
Species Composition and Seasonal Abundance of Predatory Mites (Acari: Phytoseiidae) Inhabiting Aesculus hippocastanum (Sapindaceae)


Abstract
:1. Introduction
2. Materials and Methods
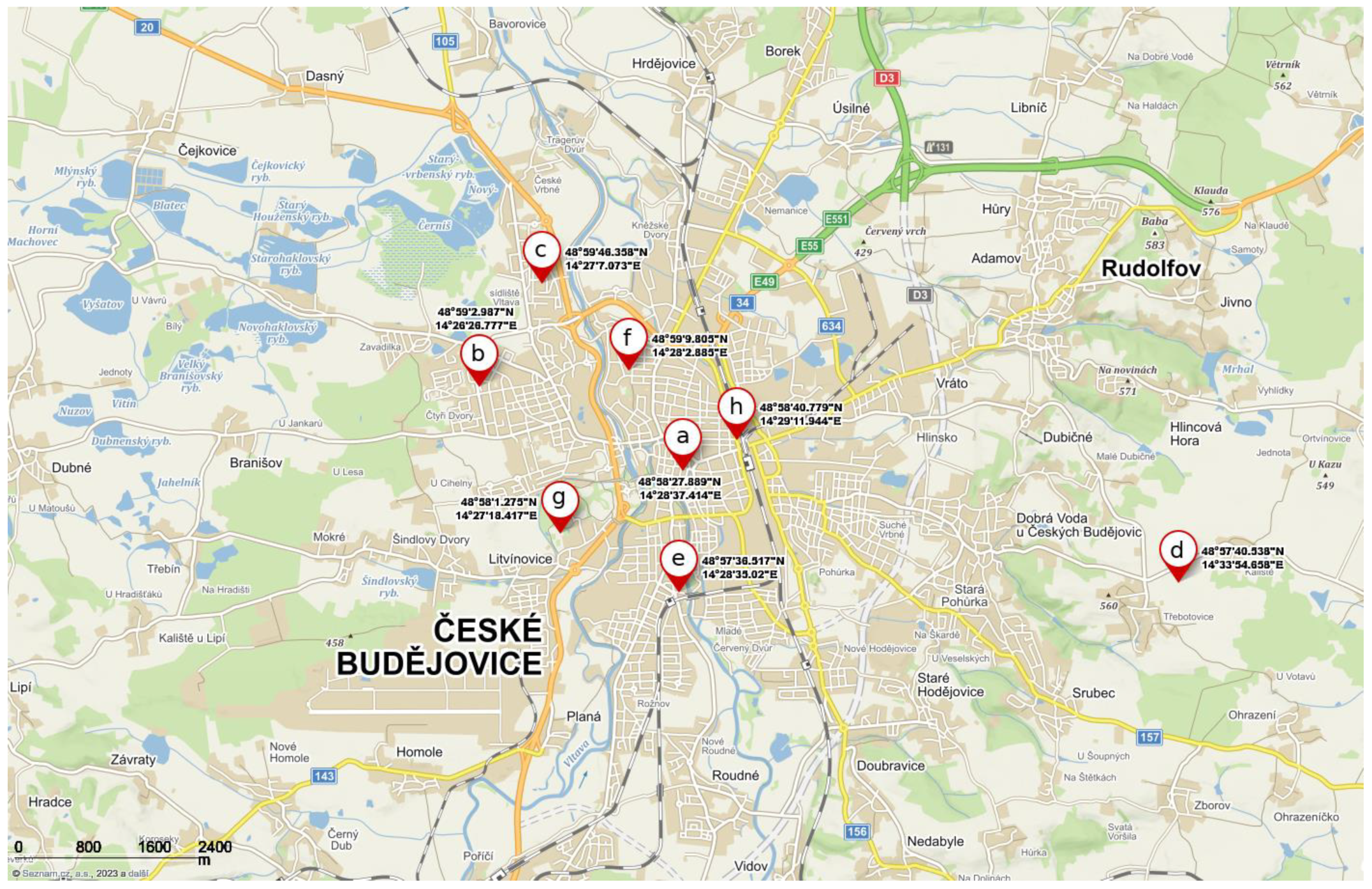
2.1. Sampling Sites
2.2. Sampling of Horse Chestnut Leaves
2.3. Collection and Identification of Phytoseiid Mites
2.4. Data Presentation and Statistical Analysis
3. Results
3.1. Species Composition
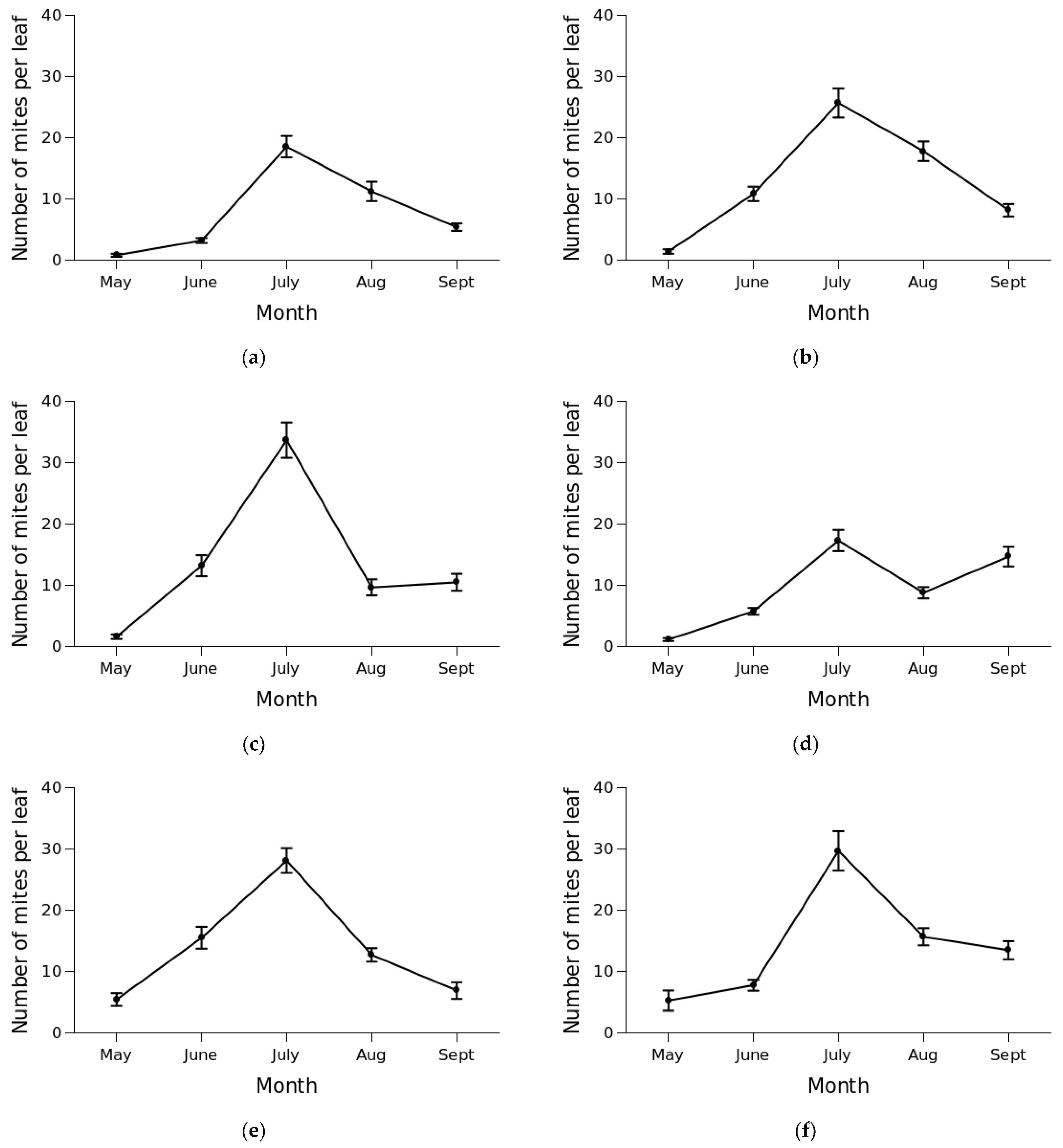
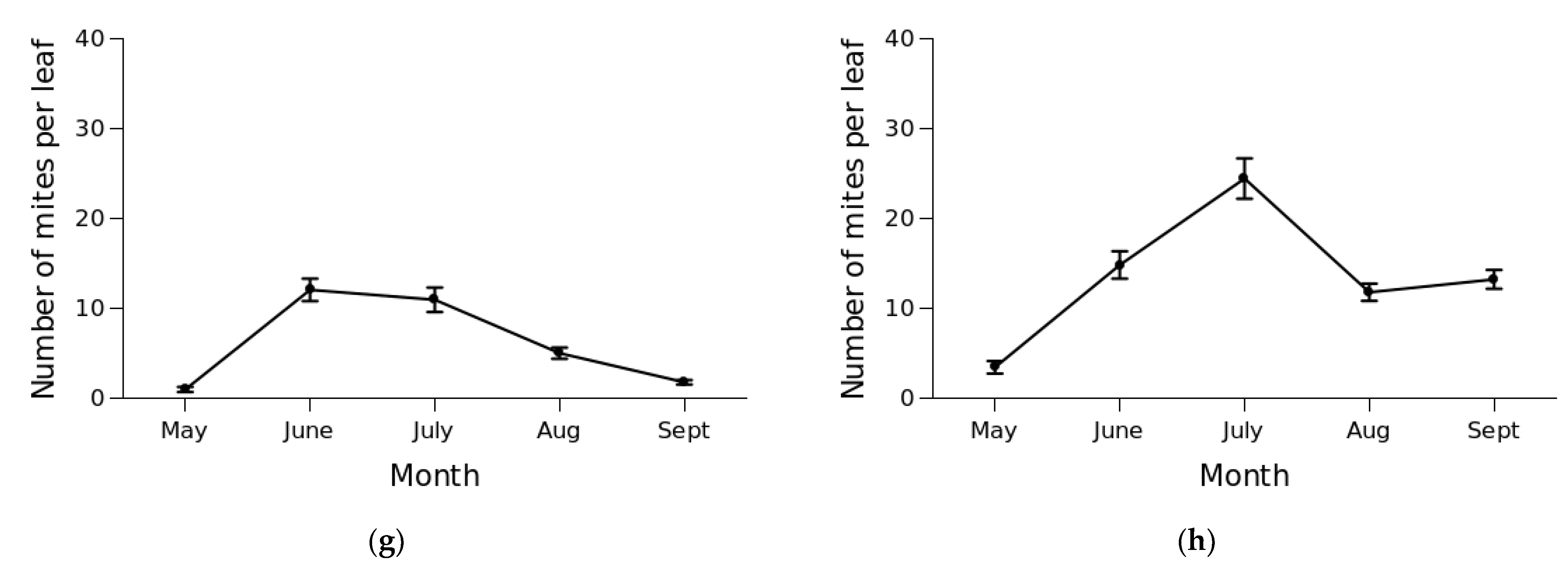
3.2. Mite Abundance and Seasonal Dynamics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, P.A.; Alhamd, O.; Iszkuło, G.; Dering, M.; Mukassabi, T.A. Biological Flora of the British Isles: Aesculus hippocastanum. J. Ecol. 2019, 107, 992–1030. [Google Scholar] [CrossRef]
- Ulbricht, C.; Tiffany, N.; Boon, H.; Ulbricht, C.; Basch, E.; Bent, S.; Barrette, E.P.; Smith, M.; Sollars, D.; Dennehy, C.; et al. Horse Chestnut: A Multidisciplinary Clinical Review. J. Herb. Pharmacother. 2002, 2, 71–85. [Google Scholar] [CrossRef]
- Shimada, T. Nutrient Compositions of Acorns and Horse Chestnuts in Relation to Seed-Hoarding: Nutrient Composition of Seeds. Ecol. Res. 2001, 16, 803–808. [Google Scholar] [CrossRef]
