
Microevolutionary Processes in a Foundation Tree Inform Macrosystem Patterns of Community Biodiversity and Structure
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species & Collection Information
2.2. Tree Genotyping
2.3. Community Data Collection
2.4. Quantifying Fungal Endophyte Community Composition and Abundance using rDNA
2.5. Population Genetic and Phylogeographic Structure of P. fremontii
2.6. Environmental Data
2.7. Identifying Drivers of Community Similarity
3. Results
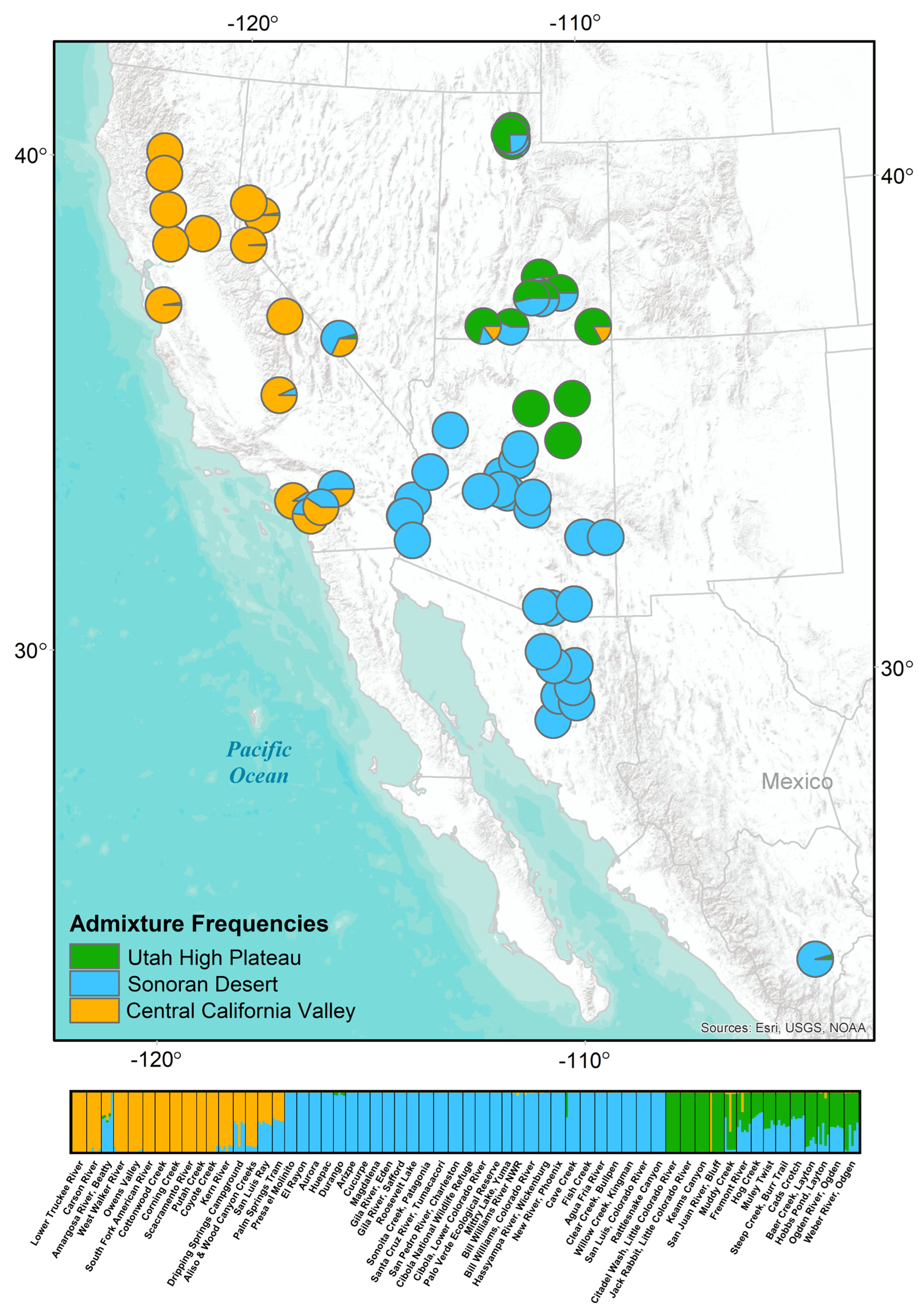
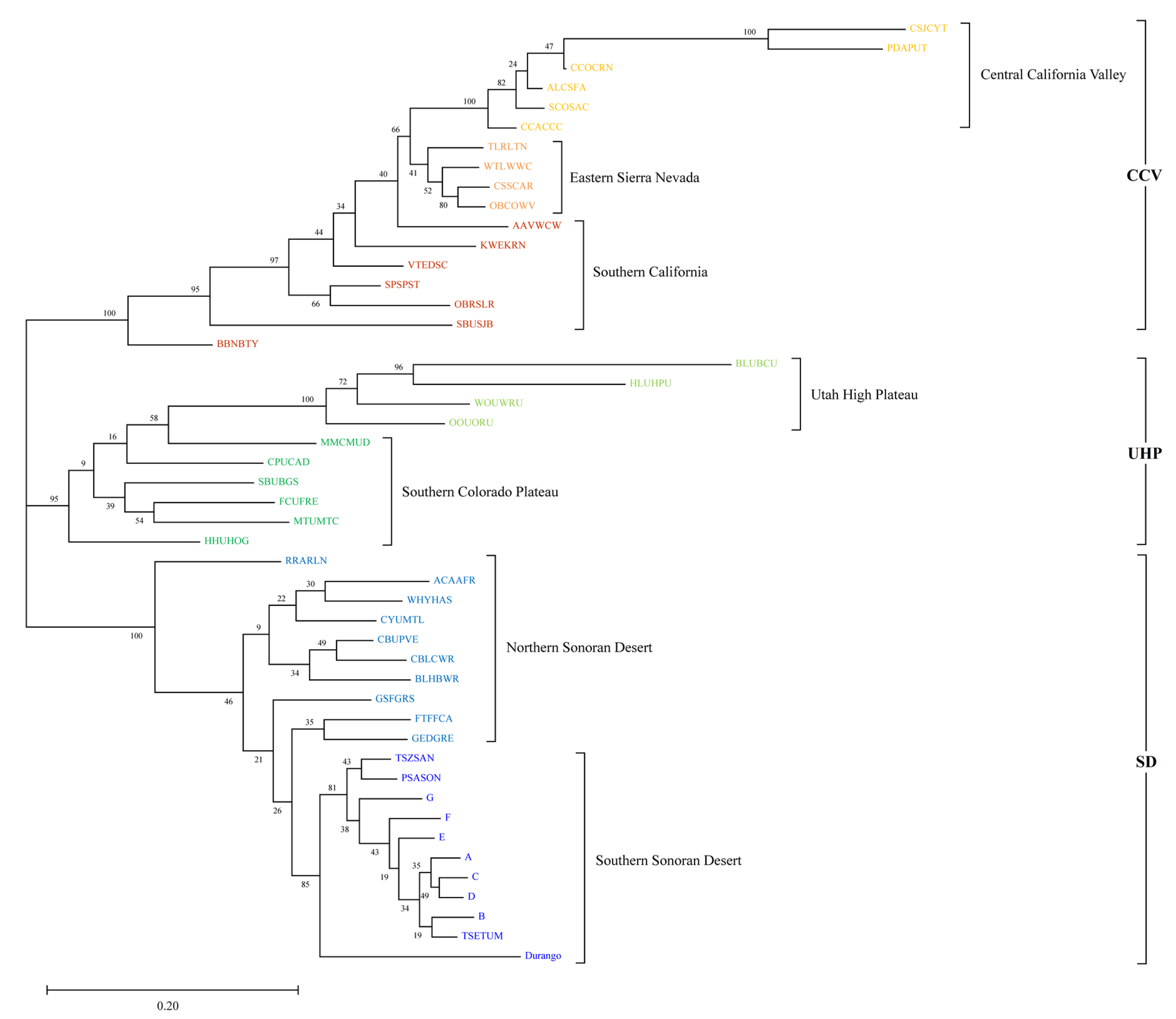
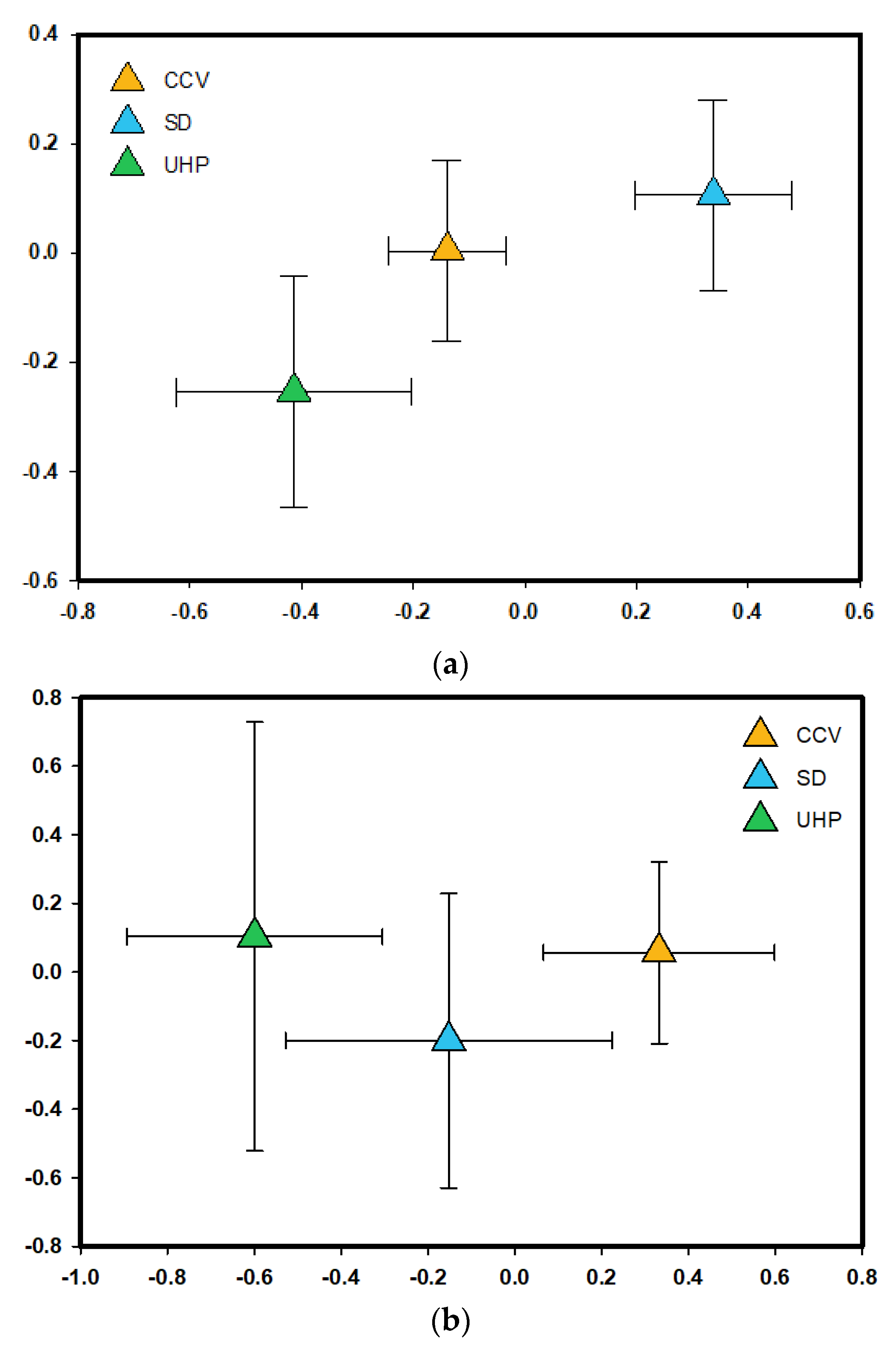
3.1. Population Genetic and Phylogeographic Structure of P. fremontii
3.2. Identifying Drivers of Community Similarity
4. Discussion
4.1. Climate as a Driver of Tree Local Adaptation
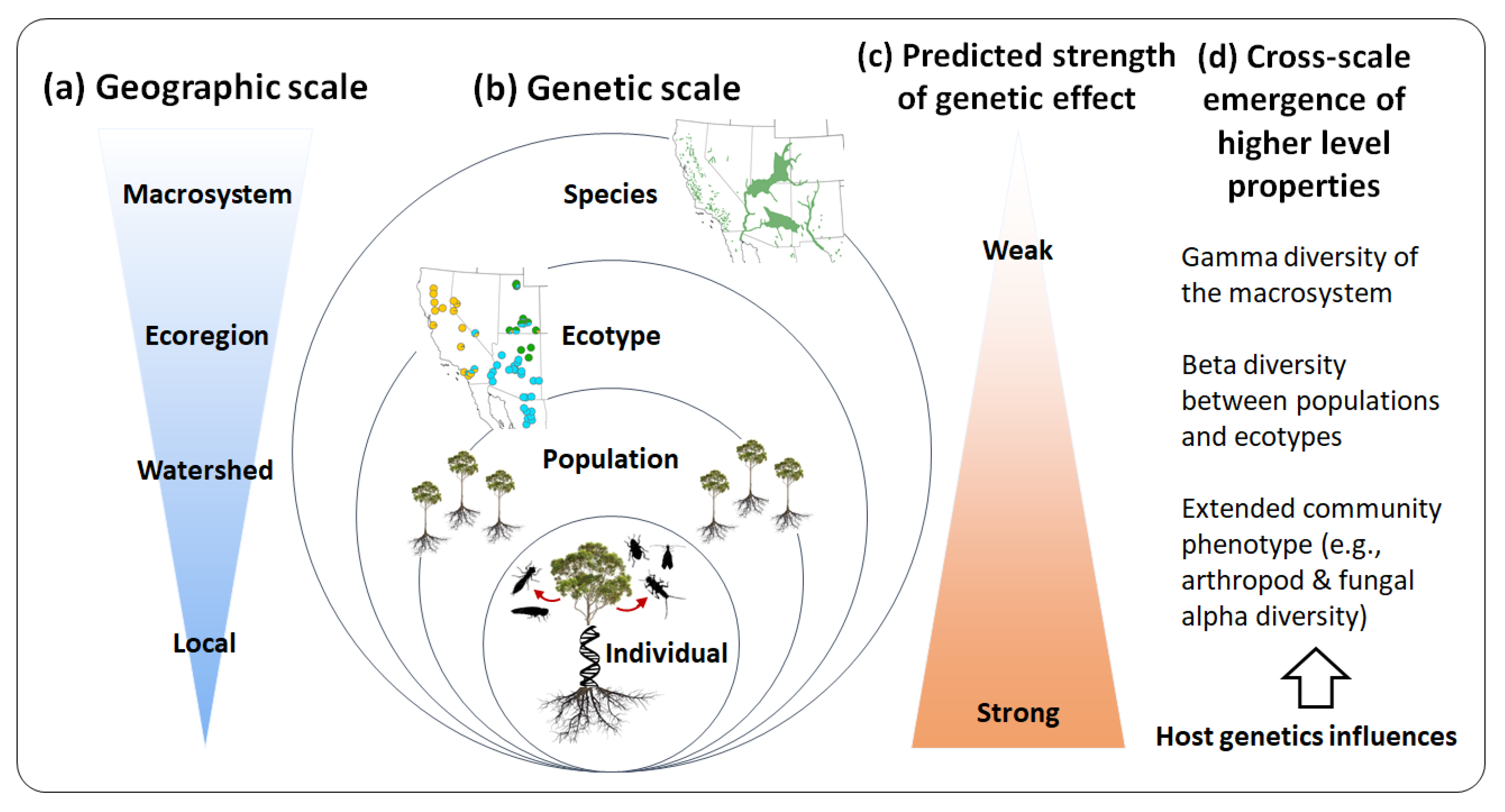
4.2. Predicting Macrosystem Properties
4.3. Conservation Management Implications
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Díaz, S.; Settele, J.; Brondízio, E.S.; Ngo, H.T.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; Chan, K.M.A.; et al. Pervasive Human-Driven Decline of Life on Earth Points to the Need for Transformative Change. Science 2019, 366, eaax3100. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Heffernan, J.B.; Soranno, P.A.; Angilletta Jr, M.J.; Buckley, L.B.; Gruner, D.S.; Keitt, T.H.; Kellner, J.R.; Kominoski, J.S.; Rocha, A.V.; Xiao, J.; et al. Macrosystems Ecology: Understanding Ecological Patterns and Processes at Continental Scales. Front. Ecol. Environ. 2014, 12, 5–14. [Google Scholar] [CrossRef]
- Peters, D.P.; Bestelmeyer, B.T.; Turner, M.G. Cross–Scale Interactions and Changing Pattern–Process Relationships: Consequences for System Dynamics. Ecosystems 2007, 10, 790–796. [Google Scholar] [CrossRef]
- Peters, D.P.; Groffman, P.M.; Nadelhoffer, K.J.; Grimm, N.B.; Collins, S.L.; Michener, W.K.; Huston, M.A. Living in an Increasingly Connected World: A Framework for Continental-Scale Environmental Science. Front. Ecol. Environ. 2008, 6, 229–237. [Google Scholar] [CrossRef]
- Peters, D.P.C.; Bestelmeyer, B.T.; Knapp, A.K. Perspectives on Global Change Theory. In The Theory of Ecology; University of Chicago Press: Chicago, IL, USA, 2011; pp. 261–281. [Google Scholar]
- Carpenter, S.R.; DeFries, R.; Dietz, T.; Mooney, H.A.; Polasky, S.; Reid, W.V.; Scholes, R.J. Millennium Ecosystem Assessment: Research Needs. Science 2006, 313, 495–497. [Google Scholar] [CrossRef]
