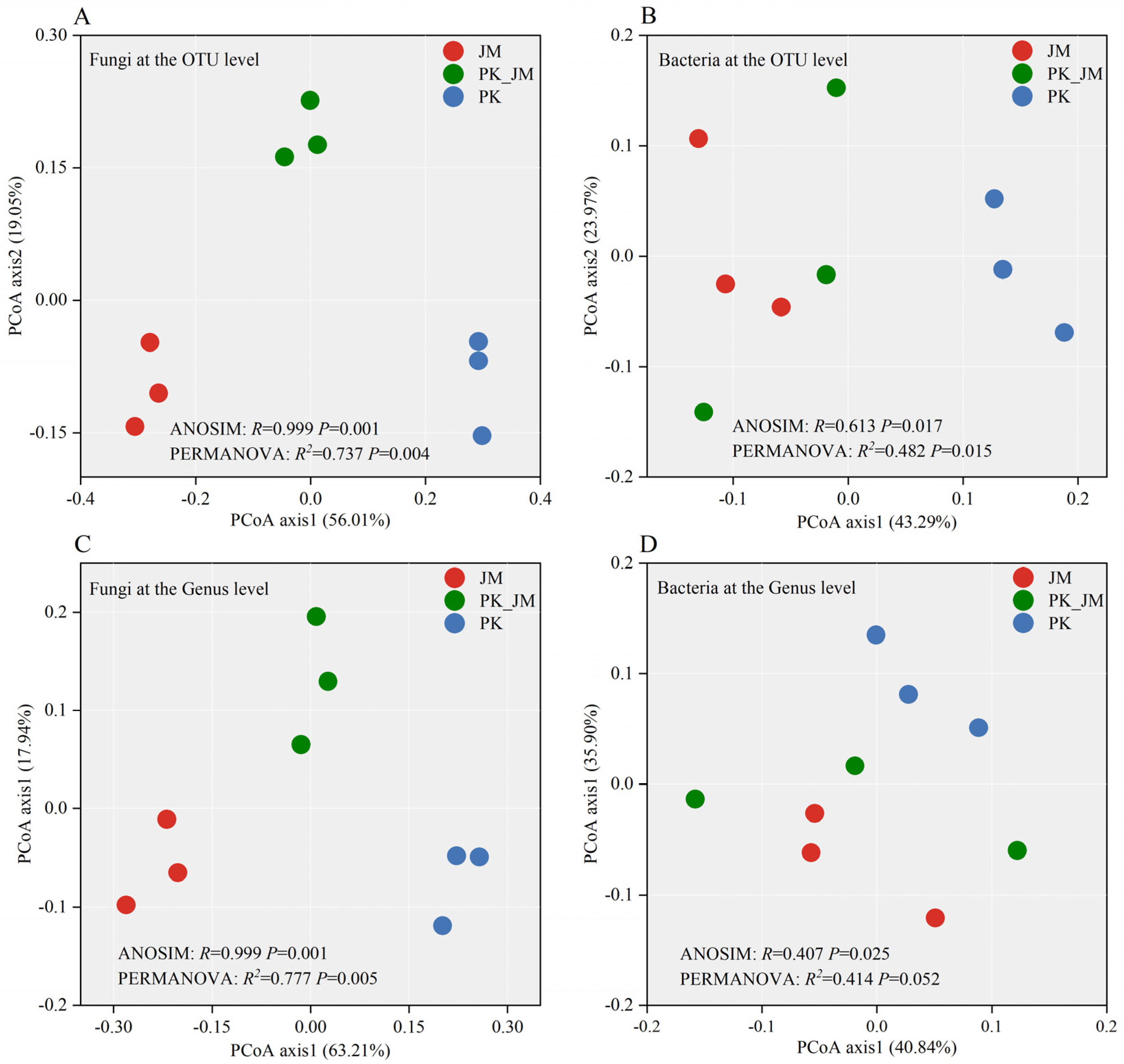
3.3. Fungal and Bacterial Alpha and Beta Diversity
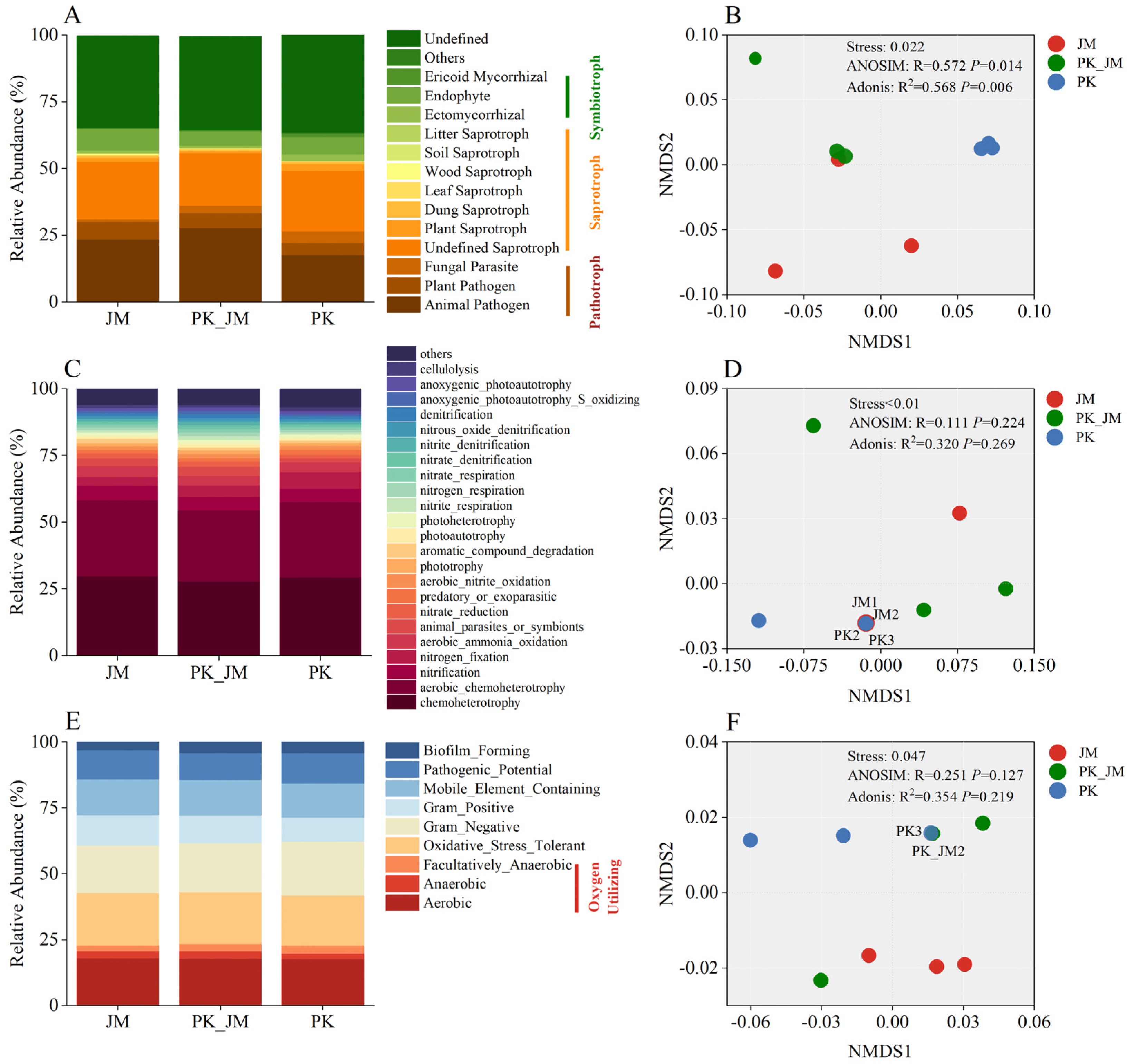
3.4. Fungal and Bacterial Functional Groups
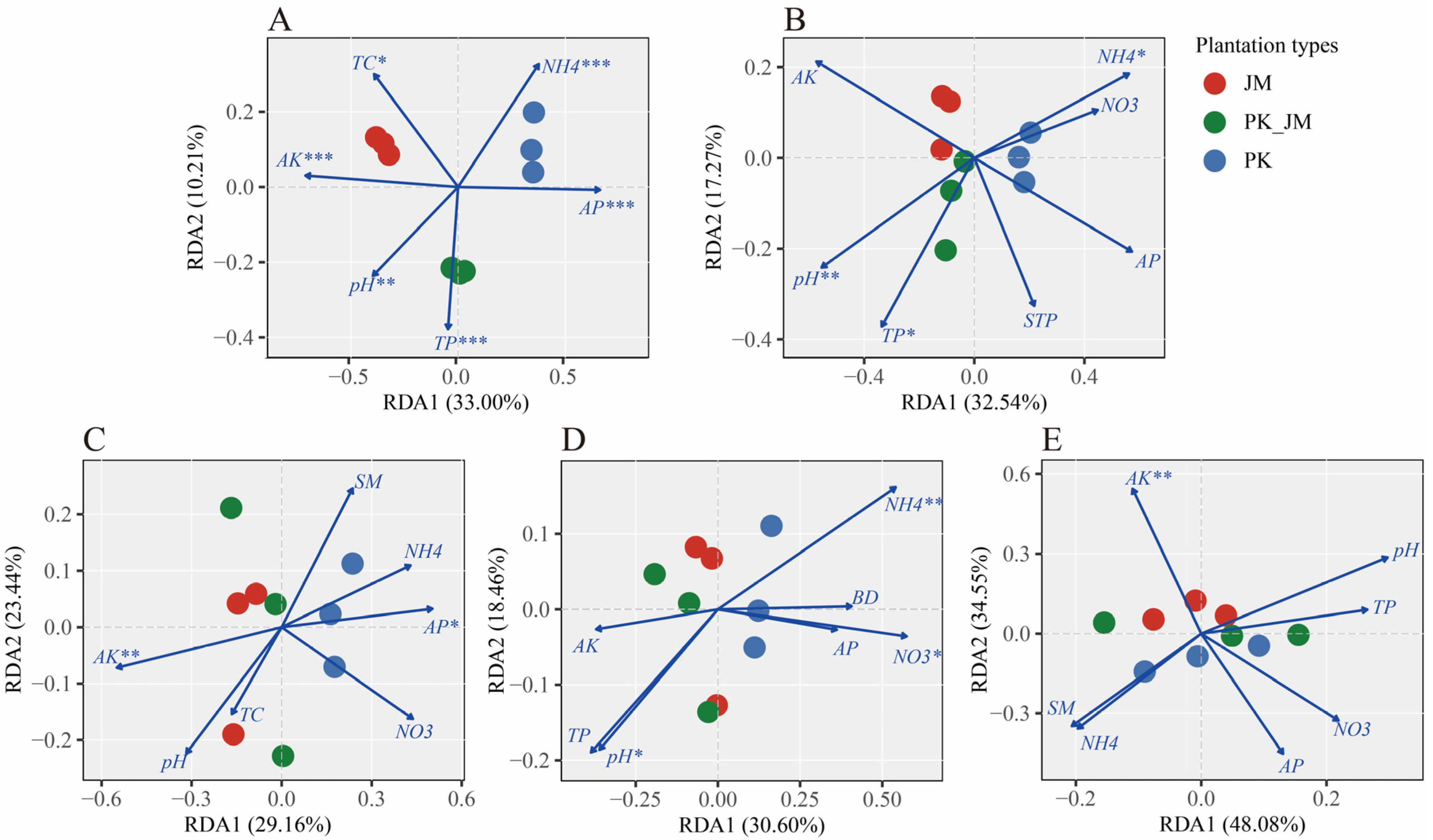
3.5. The Correlations between Soil Variables and Microbial Community Structure
3.6. The Patterns of Fungal and Bacterial Co-Occurrence Networks
4. Discussion
4.1. Influence of Plantation Type in Shaping Fungal and Bacterial Diversity and Composition
4.2. Changes in Functional Group Structures of Fungal and Bacterial Communities across Different Plantation Types
4.3. Soil Factors Play a Key Role in Shaping Soil Microbial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, X.; Li, B.-L.; Lin, Z.-S. The acceleration of succession for the restoration of the mixed-broadleaved Korean pine forests in Northeast China. For. Ecol. Manag. 2003, 177, 503–514. [Google Scholar] [CrossRef]
- Harris, J. Soil Microbial Communities and Restoration Ecology: Facilitators or Followers? Science 2009, 325, 573–574. [Google Scholar] [CrossRef]
- Bastida, F.; Eldridge, D.J.; García, C.; Kenny Png, G.; Bardgett, R.D.; Delgado-Baquerizo, M. Soil microbial diversity–biomass relationships are driven by soil carbon content across global biomes. ISME J. 2021, 15, 2081–2091. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhang, Y.; Hu, X.-F.; Wan, S.; Wang, H.; Liang, C.; Chen, F.-S. Litter manipulation effects on microbial communities and enzymatic activities vary with soil depth in a subtropical Chinese fir plantation. For. Ecol. Manag. 2021, 480, 118641. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Chodak, M.; Klimek, B.; Azarbad, H.; Jaźwa, M. Functional diversity of soil microbial communities under Scots pine, Norway spruce, silver birch and mixed boreal forests. Pedobiologia 2015, 58, 81–88. [Google Scholar] [CrossRef]
- Beugnon, R.; Du, J.; Cesarz, S.; Jurburg, S.D.; Pang, Z.; Singavarapu, B.; Wubet, T.; Xue, K.; Wang, Y.; Eisenhauer, N. Tree diversity and soil chemical properties drive the linkages between soil microbial community and ecosystem functioning. ISME Commun. 2021, 1, 41. [Google Scholar] [CrossRef]