- Augustin, S.; Guichard, S.; Heitland, W.; Freise, J.; Svatoš, A.; Gilbert, M. Monitoring and Dispersal of the Invading Gracillariidae Cameraria Ohridella. J. Appl. Entomol. 2009, 133, 58–66. [Google Scholar] [CrossRef]
- Kirichenko, N.; Augustin, S.; Kenis, M. Invasive Leafminers on Woody Plants: A Global Review of Pathways, Impact, and Management. J. Pest Sci. 2019, 92, 93–106. [Google Scholar] [CrossRef]
- Musolin, D.L.; Kirichenko, N.I.; Karpun, N.N.; Aksenenko, E.V.; Golub, V.B.; Kerchev, I.A.; Mandelshtam, M.Y.; Vasaitis, R.; Volkovitsh, M.G.; Zhuravleva, E.N.; et al. Invasive Insect Pests of Forests and Urban Trees in Russia: Origin, Pathways, Damage, and Management. Forests 2022, 13, 521. [Google Scholar] [CrossRef]
- Kopačka, M.; Nachman, G.; Zemek, R. Seasonal Changes and the Interaction between the Horse Chestnut Leaf Miner Cameraria ohridella and Horse Chestnut Leaf Blotch Disease Caused by Guignardia aesculi. Forests 2021, 12, 952. [Google Scholar] [CrossRef]
- Somme, L.; Moquet, L.; Quinet, M.; Vanderplanck, M.; Michez, D.; Lognay, G.; Jacquemart, A.-L. Food in a Row: Urban Trees Offer Valuable Floral Resources to Pollinating Insects. Urban Ecosyst. 2016, 19, 1149–1161. [Google Scholar] [CrossRef]
