
Altitudinal Differentiation of Forest Resilience to Drought in a Dryland Mountain

Abstract
:1. Introduction
2. Methodology
2.1. Research Area
2.2. Data Source and Processing
2.2.1. Meteorological Data
2.2.2. Elevation Data
2.2.3. Vegetation Data
2.3. Analysis
2.3.1. Standardized Precipitation Evapotranspiration Index
2.3.2. Theoretical Basis of Resilience Curve
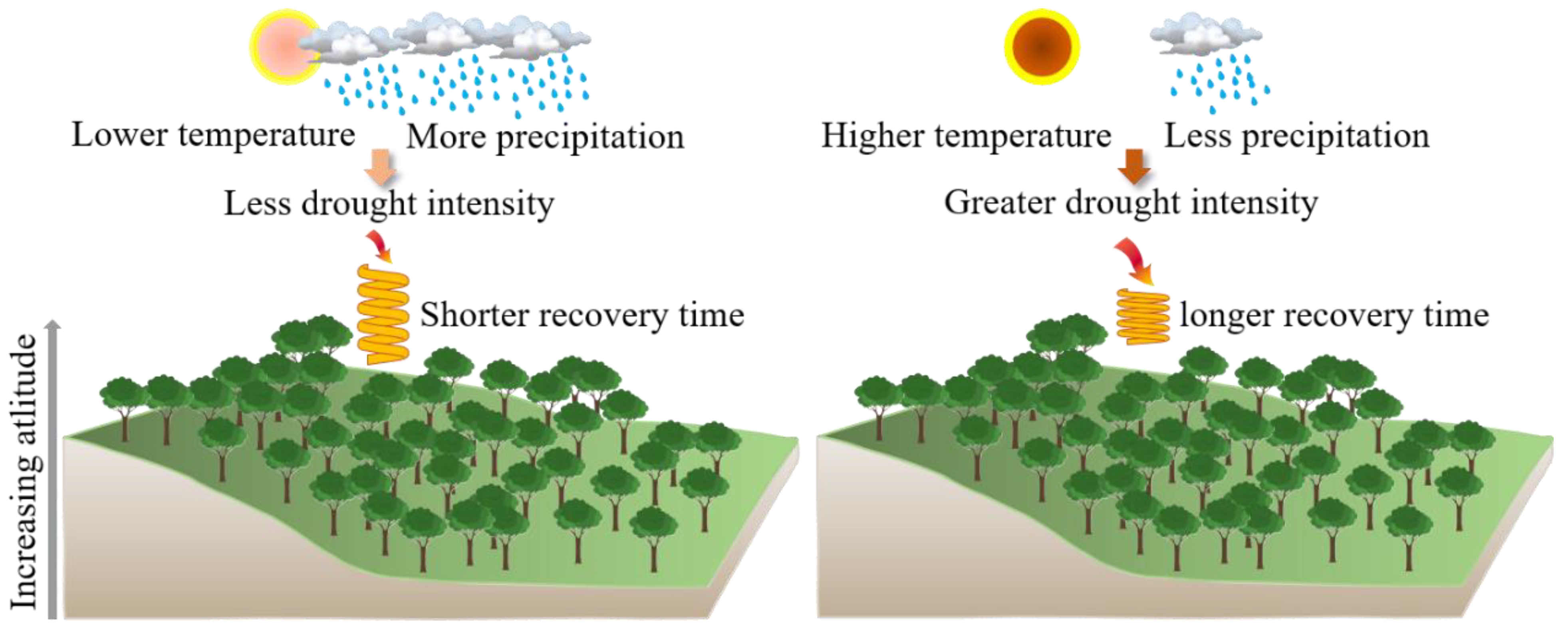
Physiological Basis of Resilience Curve
Interpretation of the Resilience Curve and Resilience Qualification
2.3.3. Resilience Curves
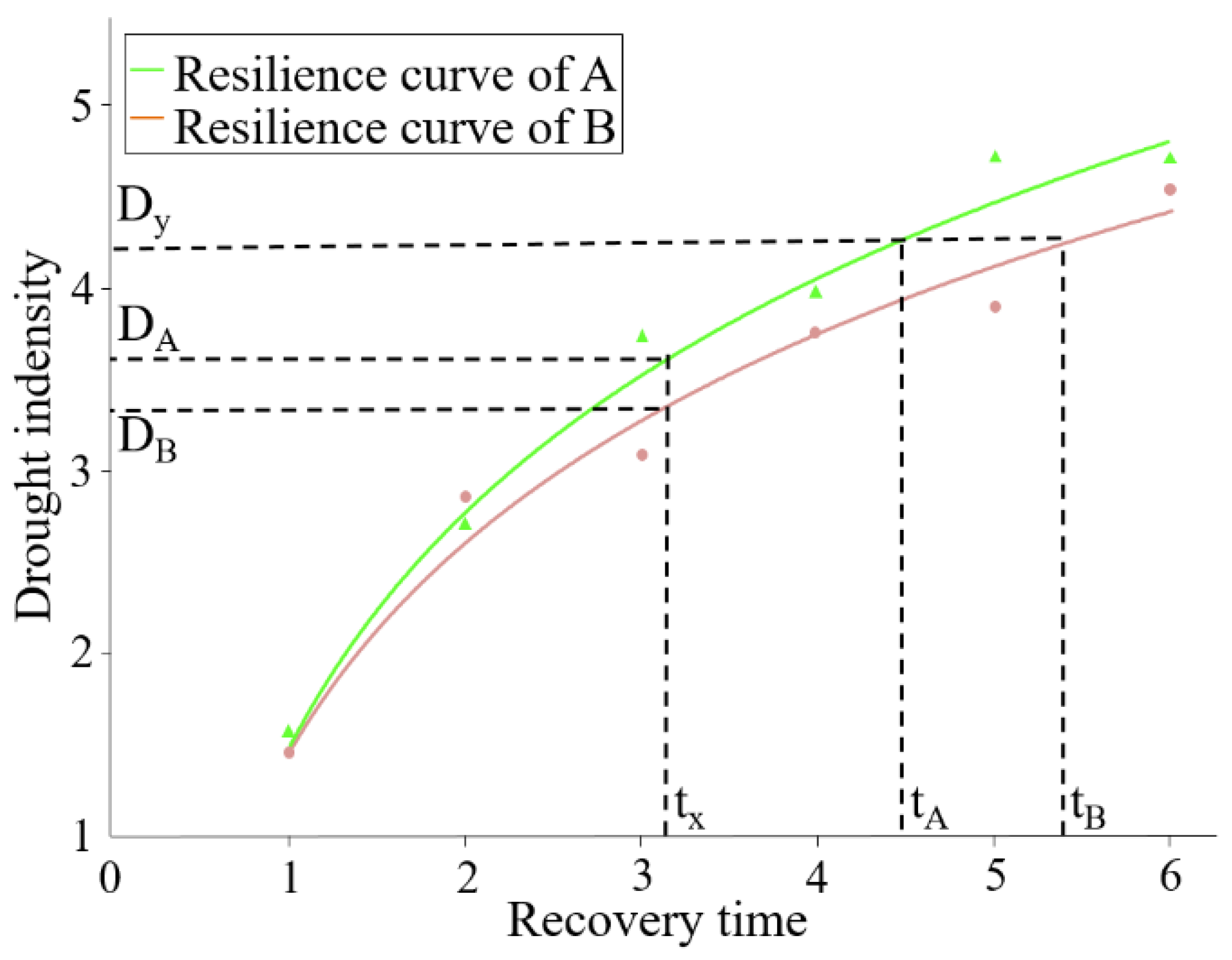
Definitions of Recovery Time and Drought Intensity
Resilience Curve and Resilience Qualification
2.3.4. Affecting Climate Factors of Forest Resilience