- Macdonald, D.W.; Chiaverini, L.; Bothwell, H.M.; Kaszta, Ż.; Ash, E.; Bolongon, G.; Can, Ö.E.; Campos-Arceiz, A.; Channa, P.; Clements, G.R.; et al. Predicting Biodiversity Richness in Rapidly Changing Landscapes: Climate, Low Human Pressure or Protection as Salvation? Biodivers. Conserv. 2020, 29, 4035–4057. [Google Scholar] [CrossRef]
- Fei, S.; Guo, Q.; Potter, K. Macrosystems Ecology: Novel Methods and New Understanding of Multi-Scale Patterns and Processes. Landsc. Ecol. 2016, 31, 1–6. [Google Scholar] [CrossRef]
- Dodds, W.K.; Rose, K.C.; Fei, S.; Chandra, S. Macrosystems Revisited: Challenges and Successes in a New Subdiscipline of Ecology. Front. Ecol. Environ. 2021, 19, 4–10. [Google Scholar] [CrossRef]
- McGill, B.J.; Chase, J.M.; Hortal, J.; Overcast, I.; Rominger, A.J.; Rosindell, J.; Borges, P.A.V.; Emerson, B.C.; Etienne, R.S.; Hickerson, M.J.; et al. Unifying Macroecology and Macroevolution to Answer Fundamental Questions about Biodiversity. Glob. Ecol. Biogeogr. 2019, 28, 1925–1936. [Google Scholar] [CrossRef]
- Araújo, M.B.; Peterson, A.T. Uses and Misuses of Bioclimatic Envelope Modeling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Woodward, F.I.; Woodward, F.I. Climate and Plant Distributions; Cambridge University Press: Cambridge, UK, 1987. [Google Scholar]
- Michalet, R.; Choler, P.; Callaway, R.M.; Whitham, T.G. Rainfall Continentality, via the Winter Gams Angle, Provides a New Dimension to Biogeographical Distributions in the Western United States. Glob. Ecol. Biogeogr. 2021, 30, 384–397. [Google Scholar] [CrossRef]
- Mitton, J.B. The Union of Ecology and Evolution: Extended Phenotypes and Community Genetics. BioScience 2003, 53, 208. [Google Scholar] [CrossRef]
- Agrawal, A.A. Community Genetics: New Insights into Community Ecology by Integrating Population Genetics1. Ecology 2003, 84, 543–544. [Google Scholar] [CrossRef]
- Shuster, S.M.; Lonsdorf, E.V.; Wimp, G.M.; Bailey, J.K.; Whitham, T.G. Community Heritability Measures the Evolutionary Consequences of Indirect Genetic Effects on Community Structure. Evolution 2006, 60, 991–1003. [Google Scholar]
- Allan, G.; Shuster, S.; Woolbright, S.; Walker, F.; Meneses, N.; Keith, K.; Bailey, J.; Bangert, R.; Whitham, T. Perspective: Interspecific Indirect Genetic Effects (IIGEs). Linking Genetics and Genomics to Community Ecology and Ecosystem Processes. In Trait-Mediated Indirect Interactions: Ecological and Evolutionary Perspectives; Cambridge University Press: New York, NY, USA, 2013; pp. 295–323. [Google Scholar]