- He, D.; Shen, W.; Eberwein, J.; Zhao, Q.; Ren, L.; Wu, Q.L. Diversity and co-occurrence network of soil fungi are more responsive than those of bacteria to shifts in precipitation seasonality in a subtropical forest. Soil Biol. Biochem. 2017, 115, 499–510. [Google Scholar] [CrossRef]
- Manzoni, S. Flexible Carbon-Use Efficiency across Litter Types and during Decomposition Partly Compensates Nutrient Imbalances—Results from Analytical Stoichiometric Models. Front. Microbiol. 2017, 8, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vesala, R.; Kiheri, H.; Hobbie, E.A.; van Dijk, N.; Dise, N.; Larmola, T. Atmospheric nitrogen enrichment changes nutrient stoichiometry and reduces fungal N supply to peatland ericoid mycorrhizal shrubs. Sci. Total Environ. 2021, 794, 148737. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, Q.; Sun, X.; Chen, D.; Insam, H.; Koide, R.T.; Zhang, S. Effects of mixed-species litter on bacterial and fungal lignocellulose degradation functions during litter decomposition. Soil Biol. Biochem. 2020, 141, 107690. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef]
- Crowther, T.W.; van den Hoogen, J.; Wan, J.; Mayes, M.A.; Keiser, A.D.; Mo, L.; Averill, C.; Maynard, D.S. The global soil community and its influence on biogeochemistry. Science 2019, 365, eaav0550. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leewis, M.-C.; Lawrence, C.R.; Schulz, M.S.; Tfaily, M.M.; Ayala-Ortiz, C.O.; Flores, G.E.; Mackelprang, R.; McFarland, J.W. The influence of soil development on the depth distribution and structure of soil microbial communities. Soil Biol. Biochem. 2022, 174, 108808. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Luo, Y. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 3072. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.M.; Bruns, T.D.; Taylor, J.W.; Smith, D.P.; Branco, S.; Glassman, S.I.; Erlandson, S.; Vilgalys, R.; Liao, H.-L.; Smith, M.E.; et al. Endemism and functional convergence across the North American soil mycobiome. Proc. Natl. Acad. Sci. USA 2014, 111, 6341–6346. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Dal Bello, M.; Lee, H.; Goyal, A.; Gore, J. Resource–diversity relationships in bacterial communities reflect the network structure of microbial metabolism. Nat. Ecol. Evol. 2021, 5, 1424–1434. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Louca, S.; Parfrey Laura, W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Yin, C. Seasonal variations of soil fungal diversity and communities in subalpine coniferous and broadleaved forests. Sci. Total Environ. 2022, 846, 157409. [Google Scholar] [CrossRef]
- Siles, J.A.; Margesin, R. Seasonal soil microbial responses are limited to changes in functionality at two Alpine forest sites differing in altitude and vegetation. Sci. Rep. 2017, 7, 2204. [Google Scholar] [CrossRef] [Green Version]
- Yeung, A.C.Y.; Kreutzweiser, D.P.; Richardson, J.S. Stronger effects of litter origin on the processing of conifer than broadleaf leaves: A test of home-field advantage of stream litter breakdown. Freshw. Biol. 2019, 64, 1755–1768. [Google Scholar] [CrossRef]
- Sui, X.; Li, M.; Frey, B.; Wang, M.; Weng, X.; Wang, X.; Chen, F.; Li, X.; Du, Z.; Yang, L.; et al. Climax forest has a higher soil bacterial diversity but lower soil nutrient contents than degraded forests in temperate northern China. Ecol. Evol. 2022, 12, e9535. [Google Scholar] [CrossRef]
- Jiang, S.; Xing, Y.; Liu, G.; Hu, C.; Wang, X.; Yan, G.; Wang, Q. Changes in soil bacterial and fungal community composition and functional groups during the succession of boreal forests. Soil Biol. Biochem. 2021, 161, 108393. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, B.; Ma, Y.; Xu, J.; Sun, H. The response of the soil bacterial community and function to forest succession caused by forest disease. Funct. Ecol. 2020, 34, 2548–2559. [Google Scholar] [CrossRef]
- Pfeiffer, B.; Fender, A.-C.; Lasota, S.; Hertel, D.; Jungkunst, H.F.; Daniel, R. Leaf litter is the main driver for changes in bacterial community structures in the rhizosphere of ash and beech. Appl. Soil Ecol. 2013, 72, 150–160. [Google Scholar] [CrossRef]