2.3.5. Relationships between Forest Resilience and Dominant Factors
3. Results
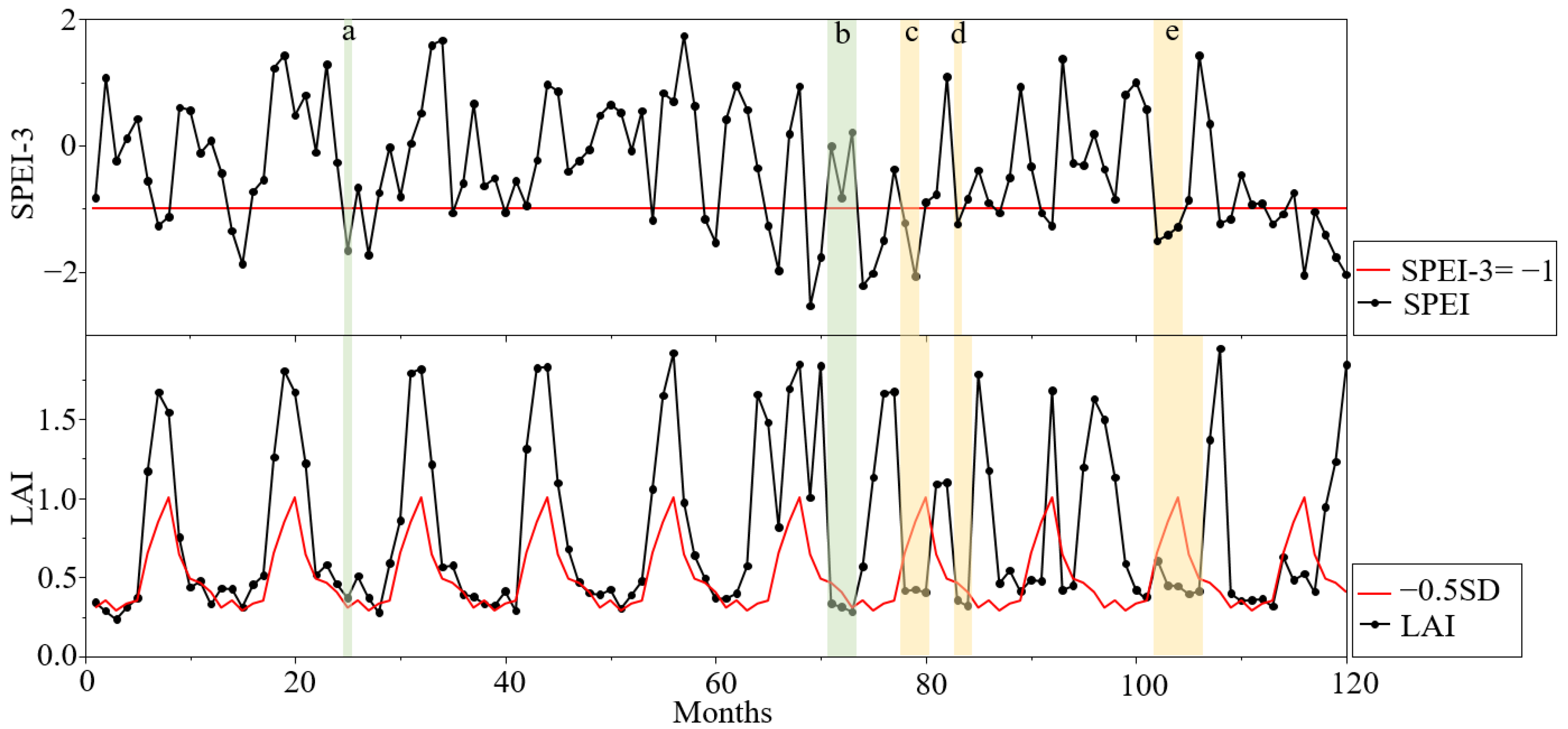
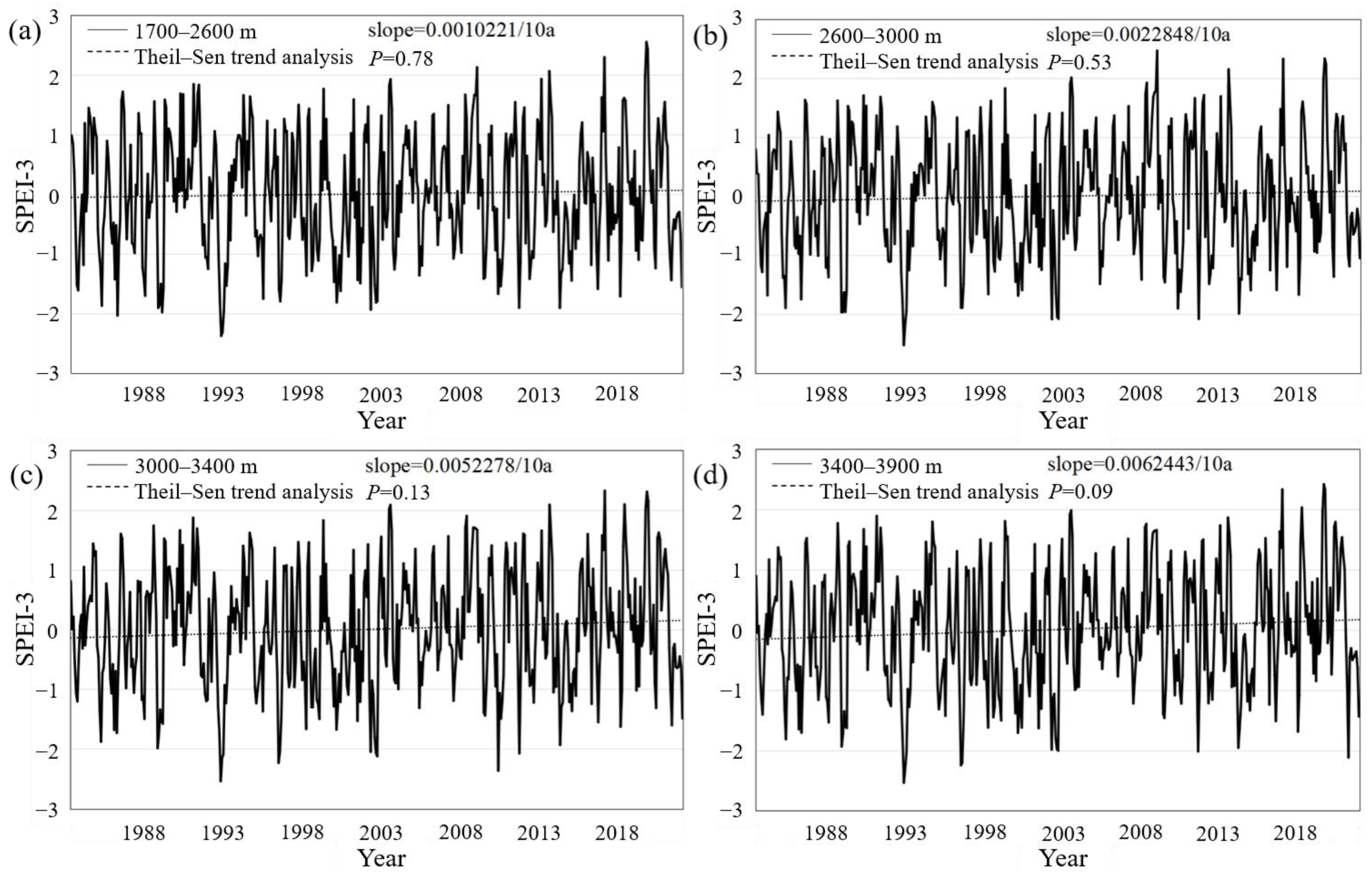
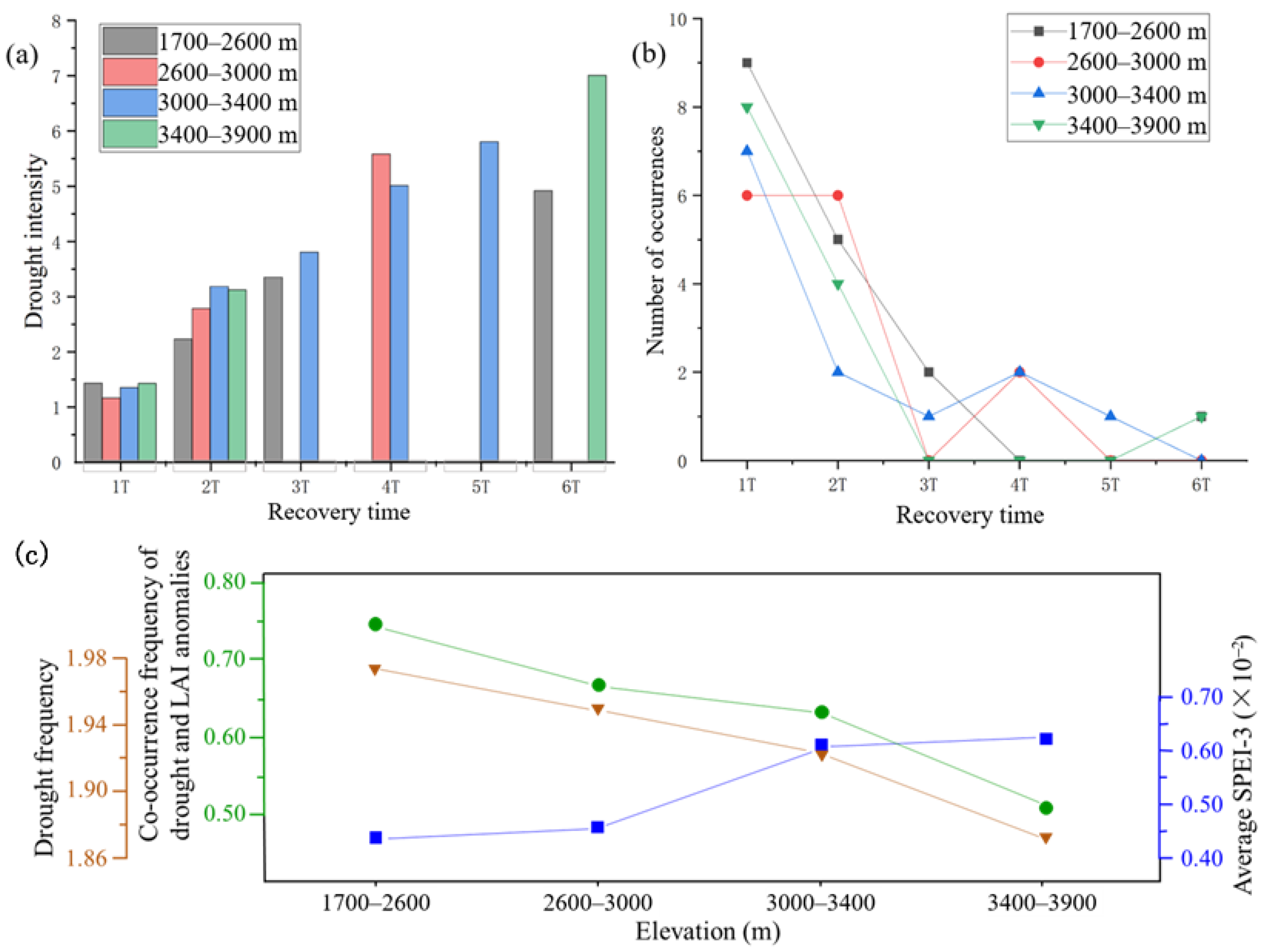
3.1. Altitudinal Differentiation of Forest Drought
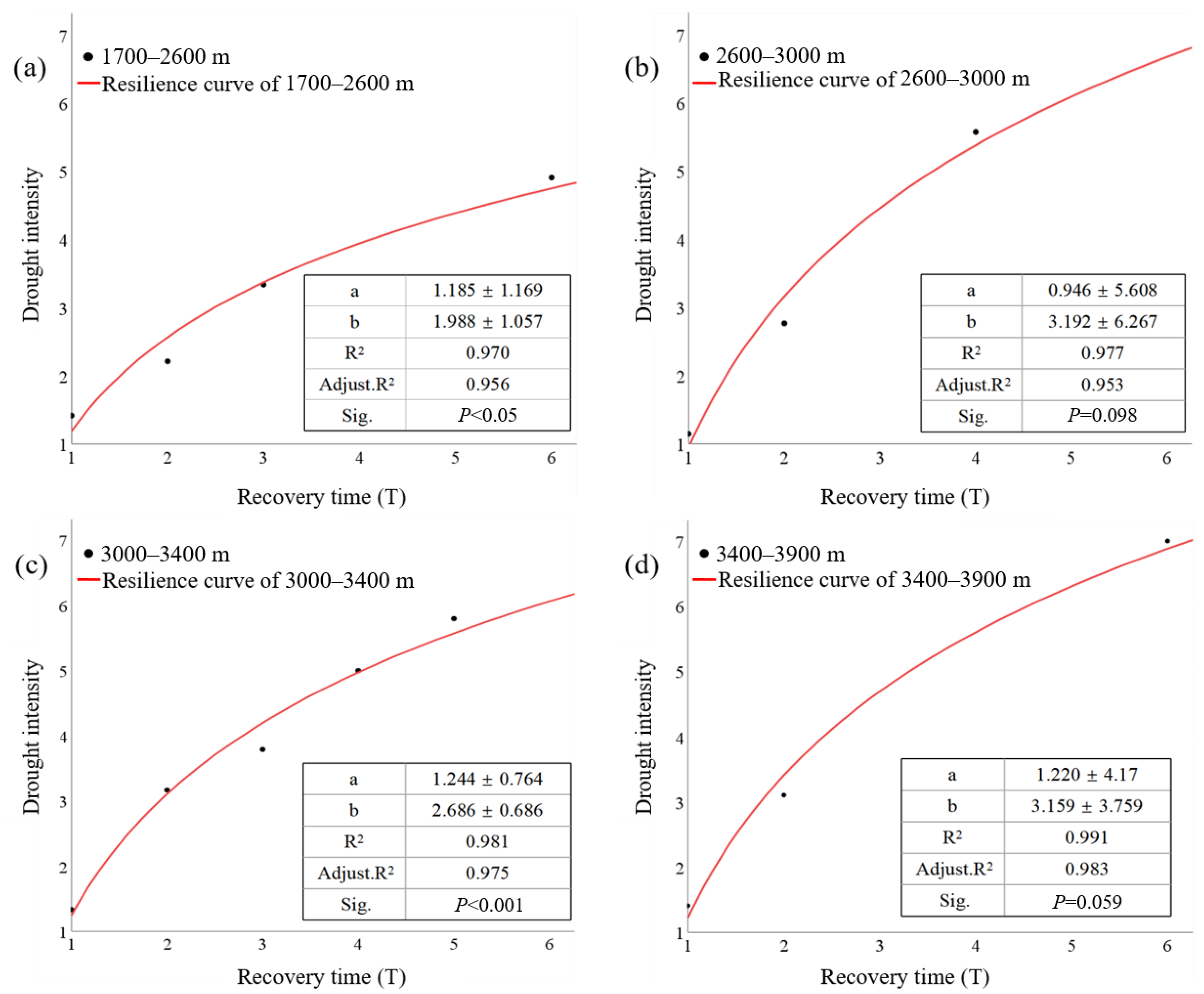
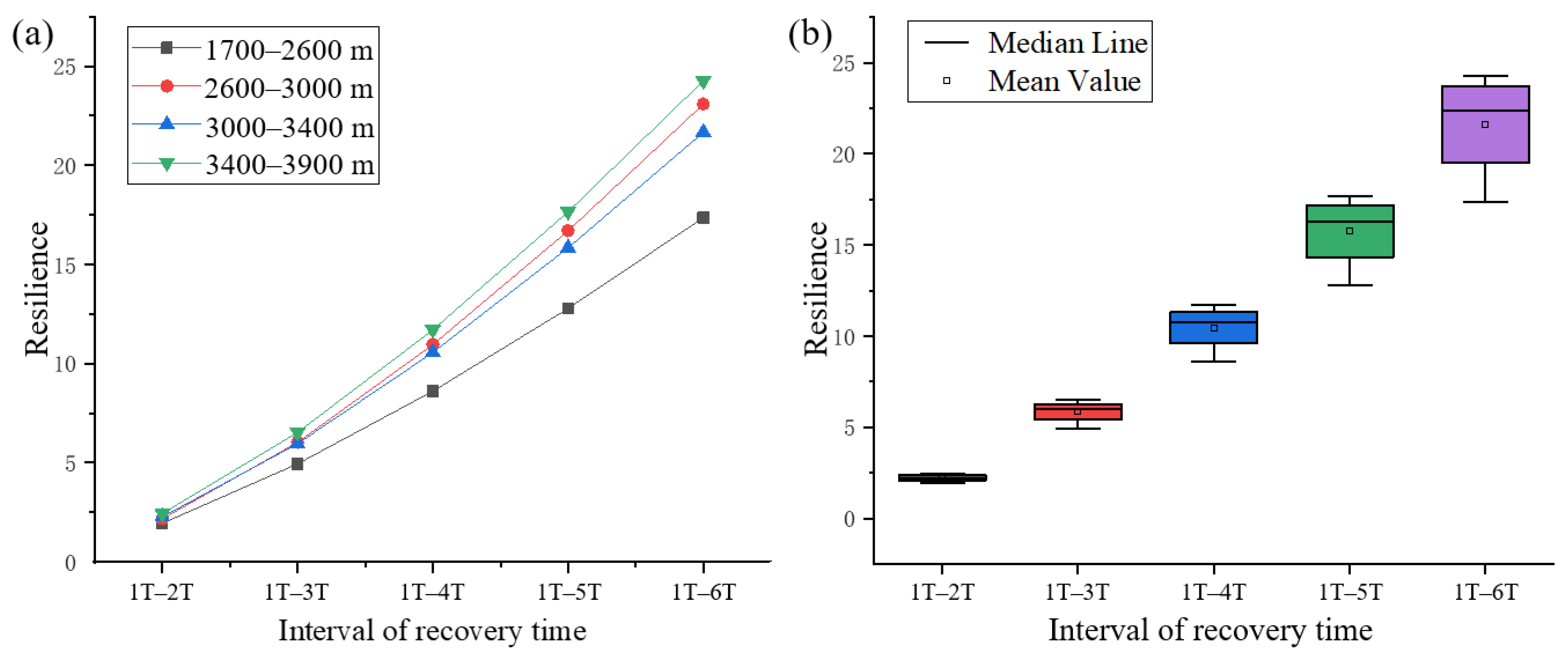
3.2. Altitudinal Differentiation of Forest Resilience to Drought
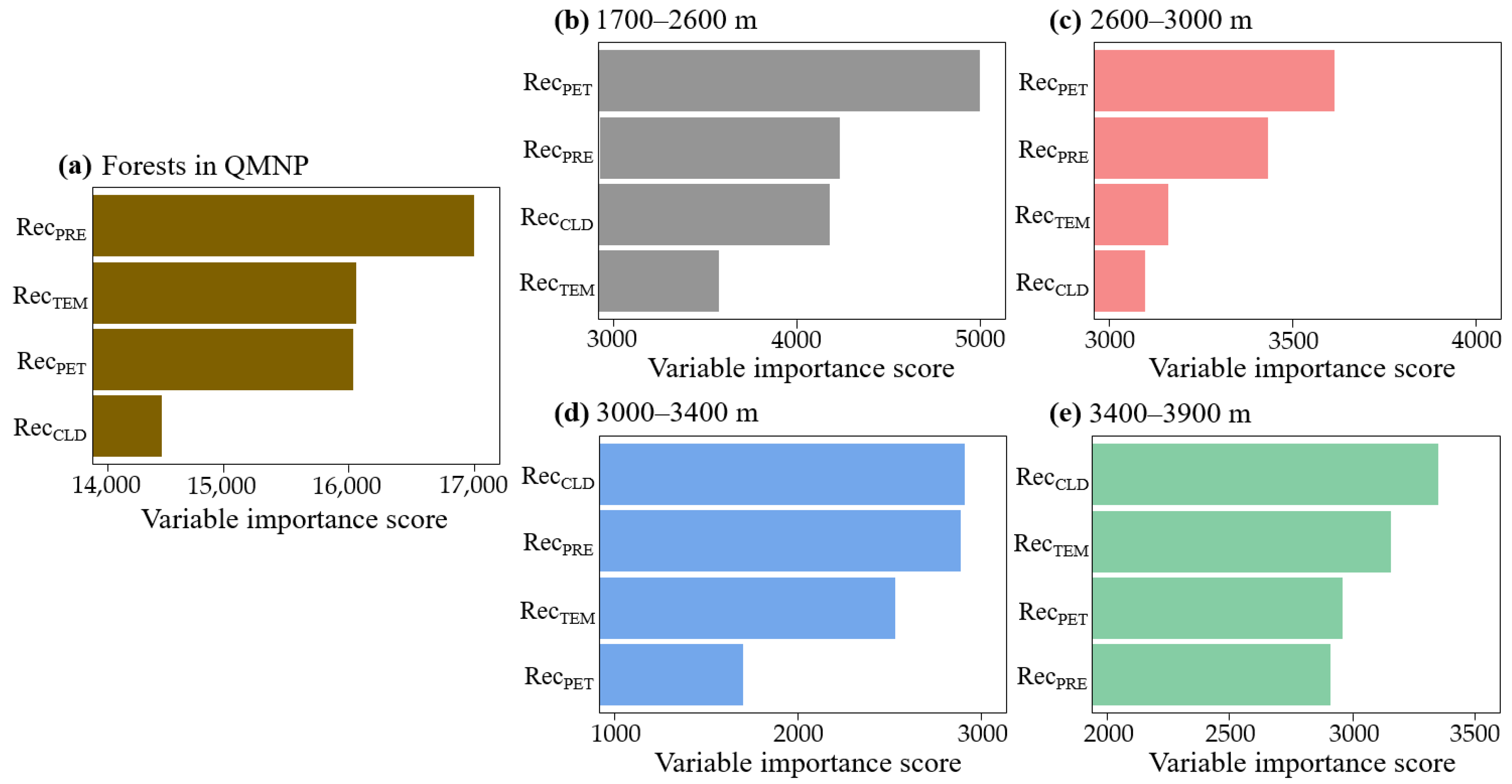
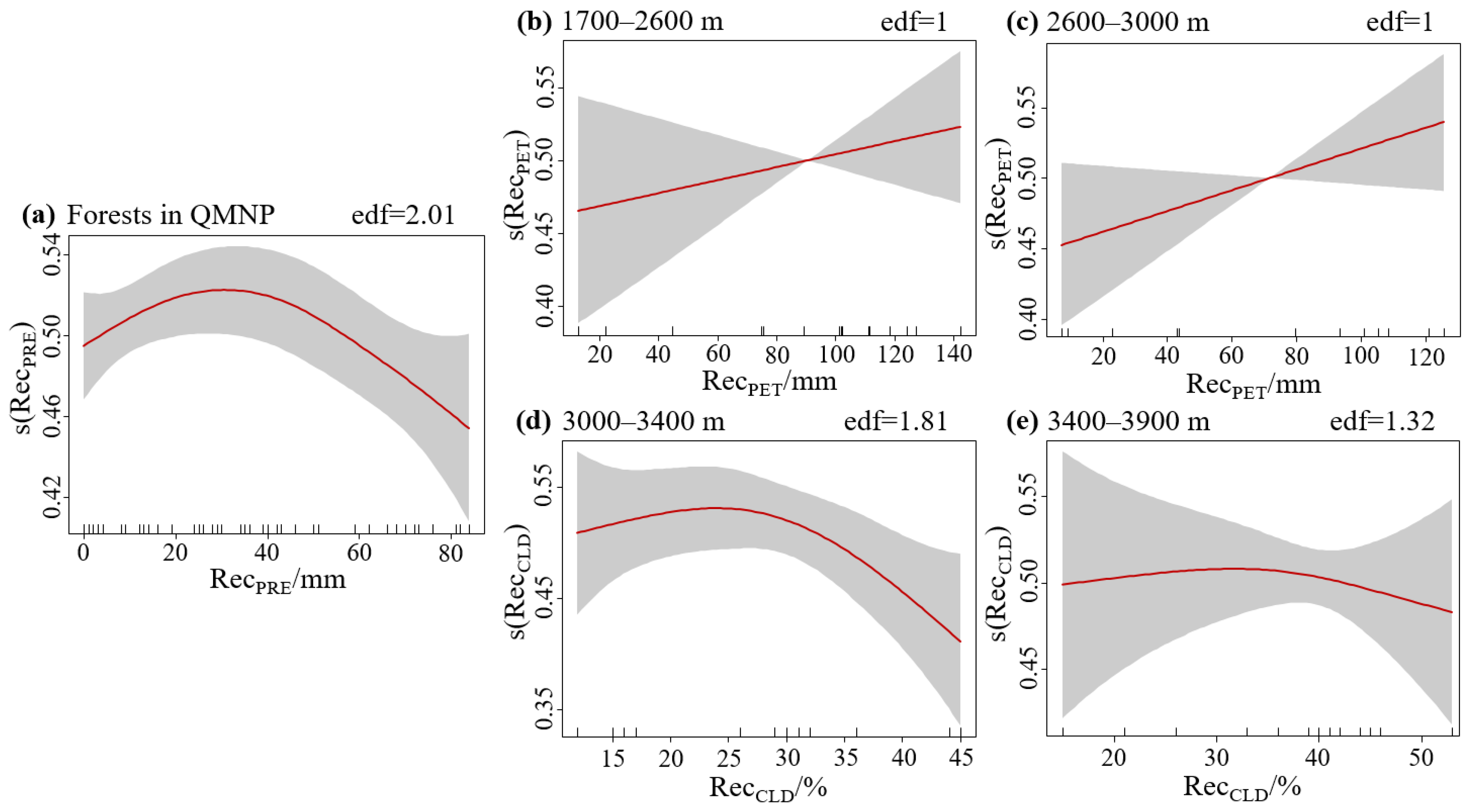
3.3. Dominant Climate Factors of Forest Resilience and Relationships
4. Discussion
4.1. Indicative Significance of the Resilience Curve for Primary Forests within the Protected Area
- (1)