- Whitham, T.G.; Bailey, J.K.; Schweitzer, J.A.; Shuster, S.M.; Bangert, R.K.; LeRoy, C.J.; Lonsdorf, E.V.; Allan, G.J.; DiFazio, S.P.; Potts, B.M.; et al. A Framework for Community and Ecosystem Genetics: From Genes to Ecosystems. Nat. Rev. Genet. 2006, 7, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Whitham, T.G.; Allan, G.J.; Cooper, H.F.; Shuster, S.M. Intraspecific Genetic Variation and Species Interactions Contribute to Community Evolution. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 587–612. [Google Scholar] [CrossRef]
- Keith, A.R.; Bailey, J.K.; Whitham, T.G. A Genetic Basis to Community Repeatability and Stability. Ecology 2010, 91, 3398–3406. [Google Scholar] [CrossRef] [PubMed]
- Ferrier, S.M.; Bangert, R.K.; Hersch-Green, E.I.; Bailey, J.K.; Allan, G.J.; Whitham, T.G. Unique Arthropod Communities on Different Host-Plant Genotypes Results in Greater Arthropod Diversity. Arthropod-Plant Interact. 2012, 6, 187–195. [Google Scholar] [CrossRef]
- Lamit, L.J.; Busby, P.E.; Lau, M.K.; Compson, Z.G.; Wojtowicz, T.; Keith, A.R.; Zinkgraf, M.S.; Schweitzer, J.A.; Shuster, S.M.; Gehring, C.A.; et al. Tree Genotype Mediates Covariance among Communities from Microbes to Lichens and Arthropods. J. Ecol. 2015, 103, 840–850. [Google Scholar] [CrossRef]
- Schweitzer, J.A.; Bailey, J.K.; Fischer, D.G.; LeRoy, C.J.; Lonsdorf, E.V.; Whitham, T.G.; Hart, S.C. Plant–Soil–Microorganism Interactions: Heritable Relationship Between Plant Genotype and Associated Soil Microorganisms. Ecology 2008, 89, 773–781. [Google Scholar] [CrossRef]
- Lamit, L.J.; Wojtowicz, T.; Kovacs, Z.; Wooley, S.C.; Zinkgraf, M.; Whitham, T.G.; Lindroth, R.L.; Gehring, C.A. Hybridization among Foundation Tree Species Influences the Structure of Associated Understory Plant Communities. Botany 2011, 89, 165–174. [Google Scholar] [CrossRef]
- Johnson, M.T.J.; Agrawal, A.A. Plant Genotype and Environment Interact to Shape a Diverse Arthropod Community on Evening Primrose (Oenothera biennis). Ecology 2005, 86, 874–885. [Google Scholar] [CrossRef]
- Bangert, R.K.; Lonsdorf, E.V.; Wimp, G.M.; Shuster, S.M.; Fischer, D.; Schweitzer, J.A.; Allan, G.J.; Bailey, J.K.; Whitham, T.G. Genetic Structure of a Foundation Species: Scaling Community Phenotypes from the Individual to the Region. Heredity 2008, 100, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Busby, P.E.; Newcombe, G.; Dirzo, R.; Whitham, T.G. Differentiating Genetic and Environmental Drivers of Plant–Pathogen Community Interactions. J. Ecol. 2014, 102, 1300–1309. [Google Scholar] [CrossRef]
- Barbour, R.C.; O’Reilly-Wapstra, J.M.; Little, D.W.D.; Jordan, G.J.; Steane, D.A.; Humphreys, J.R.; Bailey, J.K.; Whitham, T.G.; Potts, B.M. A Geographic Mosaic of Genetic Variation within a Foundation Tree Species and Its Community-Level Consequences. Ecology 2009, 90, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Zytynska, S.E.; Khudr, M.S.; Harris, E.; Preziosi, R.F. Genetic Effects of Tank-Forming Bromeliads on the Associated Invertebrate Community in a Tropical Forest Ecosystem. Oecologia 2012, 170, 467–475. [Google Scholar] [CrossRef]