- Tuovinen, T.; Rokx, J.A.H. Phytoseiid Mites (Acari: Phytoseiidae) on Apple Trees and in Surrounding Vegetation in Southern Finland. Densities and Species Composition. Exp. Appl. Acarol. 1991, 12, 35–46. [Google Scholar] [CrossRef]
- Tuovinen, T. Influence of Surrounding Trees and Bushes on the Phytoseiid Mite Fauna on Apple Orchard Trees in Finland. Agric. Ecosyst. Environ. 1994, 50, 39–47. [Google Scholar] [CrossRef]
- Kopačka, M.; Stathakis, T.I.; Broufas, G.; Papadoulis, G.T.; Zemek, R. Diversity and Abundance of Phytoseiidae (Acari: Mesostigmata) on Horse Chestnut (Aesculus hippocastanum L.) in an Urban Environment: A Comparison between Greece and the Czech Republic. Acarologia 2018, 58, 83–90. [Google Scholar] [CrossRef]
- Kabíček, J.; Řeháková, M. Phytoseiid Mite Community on Aesculus Hippocastanum in the Parks. Acta Fytotech. Zootech. 2004, 7, 114–115. [Google Scholar]
- Fidelis, E.G.; Farias, E.d.S.; Joaquim Da Silva Junior, R. Contribution of Natural Enemies and Weather to the Population Buildup of Raoiella Indica on Coconut. Syst. Appl. Acarol. 2022, 27, 8. [Google Scholar] [CrossRef]
- Tixier, M.-S.; Arnaud, A.; Douin, M.; Kreiter, S. Effects of Agroforestry on Phytoseiidae Communities (Acari: Mesostigmata) in Vineyards. A Synthesis of a 10-Year Period of Observations. Acarologia 2015, 55, 361–375. [Google Scholar] [CrossRef]