- The resilience curves can help ecological departments make decisions by assessing the resilience. The resilience to drought of lower-elevation forests in the QMNP is lower than that of higher-elevation forests, and the sensitivity to drought is higher (Figure 8 and Figure 9). This indicates that the relevant departments should focus on formulating ecological environmental protection policies and strengthening the sustainable management of lower-elevation forests in the future.
- (2)
- The resilience curves allow us to estimate the recovery time required for ecosystems to recover after a drought event of certain intensity. For instance, if the value of drought intensity in QMNP is five, the recovery times for forests at elevations of 1700–2600 m, 2600–3000 m, 3000–3400 m, and 3400–3900 m are 6.8 T, 3.6 T, 4 T, and 3.3 T, respectively (Figure 8). The resilience curve provides scientific data to help primary forests cope with drought disturbance in order to prevent ecosystem collapse in the future.
- (3)
- The protected area could predict changes in resilience and tree lines based on future climate change through the resilience curves. The average annual temperature and average annual precipitation in the QMNP have been increasing in recent years, and the climate has tended to become warmer and more humid [25,26]. The average annual temperature in the QMNP increased by 0.048 °C/y and the average annual precipitation increased by 1.17 mm/y from 1985 to 2019. Because March–October is the peak season for forest growth in the Qilian Mountains, the response of forest ecosystem resilience to drought is mainly reflected in spring, summer, and autumn, where the annual average temperature increased by 0.069, 0.056, and 0.044 °C/y, and the annual average rainfall increased by 0.020, 0.545, and 0.669 mm/y, respectively [76]. Therefore, forests in the growing season in the QMNP will become warmer and wetter in the future, forest resilience to drought will become stronger, forests with strong resilience will expand, and the lower forest line will decline and the upper forest line will rise in the future.