- Bangert, R.K.; Allan, G.J.; Turek, R.J.; Wimp, G.M.; Meneses, N.; Martinsen, G.D.; Keim, P.; Whitham, T.G. From Genes to Geography: A Genetic Similarity Rule for Arthropod Community Structure at Multiple Geographic Scales. Mol. Ecol. 2006, 15, 4215–4228. [Google Scholar] [CrossRef]
- Bothwell, H.M.; Cushman, S.A.; Woolbright, S.A.; Hersch-Green, E.I.; Evans, L.M.; Whitham, T.G.; Allan, G.J. Conserving Threatened Riparian Ecosystems in the American West: Precipitation Gradients and River Networks Drive Genetic Connectivity and Diversity in a Foundation Riparian Tree (Populus angustifolia). Mol. Ecol. 2017, 26, 5114–5132. [Google Scholar] [CrossRef]
- Evans, L.M.; Kaluthota, S.; Pearce, D.W.; Allan, G.J.; Floate, K.; Rood, S.B.; Whitham, T.G. Bud Phenology and Growth Are Subject to Divergent Selection across a Latitudinal Gradient in Populus Angustifolia and Impact Adaptation across the Distributional Range and Associated Arthropods. Ecol. Evol. 2016, 6, 4565–4581. [Google Scholar] [CrossRef]
- Omernik, J.M. Ecoregions of the Conterminous United States. Ann. Assoc. Am. Geogr. 1987, 77, 118–125. [Google Scholar] [CrossRef]
- Omernik, J.M.; Griffith, G.E. Ecoregions of the Conterminous United States: Evolution of a Hierarchical Spatial Framework. Environ. Manag. 2014, 54, 1249–1266. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.R.; Letten, A.D.; Ke, P.-J.; Anderson, C.B.; Hendershot, J.N.; Dhami, M.K.; Dlott, G.A.; Grainger, T.N.; Howard, M.E.; Morrison, B.M.L.; et al. A Global Test of Ecoregions. Nat. Ecol. Evol. 2018, 2, 1889–1896. [Google Scholar] [CrossRef]
- Turesson, G. The Genotypical Response of the Plant Species to the Habitat. Hereditas 1922, 3, 211–350. [Google Scholar] [CrossRef]
- Hufford, K.M.; Mazer, S.J. Plant Ecotypes: Genetic Differentiation in the Age of Ecological Restoration. Trends Ecol. Evol. 2003, 18, 147–155. [Google Scholar] [CrossRef]
- Ikeda, D.H.; Max, T.L.; Allan, G.J.; Lau, M.K.; Shuster, S.M.; Whitham, T.G. Genetically Informed Ecological Niche Models Improve Climate Change Predictions. Glob. Chang. Biol. 2017, 23, 164–176. [Google Scholar] [CrossRef]
- Cooper, H.F.; Grady, K.C.; Cowan, J.A.; Best, R.J.; Allan, G.J.; Whitham, T.G. Genotypic Variation in Phenological Plasticity: Reciprocal Common Gardens Reveal Adaptive Responses to Warmer Springs but Not to Fall Frost. Glob. Chang. Biol. 2019, 25, 187–200. [Google Scholar] [CrossRef]