- Mommer, L.; Cotton, T.E.A.; Raaijmakers, J.M.; Termorshuizen, A.J.; van Ruijven, J.; Hendriks, M.; van Rijssel, S.Q.; van de Mortel, J.E.; van der Paauw, J.W.; Schijlen, E.G.W.M.; et al. Lost in diversity: The interactions between soil-borne fungi, biodiversity and plant productivity. New Phytol. 2018, 218, 542–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Feng, J.; Ao, G.; Qin, W.; Han, M.; Shen, Y.; Liu, M.; Chen, Y.; Zhu, B. Globally nitrogen addition alters soil microbial community structure, but has minor effects on soil microbial diversity and richness. Soil Biol. Biochem. 2023, 179, 108982. [Google Scholar] [CrossRef]
- Peng, W.; Zhu, Y.; Song, M.; Du, H.; Song, T.; Zeng, F.; Zhang, F.; Wang, K.; Luo, Y.; Lan, X.; et al. The spatial distribution and drivers of soil microbial richness and diversity in a karst broadleaf forest. For. Ecol. Manag. 2019, 449, 117241. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, Y.; Xiao, L.; An, S. Climate and soil properties regulate soil fungal communities on the Loess Plateau. Pedobiologia 2020, 81–82, 150668. [Google Scholar] [CrossRef]
- Beimforde, C.; Feldberg, K.; Nylinder, S.; Rikkinen, J.; Tuovila, H.; Dörfelt, H.; Gube, M.; Jackson, D.J.; Reitner, J.; Seyfullah, L.J.; et al. Estimating the Phanerozoic history of the Ascomycota lineages: Combining fossil and molecular data. Mol. Phylogenetics Evol. 2014, 78, 386–398. [Google Scholar] [CrossRef]
- Yelle, D.J.; Ralph, J.; Lu, F.; Hammel, K.E. Evidence for cleavage of lignin by a brown rot basidiomycete. Environ. Microbiol. 2008, 10, 1844–1849. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Condron, L.M.; Dunfield, K.E.; Wakelin, S.A.; Chen, L. Impact of grassland afforestation with contrasting tree species on soil phosphorus fractions and alkaline phosphatase gene communities. Soil Biol. Biochem. 2021, 159, 108274. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y.H.; Chen, C.; Peng, S. Water availability regulates negative effects of species mixture on soil microbial biomass in boreal forests. Soil Biol. Biochem. 2019, 139, 107634. [Google Scholar] [CrossRef]
- Korboulewsky, N.; Heiniger, C.; De Danieli, S.; Brun, J.J. Effect of tree mixture on Collembola diversity and community structure in temperate broadleaf and coniferous forests. For. Ecol. Manag. 2021, 482, 118876. [Google Scholar] [CrossRef]
- Korboulewsky, N.; Perez, G.; Chauvat, M. How tree diversity affects soil fauna diversity: A review. Soil Biol. Biochem. 2016, 94, 94–106. [Google Scholar] [CrossRef]