- Volter, L.; Prenerová, E.; Weyda, F.; Zemek, R. Changes in the Parasitism Rate and Parasitoid Community Structure of the Horse Chestnut Leafminer, Cameraria Ohridella (Lepidoptera: Gracillariidae), in the Czech Republic. Forests 2022, 13, 885. [Google Scholar] [CrossRef]
- Seniczak, S.; Dabrowski, J.; Długosz, J. Effect of Copper Smelting Air Pollution on the Mites (Acari) Associated with Young Scots Pine Forests Polluted by a Copper Smelting Works at Giogów, Poland. I. Arboreal Mites. Water Air Soil Pollut. 1997, 94, 71–84. [Google Scholar] [CrossRef]
- Kopacka, M.; Zemek, R. Spatial Variability in the Level of Infestation of the Leaves of Horse Chestnut by the Horse Chestnut Leaf Miner, Cameraria ohridella (Lepidoptera: Gracillariidae) and in the Number of Adult Moths and Parasitoids Emerging from Leaf Litter in an Urban Environment. Eur. J. Entomol. 2017, 114, 42–52. [Google Scholar] [CrossRef]
- Jura, S. Určování Stáří Stromů. Silva Bohem. 2001, 1, 19–21. [Google Scholar]
- Mapa Znečištění Ovzduší. Available online: https://geoportal.kraj-jihocesky.gov.cz/portal/mapy/zivotni-prostredi (accessed on 15 February 2023).