- Bothwell, H.M.; Evans, L.M.; Hersch-Green, E.I.; Woolbright, S.A.; Allan, G.J.; Whitham, T.G. Genetic Data Improves Niche Model Discrimination and Alters the Direction and Magnitude of Climate Change Forecasts. Ecol. Appl. 2021, 31, e02254. [Google Scholar] [CrossRef]
- Blasini, D.E.; Koepke, D.F.; Grady, K.C.; Allan, G.J.; Gehring, C.A.; Whitham, T.G.; Cushman, S.A.; Hultine, K.R. Adaptive Trait Syndromes along Multiple Economic Spectra Define Cold and Warm Adapted Ecotypes in a Widely Distributed Foundation Tree Species. J. Ecol. 2021, 109, 1298–1318. [Google Scholar] [CrossRef]
- Yang, J.; Cushman, S.A.; Song, X.; Yang, J.; Zhang, P. Genetic Diversity and Drivers of Genetic Differentiation of Reaumuria Soongorica of the Inner Mongolia Plateau in China. Plant Ecol. 2015, 216, 925–937. [Google Scholar] [CrossRef]
- Germino, M.J.; Moser, A.M.; Sands, A.R. Adaptive Variation, Including Local Adaptation, Requires Decades to Become Evident in Common Gardens. Ecol. Appl. 2019, 29, e01842. [Google Scholar] [CrossRef] [PubMed]
- Lovell, J.T.; MacQueen, A.H.; Mamidi, S.; Bonnette, J.; Jenkins, J.; Napier, J.D.; Sreedasyam, A.; Healey, A.; Session, A.; Shu, S.; et al. Genomic Mechanisms of Climate Adaptation in Polyploid Bioenergy Switchgrass. Nature 2021, 590, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Raszick, T.J.; Song, H. The Ecotype Paradigm: Testing the Concept in an Ecologically Divergent Grasshopper. Insect Syst. Evol. 2016, 47, 363–387. [Google Scholar] [CrossRef]
- Leonhardt, S.; Hoppe, B.; Stengel, E.; Noll, L.; Moll, J.; Bässler, C.; Dahl, A.; Buscot, F.; Hofrichter, M.; Kellner, H. Molecular Fungal Community and Its Decomposition Activity in Sapwood and Heartwood of 13 Temperate European Tree Species. PLoS ONE 2019, 14, e0212120. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Schemske, D.W. Ecological and Evolutionary Perspectives on Community Assembly. Trends Ecol. Evol. 2015, 30, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Cushman, S.A.; Max, T.; Meneses, N.; Evans, L.M.; Ferrier, S.; Honchak, B.; Whitham, T.G.; Allan, G.J. Landscape Genetic Connectivity in a Riparian Foundation Tree Is Jointly Driven by Climatic Gradients and River Networks. Ecol. Appl. 2014, 24, 1000–1014. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.; Deckert, R.; Schweitzer, J.; Rehill, B.; Lindroth, R.; Gehring, C.; Whitham, T. Host Plant Genetics Affect Hidden Ecological Players: Links among Populus, Condensed Tannins, and Fungal Endophyte Infection. Can. J. Bot. 2005, 83, 356–361. [Google Scholar] [CrossRef]