- Li, S.; Huang, X.; Shen, J.; Xu, F.; Su, J. Effects of plant diversity and soil properties on soil fungal community structure with secondary succession in the Pinus yunnanensis forest. Geoderma 2020, 379, 114646. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Chang, S.X.; Xu, Q.; Li, Y.; Ma, Z.; Qin, H.; Cai, Y. Linking enhanced soil nitrogen mineralization to increased fungal decomposition capacity with Moso bamboo invasion of broadleaf forests. Sci. Total Environ. 2021, 771, 144779. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Pylro, V.S.; Baldrian, P.; van Elsas, J.D.; Salles, J.F. Ecological succession reveals potential signatures of marine–terrestrial transition in salt marsh fungal communities. ISME J. 2016, 10, 1984–1997. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Buée, M.; Deveau, A.; Mieszkin, S.; Martin, F. Ecology of the forest microbiome: Highlights of temperate and boreal ecosystems. Soil Biol. Biochem. 2016, 103, 471–488. [Google Scholar] [CrossRef]
- Wei, G.; Li, M.; Shi, W.; Tian, R.; Chang, C.; Wang, Z.; Wang, N.; Zhao, G.; Gao, Z. Similar drivers but different effects lead to distinct ecological patterns of soil bacterial and archaeal communities. Soil Biol. Biochem. 2020, 144, 107759. [Google Scholar] [CrossRef]
- Johnston, A.S.A.; Sibly, R.M. The influence of soil communities on the temperature sensitivity of soil respiration. Nat. Ecol. Evol. 2018, 2, 1597–1602. [Google Scholar] [CrossRef]
- Pugnaire, F.I.; Morillo, J.A.; Peñuelas, J.; Reich, P.B.; Bardgett, R.D.; Gaxiola, A.; Wardle, D.A.; van der Putten, W.H. Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems. Sci. Adv. 2019, 5, eaaz1834. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Cáceres, D.; Stokes, A.; Angeles-Alvarez, G.; Abadie, J.; Anthelme, F.; Bounous, M.; Freschet, G.T.; Roumet, C.; Weemstra, M.; Merino-Martín, L.; et al. Vegetation creates microenvironments that influence soil microbial activity and functional diversity along an elevation gradient. Soil Biol. Biochem. 2022, 165, 108485. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626. [Google Scholar] [CrossRef]