- Zacharda, M.; Pultar, O.; Muška, J. Washing Technique for Monitoring Mites in Apple Orchards. Exp. Appl. Acarol. 1988, 5, 181–183. [Google Scholar] [CrossRef]
- Beglyarov, G.A. Opredelitel chiscnych klescej fitoseiid (Parasitiformes, Phytoseiidae) fauny SSSR. Info Bjull IOBC 1981, 3, 1–45. [Google Scholar]
- Miedema, E. Survey of Phytoseiid Mites (Acari: Phytoseiidae) in Orchards and Surrounding Vegetation of Northwestern Europe, Especially in the Netherlands. Keys, Descriptions and Figures. Neth. J. Plant Pathol. 1987, 93, 1–63. [Google Scholar] [CrossRef]
- Chant, D.A.; Shaul, E.Y. A World Review of the Soleiger Species Group in the Genus Typhlodromus Scheuten (Acarina: Phytoseiidae). Can. J. Zool. 1982, 60, 3021–3032. [Google Scholar] [CrossRef]
- Chant, D.A.; Yoshida-Shaul, E. A World Review of the Tiliarum Species Group in the Genus Typhlodromus Scheuten (Acari: Phytoseiidae). Can. J. Zool. 1989, 67, 1006–1046. [Google Scholar] [CrossRef]
- Karg, W. Acari (Acarina), Milben, Parasitiformes (Anactinochaeta), Cohors Gamasina Leach, Raubmilben. In Tierwelt Deutschlands und der angrenzenden Meeresteile nach ihren Merkmalen und nach ihrer Lebensweise, 2nd ed.; VEB Gustav Fischer Verlag: Jena, Germany, 1993. [Google Scholar]
- Dajoz, R. Introduction to Ecology; Hodder and Stoughton: London, UK, 1977; ISBN 978-0-340-16254-5. [Google Scholar]
- Air Pollution Index Online Calculator. Available online: https://drr.ikcest.org/app/sf020 (accessed on 27 February 2023).
- SAS Institute. SAS/STAT 14.3: User’s Guide; SAS Institute: Cary, NC, USA, 2017. [Google Scholar]
- Liang, K.-Y.; Zeger, S.L. Longitudinal Data Analysis Using Generalized Linear Models. Biometrika 1986, 73, 13–22. [Google Scholar] [CrossRef]
- Ragusa di Chiara, S.; Papaioannou-Souliotis, P.; Tsolakis, H.; Tsagkarakou, N. Acari Fitoseidi (Parasitiformes, Phytoseiidae) Della Grecia Associati a Piante Forestali a Diverse Altitudini. Boll. Zool. Agr. Bachicol. 1995, 27, 85–91. [Google Scholar]
- McMurtry, J.A.; Croft, B.A. Life-Styles of Phytoseiid Mites and Their Roles in Biological Control. Annu. Rev. Entomol. 1997, 42, 291–321. [Google Scholar] [CrossRef]
- Kabíček, J. Broad Leaf Trees as Reservoirs for Phytoseiid Mites (Acari: Phytoseiidae). Plant Prot. Sci. 2003, 39, 65–69. [Google Scholar] [CrossRef]
- Kabíček, J. Intraleaf Distribution of the Phytoseiid Mites (Acari, Phytoseiidae) on Several Species of Wild Broad Leaf Trees. Biologia 2005, 60, 523–528. [Google Scholar]
- Kabíček, J. Linden Trees Are Favourable Host Plants for Phytoseiid Generalists in Urban Environment. Balt. For. 2019, 25, 1. [Google Scholar] [CrossRef]
- Kabíček, J.; Povondrová, K. Phytoseiid Mite Communities on Urban Deciduous Trees. Acta Fytotech. Zootech. 2004, 7, 119–121. [Google Scholar]
- Omeri, I. Phytoseiid Mites (Parasitiformes, Phytoseiidae) on Plants in Trostyanets Dendrological Park (Ukraine). Vestn. Zool. 2009, 43, e-7–e-14. [Google Scholar] [CrossRef]
- Barbar, Z. Occurrence, Population Dynamics and Winter Phenology of Spider Mites and Their Phytoseiid Predators in a Citrus Orchard in Syria. Acarologia 2014, 54, 409–423. [Google Scholar] [CrossRef]
- Grabovska, S.L.; Kolodochka, L.A. Species Complexes of Predatory Phytoseiid Mites (Parasitiformes, Phytoseiidae) in Green Urban Plantations of Uman’ (Ukraine). Vestn. Zool. 2014, 48, 495–502. [Google Scholar] [CrossRef]