- Rehill, B.; Clauss, A.; Wieczorek, L.; Whitham, T.; Lindroth, R. Foliar Phenolic Glycosides from Populus Fremontii, Populus Angustifolia, and Their Hybrids. Biochem. Syst. Ecol. 2005, 33, 125–131. [Google Scholar] [CrossRef]
- Grady, K.C.; Laughlin, D.C.; Ferrier, S.M.; Kolb, T.E.; Hart, S.C.; Allan, G.J.; Whitham, T.G. Conservative Leaf Economic Traits Correlate with Fast Growth of Genotypes of a Foundation Riparian Species near the Thermal Maximum Extent of Its Geographic Range. Funct. Ecol. 2013, 27, 428–438. [Google Scholar] [CrossRef]
- Fischer, D.G.; Wimp, G.M.; Hersch-Green, E.; Bangert, R.K.; LeRoy, C.J.; Bailey, J.K.; Schweitzer, J.A.; Dirks, C.; Hart, S.C.; Allan, G.J.; et al. Tree Genetics Strongly Affect Forest Productivity, but Intraspecific Diversity–Productivity Relationships Do Not. Funct. Ecol. 2017, 31, 520–529. [Google Scholar] [CrossRef]
- Cooper, H.F.; Best, R.J.; Andrews, L.V.; Corbin, J.P.M.; Garthwaite, I.; Grady, K.C.; Gehring, C.A.; Hultine, K.R.; Whitham, T.G.; Allan, G.J. Evidence of Climate-Driven Selection on Tree Traits and Trait Plasticity across the Climatic Range of a Riparian Foundation Species. Mol. Ecol. 2022, 31, 5024–5040. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Dey, A.; Kumar, V.; Batiha, G.E.-S.; El-Esawi, M.A.; Tomczyk, M.; Ray, P. Fungal Endophyte: An Interactive Endosymbiont with the Capability of Modulating Host Physiology in Myriad Ways. Front. Plant Sci. 2021, 12, 1–19. [Google Scholar] [CrossRef]
- Rollinson, C.R.; Finley, A.O.; Alexander, M.R.; Banerjee, S.; Dixon Hamil, K.-A.; Koenig, L.E.; Locke, D.H.; DeMarche, M.L.; Tingley, M.W.; Wheeler, K.; et al. Working across Space and Time: Nonstationarity in Ecological Research and Application. Front. Ecol. Environ. 2021, 19, 66–72. [Google Scholar] [CrossRef]
- Thompson, C.M.; McGarigal, K. The Influence of Research Scale on Bald Eagle Habitat Selection along the Lower Hudson River, New York (USA). Landsc. Ecol. 2002, 17, 569–586. [Google Scholar] [CrossRef]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double Digest RADseq: An Inexpensive Method for De Novo SNP Discovery and Genotyping in Model and Non-Model Species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef]
- Rohland, N.; Reich, D. Cost-Effective High-Throughput DNA Sequencing Libraries. Genome Res. 2012, 22, 939–946. [Google Scholar] [CrossRef]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An Analysis Tool Set for Population Genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [PubMed]
- Andrews, L.V. Akutils RADseq Utility: Simplified Processing of RADseq Data through Stacks. 2018. Available online: https://zenodo.org/record/1205089#.ZEv4ZebMJ3g (accessed on 28 April 2023).