- Prescott, C.; Blevins, L.; Staley, C. Litter decomposition in B.C. forests: Controlling factors and influences of forestry activities. J. Ecosyst. Manag. 2005, 5, 44–57. [Google Scholar]
- Shihan, A.; Hättenschwiler, S.; Milcu, A.; Joly, F.-X.; Santonja, M.; Fromin, N. Changes in soil microbial substrate utilization in response to altered litter diversity and precipitation in a Mediterranean shrubland. Biol. Fertil. Soils 2017, 53, 171–185. [Google Scholar] [CrossRef]
- Whalen, E.D.; Lounsbury, N.; Geyer, K.; Anthony, M.; Morrison, E.; van Diepen, L.T.A.; Le Moine, J.; Nadelhoffer, K.; vanden Enden, L.; Simpson, M.J.; et al. Root control of fungal communities and soil carbon stocks in a temperate forest. Soil Biol. Biochem. 2021, 161, 108390. [Google Scholar] [CrossRef]
- Marklein, A.R.; Houlton, B.Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.P.; Jumpponen, A. Phylogenetic diversity analyses reveal disparity between fungal and bacterial communities during microbial primary succession. Soil Biol. Biochem. 2015, 89, 52–60. [Google Scholar] [CrossRef]
- Tu, Q.; Yan, Q.; Deng, Y.; Michaletz, S.T.; Buzzard, V.; Weiser, M.D.; Waide, R.; Ning, D.; Wu, L.; He, Z.; et al. Biogeographic patterns of microbial co-occurrence ecological networks in six American forests. Soil Biol. Biochem. 2020, 148, 107897. [Google Scholar] [CrossRef]
- Gao, C.; Xu, L.; Montoya, L.; Madera, M.; Hollingsworth, J.; Chen, L.; Purdom, E.; Singan, V.; Vogel, J.; Hutmacher, R.B.; et al. Co-occurrence networks reveal more complexity than community composition in resistance and resilience of microbial communities. Nat. Commun. 2022, 13, 3867. [Google Scholar] [CrossRef] [PubMed]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Variable | Plantation Type | ANOVA | |||
|---|---|---|---|---|---|
| JM | PK_JM | PK | F | p | |
| SM (%) | 27.31 ± 0.77 b | 40.18 ± 4.11 a | 39.52 ± 0.11 a | 8.98 | 0.016 * |
| BD (g·cm−3) | 0.78 ± 0.02 a | 0.70 ± 0.032 a | 0.78 ± 0.02 a | 3.35 | 0.105 |
| STP (%) | 75.54 ± 2.98 b | 86.03 ± 0.99 a | 83.17 ± 2.91 ab | 4.82 | 0.047 * |
| pH | 5.27 ± 0.08 a | 5.35 ± 0.09 a | 5.08 ± 0.01 b | 6.22 | 0.034 * |
| TC (g·kg−1) | 115.94 ± 0.66 a | 113.25 ± 0.31 b | 114.31 ± 0.35 b | 8.47 | 0.018 * |