4.2. Dominant Climate Factors Underlying Altitude Differentiation of Forest Resilience
4.3. Limitations and Perspectives
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
- (1)
- Calculation of
- (2)
- Calculation of the difference between precipitation and
- (3)
- Normalization of the data series using the log-logistic probability distribution
- (4)
- The cumulative probability distribution of the calculated sequence is normalized to yield the value
References
- Berdugo, M.; DelgadoBaquerizo, M.; Soliveres, S.; Hernández-Clemente, R.; Zhao, Y.; Gaitán, J.J.; Gross, N.; Saiz, H.; Maire, V.; Lehmann, A.; et al. Global ecosystem thresholds driven by aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Orimoloye, I.R.; Belle, J.A.; Ololade, O.O. Drought disaster monitoring using MODIS derived index for drought years: A space-based information for ecosystems and environmental conservation. J. Environ. Manag. 2021, 284, 112028. [Google Scholar] [CrossRef]
- Zhang, Y.; Keenan, T.F.; Zhou, S. Exacerbated drought impacts on global ecosystems due to structural overshoot. Nat. Ecol. Evol. 2021, 5, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.C.; Camarero, J.J.; Carreira, J.A. Competition modulates the adaptation capacity of forests to climatic stress: Insights from recent growth decline and death in relict stands of the Mediterranean fir Abies pinsapo. J. Ecol. 2010, 98, 592–603. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Ledger, M.E.; Brown, L.E.; Edwards, F.K.; Milner, A.M.; Woodward, G. Drought alters the structure and functioning of complex food webs. Nat. Clim. Chang. 2013, 3, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Hanke, H.; Borjeson, L.; Hylander, K.; Enfors-Kautsky, E. Corrigendum to “Drought tolerant species dominate as rainfall and tree cover returns in the West African Sahel”. Land Use Policy 2016, 59, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Z.; Chen, G. A sensitivity study of applying a two-source potential evapotranspiration model in the standardized precipitation evapotranspiration index for drought monitoring. Land Degrad. Dev. 2017, 28, 783–793. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Quiring, S.M.; Peña-Gallardo, M.; Yuan, S.; Domínguez-Castro, F. A review of environmental droughts: Increased risk under global warming? Earth Sci. Rev. 2020, 201, 102953. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Dakos, V.; Scheffer, M.; Van Nes, E.H.; Brovkin, V.; Petoukhov, V.; Held, H. Slowing down as an early warning signal for abrupt climate change. Proc. Natl Acad. Sci. USA 2008, 105, 14308–14312. [Google Scholar] [CrossRef] [Green Version]
- Arani, B.M.S.; Carpenter, S.R.; Lahti, L.; van Nes, E.H.; Scheffer, M. Exit time as a measure of ecological resilience. Science 2021, 372, 4895. [Google Scholar] [CrossRef]
- De Faria, B.L.; Marano, G.; Piponiot, C.; Silva, C.A.; Dantas, V.d.L.; Rattis, L.; Rech, A.R.; Collalti, A. Model-based estimation of Amazonian forests recovery time after drought and fire events. Forests 2021, 12, 8. [Google Scholar] [CrossRef]
- Schwalm, C.R.; Anderegg, W.R.L.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Wolf, A.; et al. Global patterns of drought recovery. Nature 2017, 548, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Li, D.; Hararuk, O.; Schwalm, C.R.; Luo, Y.; Yan, L.; Niu, S. Recovery time and state change of terrestrial carbon cycle after disturbance. Environ. Res. Lett. 2017, 12, 104004. [Google Scholar] [CrossRef]
- Ingrisch, J.; Bahn, M. Towards a Comparable Quantification of Resilience. Trends Ecol. Evol. 2018, 33, 251–259. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Wu, X.; Li, X.; Shi, F. Postdrought recovery time across global terrestrial ecosystems. J. Geophys. Res. Biogeosci. 2021, 126, e2020JG005699. [Google Scholar] [CrossRef]
- Yao, Y.; Fu, B.; Liu, Y.; Li, Y.; Wang, S.; Zhan, T.; Wang, Y.; Gao, D. Evaluation of ecosystem resilience to drought based on drought intensity and recovery time. Agric. For. Meteorol. 2022, 314, 108809. [Google Scholar] [CrossRef]
- Forzieri, G.; Dakos, V.; McDowell, N.G.; Ramdane, A.; Cescatti, A. Emerging signals of declining forest resilience under climate change. Nature 2022, 608, 534–539. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, J.; Wang, J. Response of vegetation cover to drought in the Qilian Mountains Region from 2001 to 2016. Pratac. Sci. 2021, 38, 419–431, (Chinese with English Abstract). [Google Scholar]
- Ding, W.; Gou, X.; Li, Y. Annual Report On Development of Qilian Ecosystem (2020); Social Sciences Academic Press (China): Beijing, China, 2020; pp. 18–43. (In Chinese) [Google Scholar]
- Li, Y.; Li, Z.; Zhang, X.; Gui, J.; Xue, J. Vegetation variations and its driving factors in the transition zone between Tibetan Plateau and arid region. Ecol. Indic. 2022, 141, 109101. [Google Scholar]
- Ma, Y.; Guan, Q.; Sun, Y.; Zhang, J.; Yang, L.; Yang, E.; Li, H.; Du, Q. Three-dimensional dynamic characteristics of vegetation and its response to climatic factors in the Qilian Mountains. CATENA 2022, 208, 105694. [Google Scholar] [CrossRef]
- Gou, X.; Hou, F.; Li, Y.; Zhao, C.; Zou, S. Scientific Investigation Report on Ecosystem Changes in Qilian Mountains; Science Press: Beijing, China, 2022; pp. 20–52. (In Chinese) [Google Scholar]
- Xu, Z.; Zhao, C.; Feng, Z. A study of the impact of climate change on the potential distribution of Qinghai spruce (Picea crassifolia) in Qilian Mountains. Acta Ecol. Sin. 2009, 29, 278–285. [Google Scholar] [CrossRef]
- Zhang, H.; Han, W.; Song, J.; Li, M. Spatial-temporal variations of habitat quality in Qilian Mountain National Park. Chin. J. Ecol. 2021, 40, 1419–1430, (Chinese with English Abstract). [Google Scholar]
- Xue, J.; Li, Z.; Feng, Q.; Miao, C.; Deng, X.; Di, Z.; Ye, A.; Gong, W.; Zhang, B.; Gui, J.; et al. Spatiotemporal variation characteristics of water conservation amount in the Qilian Mountains from 1980 to 2017. J. Giaciol. Geocryol. 2022, 44, 1–13, (Chinese with English Abstract). [Google Scholar]
- Zhou, X.; Li, Y. Response of dry-wet change to millennial and centennial warm periods in the Qilian Mountains. Acta Geogr. Sin. 2022, 77, 1138–1152, (Chinese with English Abstract). [Google Scholar]
- Harris, I.; Jones, P.; Osborn, T.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.; Hijmans, R. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Peng, S.; Ding, Y.; Liu, W.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Ding, Y.; Wen, Z.; Chen, Y.; Cao, Y.; Ren, J. Spatiotemporal change and trend analysis of potential evapotranspiration over the Loess Plateau of China during 2011–2100. Agric. For. Meteorol. 2017, 233, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Black, T.A. Measuring leaf area index of plant canopies with branch architecture. Agric. For. Meteorol. 1991, 57, 1–12. [Google Scholar] [CrossRef]
- Fang, H.; Baret, F.; Plummer, S.; Schaepman-Strub, G. An overview of global leaf area index (LAI): Methods, products, validation, and applications. Rev. Geophys. 2019, 57, 739–799. [Google Scholar] [CrossRef]