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.I.; Hefer, C.A.; Kolosova, N.; Douglas, C.J.; Cronk, Q.C.B. Whole Plastome Sequencing Reveals Deep Plastid Divergence and Cytonuclear Discordance between Closely Related Balsam Poplars, Populus Balsamifera and P. Trichocarpa (Salicaceae). New Phytol. 2014, 204, 693–703. [Google Scholar] [CrossRef]
- Kersten, B.; Faivre Rampant, P.; Mader, M.; Le Paslier, M.-C.; Bounon, R.; Berard, A.; Vettori, C.; Schroeder, H.; Leplé, J.-C.; Fladung, M. Genome Sequences of Populus Tremula Chloroplast and Mitochondrion: Implications for Holistic Poplar Breeding. PLoS ONE 2016, 11, e0147209. [Google Scholar] [CrossRef]
- Taylor, D.L.; Walters, W.A.; Lennon, N.J.; Bochicchio, J.; Krohn, A.; Caporaso, J.G.; Pennanen, T. Accurate Estimation of Fungal Diversity and Abundance through Improved Lineage-Specific Primers Optimized for Illumina Amplicon Sequencing. Appl. Environ. Microbiol. 2016, 82, 7217–7226. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, P.; Teixeira, M.D.M.; Andrews, L.; Fernandez, A.; Santander, G.; Doyle, A.; Perez, M.; Yegres, F.; Barker, B.M. Detection of Coccidioides Posadasii from Xerophytic Environments in Venezuela Reveals Risk of Naturally Acquired Coccidioidomycosis Infections. Emerg. Microbes Infect. 2018, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Tedersoo, L.; Ryberg, M.; Kristiansson, E.; Hartmann, M.; Unterseher, M.; Porter, T.M.; Bengtsson-Palme, J.; Walker, D.M.; de Sousa, F.; et al. A Comprehensive, Automatically Updated Fungal ITS Sequence Dataset for Reference-Based Chimera Control in Environmental Sequencing Efforts. Microbes Environ. 2015, 30, 145–150. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and Fast Clustering Method for Amplicon-Based Studies. PeerJ 2014, 2, e593. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a Unified Paradigm for Sequence-Based Identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-Filtering Vastly Improves Diversity Estimates from Illumina Amplicon Sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential Abundance Analysis for Microbial Marker-Gene Surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef]
- Zheng, X.; Levine, D.; Shen, J.; Gogarten, S.M.; Laurie, C.; Weir, B.S. A High-Performance Computing Toolset for Relatedness and Principal Component Analysis of SNP Data. Bioinformatics 2012, 28, 3326–3328. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation: Vienna, Austria, 2020. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of Population Structure Using Multilocus Genotype Data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software Structure: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A Cluster Matching and Permutation Program for Dealing with Label Switching and Multimodality in Analysis of Population Structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Rosenberg, N.A. DISTRUCT: A Program for the Graphical Display of Population Structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.-E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Mesinger, F.; DiMego, G.; Kalnay, E.; Mitchell, K.; Shafran, P.C.; Ebisuzaki, W.; Jović, D.; Woollen, J.; Rogers, E.; Berbery, E.H.; et al. North American Regional Reanalysis. Bull. Am. Meteorol. Soc. 2006, 87, 343–360. [Google Scholar] [CrossRef]
- National Centers for Environmental Prediction/National Weather Service/NOAA/U.S. Department of Commerce NCEP North American Regional Reanalysis (NARR). Research Data Archive at the National Center for Atmospheric Research, Computational and Information Systems Laboratory. 2005. Available online: https://rda.ucar.edu/datasets/ds608.0/ (accessed on 16 April 2019).
- Böhner, J.; Koethe, R.; Conrad, O.; Gross, J.; Ringeler, A.; Selige, T. Soil Regionalisation by Means of Terrain Analysis and Process Parameterisation. In Soil Classifiction 2001; Research Report No. 7, EUR 20398 EN; European Soil Bureau: Luxembourg, 2001; pp. 213–222. [Google Scholar]
- Rood, S.B.; Samuelson, G.M.; Braatne, J.H.; Gourley, C.R.; Hughes, F.M.; Mahoney, J.M. Managing River Flows to Restore Floodplain Forests. Front. Ecol. Environ. 2005, 3, 193–201. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Goslee, S.C.; Urban, D.L. The Ecodist Package for Dissimilarity-Based Analysis of Ecological Data. J. Stat. Softw. 2007, 22, 1–19. [Google Scholar] [CrossRef]
- Shirk, A.J.; Wallin, D.O.; Cushman, S.A.; Rice, C.G.; Warheit, K.I. Inferring Landscape Effects on Gene Flow: A New Model Selection Framework. Mol. Ecol. 2010, 19, 3603–3619. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data, V7. 2016. Available online: https://www.wildblueberrymedia.net/pcord (accessed on 27 April 2021).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package v2.5-6. 2019. Available online: https://cran.r-project.org/web/packages/vegan/ (accessed on 22 February 2018).