| TN (g·kg−1) | 6.84 ± 0.24 a | 6.83 ± 0.09 a | 6.78 ± 0.10 a | 0.04 | 0.960 |
| TP (g·kg−1) | 0.84 ± 0.01 b | 1.15 ± 0.06 a | 0.81 ± 0.05 b | 17.34 | 0.003 ** |
| SOC (g·kg−1) | 60.71 ± 2.44 a | 62.98 ± 0.76 a | 63.36 ± 0.75 a | 0.887 | 0.460 |
| NO3−-N-N (mg·kg−1) | 38.98 ± 1.14 ab | 36.32 ± 1.47 b | 42.80 ± 0.94 a | 7.31 | 0.025 * |
| NH4+-N (mg·kg−1) | 5.31 ± 0.22 b | 3.88 ± 0.36 c | 7.14 ± 0.15 a | 40.59 | 0.000 *** |
| AP (mg·kg−1) | 12.14 ± 0.44 c | 16.85 ± 0.71 b | 22.80 ± 1.98 a | 18.61 | 0.003 ** |
| AK (mg·kg−1) | 140.38 ± 2.19 a | 120.83 ± 1.70 b | 105.55 ± 0.90 c | 107.71 | 0.000 *** |
| Network Features | Fungi | Bacteria | |
|---|---|---|---|
| Empirical network | Similarity threshold | 0.94 | 0.96 |
| Number of nodes | 387 | 685 | |
| Number of edges | 407 | 553 | |
| R2 of power law | 0.933 | 0.954 | |
| The percentage of positive correlation | 73.71% | 75.95% | |
| The percentage of negative correlation | 26.29% | 24.05% | |
| Average degree (AvgK) | 2.103 | 1.615 | |
| Average clustering coefficient (AvgCC) | 0.133 | 0.136 | |
| Average path distance (GD) | 8.339 | 7.394 | |
| Modularity | 0.880 | 0.951 | |
| Random network | Average clustering coefficient ± SD | 0.004 ± 0.003 | 0.002 ± 0.001 |
| Average path distance ± SD | 6.520 ± 0.248 | 9.044 ± 0.727 | |
| Modularity ± SD | 0.796 ± 0.008 | 0.927 ± 0.007 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, F.; Liu, N.; Wang, Y.; Liu, H.; Jia, H.; Yang, L. The Effects of Korean Pine and Manchurian Walnut Monocultures and Mixed Plantations on Soil Fungal and Bacterial Communities. Forests 2023, 14, 1594. https://doi.org/10.3390/f14081594
Shen F, Liu N, Wang Y, Liu H, Jia H, Yang L. The Effects of Korean Pine and Manchurian Walnut Monocultures and Mixed Plantations on Soil Fungal and Bacterial Communities. Forests. 2023; 14(8):1594. https://doi.org/10.3390/f14081594
Chicago/Turabian StyleShen, Fangyuan, Ning Liu, Yujiao Wang, Huifeng Liu, Haikuan Jia, and Lixue Yang. 2023. "The Effects of Korean Pine and Manchurian Walnut Monocultures and Mixed Plantations on Soil Fungal and Bacterial Communities" Forests 14, no. 8: 1594. https://doi.org/10.3390/f14081594