- Yan, G.; Hu, R.; Luo, J.; Weiss, M.; Jiang, H.; Mu, X.; Xie, D.; Zhang, W. Review of indirect optical measurements of leaf area index: Recent advances, challenges, and perspectives. Agric. For. Meteorol. 2019, 265, 390–411. [Google Scholar] [CrossRef]
- Wang, D.; Lv, S.; Han, B.; Meng, X.; Li, Z.; Zhang, J. The characteristics of spring vegetation cover and its response to spring drought over the Loess Plateau. Plateau Meteorol. 2018, 37, 1208–1219, (Chinese with English Abstract). [Google Scholar]
- Yang, J.; Huang, X. The 30 m annual land cover datasets and its dynamics in China from 1990 to 2020 (1.0.0) [Data set]. Zenodo 2021, 13, 3907–3925. [Google Scholar]
- Li, Y.; Ding, J.; Zhang, J.; Wu, P. Response of vegetation cover to drought in the northern slope of the Tianshan Mountains during 2001--2015 based on the land-use and land-cover change. Acta Ecol. Sin. 2019, 39, 6206–6217, (Chinese with English Abstract). [Google Scholar]
- Hou, Q.; Pei, T.; Chen, Y.; Ji, Z.; Xie, B. Variations of drought and its trend in the Loess Plateau from 1986 to 2019. Chin. J. Appl. Ecol. 2021, 32, 649–660, (Chinese with English Abstract). [Google Scholar]
- Greenwood, S.; Ruiz-Benito, P.; Martínez-Vilalta, J.; Lloret, F.; Kitzberger, T.; Allen, C.D.; Fensham, R.; Laughlin, D.C.; Kattge, J.; Bönisch, G.; et al. Tree mortality across biomes is promoted by drought intensity, lower wood density and higher specific leaf area. Ecol. Lett. 2017, 20, 539–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slette, I.J.; Post, A.K.; Awad, M.; Even, T.; Punzalan, A.; Williams, S.; Smith, M.D.; Knapp, A.K. How ecologists define drought, and why we should do better. Glob. Chang. Biol. 2019, 25, 3193–3200. [Google Scholar] [CrossRef]
- Bachmair, S.; Kohn, I.; Stahl, K. Exploring the link between drought indicators and impacts. Nat. Hazards Earth Syst. Sci. 2015, 15, 1381–1397. [Google Scholar] [CrossRef] [Green Version]
- Blauhut, V.; Stahl, K.; Stagge, J.H.; Tallaksen, L.M.; De Stefano, L.; Vogt, J. Estimating drought risk across Europe from reported drought impacts, drought indices, and vulnerability factors. Hydrol. Earth Syst. Sci. 2016, 20, 2779–2800. [Google Scholar] [CrossRef] [Green Version]
- Jiao, L.; Chen, K.; Liu, X.; Qi, C.; Xue, R. Comparison of the response stability of Siberian larch to climate change in the Altai and Tianshan. Ecol. Indic. 2021, 128, 107823. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Ren, Y.; Li, Q.; Zhou, S.; Zhao, C.; Chai, R.; Deng, P.; Wang, J. Intra-annual differences of 3-month Standardized Precipitation-Evapotranspiration Index dryness/wetness sensitivity over southwest China. Atmos. Sci. Lett. 2018, 19, e830. [Google Scholar] [CrossRef] [Green Version]
- Jiao, W.; Wang, L.; Smith, W.K.; Chang, Q.; Wang, H.; D’Odorico, P. Observed increasing water constraint on vegetation growth over the last three decades. Nat. Commun. 2021, 12, 3777. [Google Scholar] [CrossRef]
- Thornthwaite, C.W. An approach towards a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Franzke, C. Nonlinear climate change. Nat. Clim. Chang. 2014, 4, 423–424. [Google Scholar] [CrossRef]
- Ji, F.; Wu, Z.; Huang, J.; Chassignet, E.P. Evolution of land surface air temperature trend. Nat. Clim. Chang. 2014, 4, 462–466. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Anderegg, W.R.L.; Vicente-Serrano, S.M. Impacts of droughts on the growth resilience of northern hemisphere forests: Forest growth resilience to drought. Glob. Ecol. Biogeogr. 2017, 26, 166–176. [Google Scholar] [CrossRef]
- Reyniers, N.; Osborn, T.J.; Addor, N.; Darch, G. Projected changes in droughts and extreme droughts in Great Britain strongly influenced by the choice of drought index. Hydrol. Earth Syst. Sci. 2023, 27, 1151–1171. [Google Scholar] [CrossRef]
- Gampe, D.; Zscheischler, J.; Reichstein, M.; O’Sullivan, M.; Smith, W.K.; Sitch, S.; Buermann, W. Increasing impact of warm droughts on northern ecosystem productivity over recent decades. Nat. Clim. Chang. 2021, 11, 772–779. [Google Scholar] [CrossRef]
- Rhee, J.; Im, J. Meteorological drought forecasting for ungauged areas based on machine learning: Using long-range climate forecast and remote sensing data. Agric. For. Meteorol. 2017, 237–238, 105–122. [Google Scholar] [CrossRef]
- Yao, Y.; Liu, Y.; Zhou, S.; Song, J.; Fu, B. Soil moisture determines the recovery time of ecosystems from drought. Glob. Chang. Biol. 2023, 29, 3562–3574. [Google Scholar] [CrossRef]
- Liu, Y.; Kumar, M.; Katul, G.G.; Porporato, A. Reduced resilience as an early warning signal of forest mortality. Nat. Clim. Chang. 2019, 9, 880–885. [Google Scholar] [CrossRef]
- Zhai, J.; Mondal, S.K.; Fischer, T.; Wang, Y.; Su, B.; Huang, B.; Tao, H.; Wang, G.; Ullah, W.; Uddin, M.J. Future drought characteristics through a multi-model ensemble from CMIP6 over South Asia. Atmos. Res. 2020, 246, 105111. [Google Scholar] [CrossRef]
- Chiang, F.; Mazdiyasni, O.; AghaKouchak, A. Evidence of anthropogenic impacts on global drought frequency, duration, and intensity. Nat. Commun. 2021, 12, 2754. [Google Scholar] [CrossRef]
- Jones, S.R. Areas, anti-derivatives, and adding up pieces: Definite integrals in pure mathematics and applied science contexts. J. Math. Behav. 2015, 38, 9–28. [Google Scholar] [CrossRef]
- Moore, D.; Bach, V.; Finkbeiner, M.; Honkomp, T.; Ahn, H.; Sprenger, M.; Froese, L.; Gratzel, D. Offsetting environmental impacts beyond climate change: The Circular Ecosystem Compensation approach. J. Environ. Manag. 2023, 329, 117068. [Google Scholar] [CrossRef]
- Dang, D.; Li, X.; Li, S.; Lyu, X.; Dou, H.; Li, M.; Wang, K. Changed ecosystem stability to climate anomalies in the context of ecological restoration projects. Land. Degrad. Dev. 2023, 34, 3003–3016. [Google Scholar] [CrossRef]
- Higgins, S.I.; Conradi, T.; Muhoko, E. Shifts in vegetation activity of terrestrial ecosystems attributable to climate trends. Nat. Geosci. 2023, 16, 147–153. [Google Scholar] [CrossRef]
- Wu, J.; Liang, S. Assessing Terrestrial Ecosystem Resilience using Satellite Leaf Area Index. Remote Sens. 2020, 12, 595. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- Isabona, J.; Imoize, A.L.; Kim, Y. Machine learning-based boosted regression ensemble combined with hyperparameter tuning for optimal adaptive learning. Remote Sens. 2022, 22, 3776. [Google Scholar] [CrossRef] [PubMed]
- Hastie, T.; Tibshirani, R. Generalized Additive Models. Stat. Sci. 1986, 3, 297–318. [Google Scholar] [CrossRef]
- Guisan, A.; Edwards, T.C.; Hastie, T. Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Xu, L.; Bjørnstad, O.N.; Liu, K.; Song, T.; Chen, A.; Xu, B.; Liu, Q.; Stenseth, N.C. Climate-driven variation in mosquito density predicts the spatiotemporal dynamics of dengue. Proc. Natl. Acad. Sci. USA 2019, 116, 3624–3629. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, Y.; Shao, M.; Jia, X.; Li, X. Spatiotemporal analysis of multiscalar drought characteristics across the Loess Plateau of China. J. Hydrol. 2016, 534, 281–299. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, Q.; Zhao, X.; Wu, P.; Pan, W.; Gao, X.; Sun, M. Temporal and spatial evolution of the standardized precipitation evapotranspiration index (SPEI) in the Loess Plateau under climate change from 2001 to 2050. Sci. Total Environ. 2017, 595, 191–200. [Google Scholar] [CrossRef]