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Hultine, K.R.; Allan, G.J.; Blasini, D.; Bothwell, H.M.; Cadmus, A.; Cooper, H.F.; Doughty, C.E.; Gehring, C.A.; Gitlin, A.R.; Grady, K.C.; et al. Adaptive Capacity in the Foundation Tree Species Populus Fremontii: Implications for Resilience to Climate Change and Non-Native Species Invasion in the American Southwest. Conserv. Physiol. 2020, 8, coaa061. [Google Scholar] [CrossRef]
- Wimp, G.M.; Martinsen, G.D.; Floate, K.D.; Bangert, R.K.; Whitham, T.G. Plant Genetic Determinants of Arthropod Community Structure and Diversity. Evolution 2005, 59, 61–69. [Google Scholar] [CrossRef]
- Busby, P.E.; Newcombe, G.; Dirzo, R.; Whitham, T.G. Genetic Basis of Pathogen Community Structure for Foundation Tree Species in a Common Garden and in the Wild. J. Ecol. 2013, 101, 867–877. [Google Scholar] [CrossRef]
- Herfeld, C.; Ivanova, M. Introduction: First Principles in Science—Their Status and Justification. Synthese 2021, 198, S3297–S3308. [Google Scholar] [CrossRef]
- Thomas, J.A.; Telfer, M.; Roy, D.B.; Preston, C.D.; Greenwood, J.; Asher, J.; Fox, R.; Clarke, R.T.; Lawton, J.H. Comparative Losses of British Butterflies, Birds, and Plants and the Global Extinction Crisis. Science 2004, 303, 1879–1881. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 Percent Decline over 27 Years in Total Flying Insect Biomass in Protected Areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef]
- Banerjee, A.; Cornejo, J.; Bandopadhyay, R. Emergent Climate Change Impact throughout the World: Call for “Microbiome Conservation” before It’s Too Late. Biodivers. Conserv. 2020, 29, 345–348. [Google Scholar] [CrossRef]
- Poff, B.; Koestner, K.A.; Neary, D.G.; Merritt, D. Threats to Western United States Riparian Ecosystems: A Bibliography; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ft. Collins, CO, USA, 2012; p. 269.
- Noss, R.F.; Scott, J.M. Endangered Ecosystems of the United States: A Preliminary Assessment of Loss and Degradation; Biological Report 28; US Department of the Interior, National Biological Service: Washington, DC, USA, 1995.
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, Migration or Extirpation: Climate Change Outcomes for Tree Populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Ikeda, D.H.; Bothwell, H.M.; Lau, M.K.; O’Neill, G.A.; Grady, K.C.; Whitham, T.G. A Genetics-Based Universal Community Transfer Function for Predicting the Impacts of Climate Change on Future Communities. Funct. Ecol. 2014, 28, 65–74. [Google Scholar] [CrossRef]
- Ikeda, D.H.; Grady, K.C.; Shuster, S.M.; Whitham, T.G. Incorporating Climate Change and Exotic Species into Forecasts of Riparian Forest Distribution. PLoS ONE 2014, 9, e107037. [Google Scholar] [CrossRef] [PubMed]
- Prober, S.; Byrne, M.; McLean, E.; Steane, D.; Potts, B.; Vaillancourt, R.; Stock, W. Climate-Adjusted Provenancing: A Strategy for Climate-Resilient Ecological Restoration. Front. Ecol. Evol. 2015, 3, 65. [Google Scholar] [CrossRef]
- Aitken, S.N.; Whitlock, M.C. Assisted Gene Flow to Facilitate Local Adaptation to Climate Change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 367–388. [Google Scholar] [CrossRef]
- O’Neill, G.; TongLi, W.; Ukrainetz, N.; Charleson, L.; McAuley, L.; Yanchuk, A.; Zedel, S. A Proposed Climate-Based Seed Transfer System for British Columbia; Technical Report; Ministry of Forests Lands Natural Resource Operations: Victoria, BC, Canada, 2017.
- Keith, A.R.; Bailey, J.K.; Whitham, T.G. Assisted Migration Experiments along a Distance/Elevation Gradient Show Limits to Supporting Home Site Communities. PLoS Clim. 2023, accepted. [Google Scholar]
- Morris, J.R.; Perfecto, I. An Aggressive Nonconsumptive Effect Mediates Pest Control and Multipredator Interactions in a Coffee Agroecosystem. Ecol. Appl. 2022, 32, e2653. [Google Scholar] [CrossRef]
- Thompson, J.N. The Geographic Mosaic of Coevolution; University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | ENDOPHYTES | ARTHROPODS | |||||
|---|---|---|---|---|---|---|---|
| Geog | Climate | GenD | Geog | Climate | GenD | ||
| Geog | 0.16 ** | 0.09 † | 0.07 | Geog | 0.08 ** | 0.17 *** | −0.03 |
| Climate | 0.09 † | 0.16 ** | 0.04 | Climate | −0.02 | 0.18 *** | −0.05 |
| GenD | 0.15 ** | 0.14 * | 0.09 | GenD | 0.08 ** | 0.19 *** | −0.02 |
| (b) | |||||||
| Geog | Climate | GenD | Geog | Climate | GenD | ||
| Geog | 0.15 * | 0.01 | 0.10 | Geog | 0.44 *** | 0.24 * | −0.09 |
| Climate | 0.12 † | 0.09 | 0.16 † | Climate | 0.15 † | 0.47 *** | −0.16 |
| GenD | 0.05 | 0.07 | 0.17 * | GenD | 0.43 *** | 0.48 *** | 0.13 † |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bothwell, H.M.; Keith, A.R.; Cooper, H.F.; Hull, J.B.; Andrews, L.V.; Wehenkel, C.; Hultine, K.R.; Gehring, C.A.; Cushman, S.A.; Whitham, T.G.; et al. Microevolutionary Processes in a Foundation Tree Inform Macrosystem Patterns of Community Biodiversity and Structure. Forests 2023, 14, 943. https://doi.org/10.3390/f14050943
Bothwell HM, Keith AR, Cooper HF, Hull JB, Andrews LV, Wehenkel C, Hultine KR, Gehring CA, Cushman SA, Whitham TG, et al. Microevolutionary Processes in a Foundation Tree Inform Macrosystem Patterns of Community Biodiversity and Structure. Forests. 2023; 14(5):943. https://doi.org/10.3390/f14050943
Chicago/Turabian StyleBothwell, Helen M., Arthur R. Keith, Hillary F. Cooper, Julia B. Hull, Lela V. Andrews, Christian Wehenkel, Kevin R. Hultine, Catherine A. Gehring, Samuel A. Cushman, Thomas G. Whitham, and et al. 2023. "Microevolutionary Processes in a Foundation Tree Inform Macrosystem Patterns of Community Biodiversity and Structure" Forests 14, no. 5: 943. https://doi.org/10.3390/f14050943