- Dong, Z.; Liu, H.; Baiyinbaoligao; Hu, H.; Khan, M.Y.A.; Wen, J.; Chen, L.; Tian, F. Future projection of seasonal drought characteristics using CMIP6 in the Lancang-Mekong River Basin. J. Hydrol. 2022, 610, 127815. [Google Scholar] [CrossRef]
- Xia, S. A discussion of discrete cofficient. Coll. Math. 1997, 2, 144–146, (Chinese with English Abstract). [Google Scholar]
- Gao, X.; Huang, X.; Lo, K.; Dang, Q.; Wen, R. Vegetation responses to climate change in the Qilian Mountain Nature Reserve, Northwest China. Glob. Ecol. Conserv. 2021, 28, e01698. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, C.; Wang, X. Elevational differences in the net primary productivity response to climate constraints in a dryland mountain ecosystem of northwestern China. Land. Degrad. Dev. 2020, 31, 2087–2103. [Google Scholar] [CrossRef]
- Du, Q.; Sun, Y.; Guan, Q.; Pan, N.; Wang, Q.; Ma, Y.; Li, H.; Liang, L. Vulnerability of grassland ecosystems to climate change in the Qilian Mountains, northwest China. J. Hydrol. 2022, 612, 128305. [Google Scholar] [CrossRef]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 2018, 556, 235–238. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Bohner, T.; Diez, J. Tree resistance and recovery from drought mediated by multiple abiotic and biotic processes across a large geographic gradientzone. Sci. Total Environ. 2021, 789, 147744. [Google Scholar] [CrossRef]
- Marcotti, E.; Amoroso, M.M.; Rodríguez-Caton, M.; Vega, L.; Srur, A.M.; Villalba, R. Growth resilience of Austrocedrus chilensis to drought along a precipitation gradientzone in Patagonia. Argentina. For. Ecol. Manag. 2021, 496, 119388. [Google Scholar] [CrossRef]
- Jing, M.; Che, Z.; Jing, W.; Zhao, W.; Ma, Y.; Liu, S. Spatiotemporal Changes of Precipitation and Temperature at Water Resources Conservation Forests Area in Oilian Mountains, China. J. Desert Res. 2012, 32, 1071–1076, (Chinese with English Abstract). [Google Scholar]
- Zhang, X.; Zhang, Y.; Tian, J.; Ma, N.; Wang, Y. CO2 fertilization is spatially distinct from stomatal conductance reduction in controlling ecosystem water-use efficiency increase. Environ. Res. Lett. 2022, 17, 054048. [Google Scholar] [CrossRef]
- Coban, O.; De Deyn, G.B. Soil microbiota as game-changers in restoration of degraded lands. Science 2022, 375, 6584. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Li, Y.; Teuling, A.J.; Zhao, L.; Spracklen, D.V.; Garcia-Carreras, L.; Meier, R.; Chen, L.; Zheng, Y.; Lin, H.; et al. Contrasting impacts of forests on cloud cover based on satellite observations. Nat. Commun. 2022, 13, 670. [Google Scholar] [CrossRef]
- Ji, D.; Li, Q.; Guo, Y.; An, W.; Manavski, N.; Meurer, J.; Chi, W. NADP+ supply adjusts the synthesis of photosystem I in Arabidopsis chloroplasts. Plant. Physiol. 2022, 189, 2128–2143. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carreras, L.; Parker, D.J.; Taylor, C.M.; Reeves, C.E.; Murphy, J.G. Impact of mesoscale vegetation heterogeneities on the dynamical and thermodynamic properties of the planetary boundary layer. J. Geophys. Res. Atmos. 2010, 115, D03102. [Google Scholar] [CrossRef] [Green Version]
- Zlobin, I.E. Linking the growth patterns of coniferous species with their performance under climate aridization. Sci. Total Environ. 2022, 831, 154971. [Google Scholar] [CrossRef]
- Lloret, F.; Jaime, L.A.; Margalef-Marrase, J.; Pérez-Navarro, M.A.; Batllori, E. Short-term forest resilience after drought-induced die-off in Southwestern European forests. Sci. Total Environ. 2022, 806, 150940. [Google Scholar] [CrossRef]
- Ciceu, A.; Popa, I.; Leca, S.; Pitar, D.; Chivulescu, S.; Badea, O. Climate change effects on tree growth from Romanian forest monitoring Level II plots. Sci. Total Environ. 2020, 698, 134129. [Google Scholar] [CrossRef]
- Du, M.; Zhang, F.; Gou, X.; Liu, L.; Xia, J.; Wu, X. Different responses of radial growth of Pica crassifolia to climate warming in the middle and eastern Qilian Mountains. J. Giaciol. Geocryol. 2022, 44, 14–23, (Chinese with English Abstract). [Google Scholar]
- Huang, T.; Hao, J.; Du, Y.; Tian, C.; Zhang, J.; Wang, H.; Hou, L. Regeneration characteristics of three natural Junipers forests in the Three-River Headwater Region of Qinghai Province, China. Chin. J. Appl. Ecol. 2022, 33, 297–303, (Chinese with English Abstract). [Google Scholar]
- Zhang, F.; Gou, X.; Su, J.; Gao, L.; Liu, W.; Man, Z. Age-Dependent Responses of Tree Radial Growth of Pinus tabulaeformis to Climate in Eastern Section of the Qilian Mountains. J. Giaciol. Geocryol. 2011, 33, 634–639, (Chinese with English Abstract). [Google Scholar]
- Zhao, Z.; Wang, S.; Luo, Y.; Liu, R.; Han, F.; Ma, Y. Physiological responses of a young Gansu elm (Ulmus glaucescens) to soil water loss and rehydration. Arid Zone Res. 2022, 39, 1534–1542, (Chinese with English Abstract). [Google Scholar]
- Li, X.; Piao, S.; Wang, K.; Wang, X.; Wang, T.; Ciais, P.; Chen, A.; Lian, X.; Peng, S.; Peñuelas, J. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought. Nat. Ecol. Evol. 2020, 4, 1075–1083. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, B.; Liu, Y.; Li, Y.; Feng, X.; Wang, S. Response of vegetation to drought in the Tibetan Plateau: Elevation differentiation and the dominant factors. Agric. For. Meteorol. 2021, 306, 108468. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | 1 T–2 T | 1 T–3 T | 1 T–4 T | 1 T–5 T | 1 T–6 T |
|---|---|---|---|---|---|
| 1700–2600 m | 1.9530 | 4.9461 | 8.6148 | 12.7858 | 17.3571 |
| 2600–3000 m | 2.1791 | 6.0283 | 10.9622 | 16.7026 | 23.0858 |
| 3000–3400 m | 2.2816 | 5.9686 | 10.5683 | 15.8468 | 21.6660 |
| 3400–3900 m | 2.4403 | 6.5335 | 11.7002 | 17.6651 | 24.2660 |
| Dispersion coefficient | 0.0923 | 0.1134 | 0.1259 | 0.1340 | 0.1398 |
| Dominant Factor | Standard Error | F-Value | p-Value | Deviance Explained/% | |
|---|---|---|---|---|---|
| Forests in QMNP | RecPRE | 0.03526 | 2.525 | 0.0934 | 11.7 |
| 1700–2600 m | RecPET | 0.07525 | 0.773 | 0.395 | 5.61 |
| 2600–3000 m | RecPET | 0.07649 | 2.665 | 0.134 | 21 |
| 3000–3400 m | RecCLD | 0.0732 | 2.457 | 0.132 | 38.7 |
| 3400–3900 m | RecCLD | 0.06676 | 0.116 | 0.821 | 7.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Gao, X.; Yan, A.; Chang, S.; Li, Q. Altitudinal Differentiation of Forest Resilience to Drought in a Dryland Mountain. Forests 2023, 14, 1284. https://doi.org/10.3390/f14071284
Li J, Gao X, Yan A, Chang S, Li Q. Altitudinal Differentiation of Forest Resilience to Drought in a Dryland Mountain. Forests. 2023; 14(7):1284. https://doi.org/10.3390/f14071284
Chicago/Turabian StyleLi, Jie, Xiang Gao, An Yan, Shuhang Chang, and Qiuran Li. 2023. "Altitudinal Differentiation of Forest Resilience to Drought in a Dryland Mountain" Forests 14, no. 7: 1284. https://doi.org/10.3390/f14071284