- Vieira de Souza, I.; Argolo, P.S.; Gondim Júnior, M.G.C.; de Moraes, G.J.; Bittencourt, M.A.L.; Oliveira, A.R. Phytoseiid Mites from Tropical Fruit Trees in Bahia State, Brazil (Acari, Phytoseiidae). ZK 2015, 533, 99–131. [Google Scholar] [CrossRef]
- Stojnić, B.; Mladenović, K.; Marčić, D. Spider Mites and Predatory Mites (Acari: Tetranychidae, Phytoseiidae) on Stone Fruit Trees ( Prunus Spp.) in Serbia. Int. J. Acarol. 2018, 44, 322–329. [Google Scholar] [CrossRef]
- Grabovska, S.L.; Mykolaiko, I.I.; Mykolaiko, V.P. Structure Patterns of Phytoseiid Mite Communities in Urban Plant Associations. Ukr. J. Ecol. 2017, 7, 179–186. [Google Scholar] [CrossRef]
- Grabovska, S.L.; Mykolaiko, I.I. Mites of Phytoseiidae (Acari, Parasitiformes) in Urban Vegetative Plantations. Ukr. J. Ecol. 2017, 7, 216–222. [Google Scholar] [CrossRef]
- Ripka, G. Checklist of the Phytoseiidae of Hungary (Acari: Mesostigmata). Folia Entomol. Hung. 2006, 67, 229–260. [Google Scholar]
- Ripka, G. New Data to the Knowledge on the Phytoseiid Fauna in Hungary (Acari: Mesostigmata). Acta Phytopathol. Entomol. Hung. 1998, 33, 395–405. [Google Scholar]
- Komlovszky, S.Z.I.; Jenser, G. The frequent occurrence of the predatory mites Amblyseius finlandicus Oudemans and Phytoseius plumifer. Növényvédelem 1987, 23, 193–201. [Google Scholar]
- Gyenis, K.; Pénzes, B.; Hegyi, T. Phytophagous and predatory mites on the horse chestnut tree. Növényvédelem 2005, 41, 143–148. [Google Scholar]
- Ripka, G.; de Lillo, E. New Data to the Knowledge on the Eriophyoid Fauna in Hungary (Acari: Eriophyoidea). Folia Entomol. Hung. 1997, 58, 147–157. [Google Scholar]
- Karban, R.; English-Loeb, G.; Walker, M.A.; Thaler, J. Abundance of Phytoseiid Mites on Vitis Species: Effects of Leaf Hairs, Domatia, Prey Abundance and Plant Phylogeny. Exp. Appl. Acarol. 1995, 19, 189–197. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Haratym, W. Leaf Micromorphology of Aesculus Hippocastanum L. and Damage Caused by Leaf-Mining Larvae of Cameraria Ohridella Deschka and Dimić. Acta Agrobot. 2012, 65, 25–34. [Google Scholar] [CrossRef]
- Muhammad, S.; Wuyts, K.; Samson, R. Atmospheric Net Particle Accumulation on 96 Plant Species with Contrasting Morphological and Anatomical Leaf Characteristics in a Common Garden Experiment. Atmos. Environ. 2019, 202, 328–344. [Google Scholar] [CrossRef]
- Overmeer, W.P.J.; Zon, A.Q. The Preference of Amblyseius Potentillae (Garman) (Acarina: Phytoseiidae) for Certain Plant Substrates; Griffiths, D., Bowman, C., Eds.; Ellis Horwood Limited, Chichester: Chichester, UK, 1984; pp. 591–596. [Google Scholar]
- O’Dowd, D.J.; Willson, M.F. Leaf Domatia and Mites on Australasian Plants: Ecological and Evolutionary Implications. Biol. J. Linn. Soc. 1989, 37, 191–236. [Google Scholar] [CrossRef]
- Walter, D.E.; O’Dowd, D.J. Leaves with Domatia Have More Mites. Ecology 1992, 73, 1514–1518. [Google Scholar] [CrossRef]
- Kreiter, S.; Tixier, M.-S.; Croft, B.A.; Auger, P.; Barret, D. Plants and Leaf Characteristics Influencing the Predaceous Mite Kampimodromus Aberrans (Acari: Phytoseiidae) in Habitats Surrounding Vineyards. Environ. Entomol. 2002, 31, 648–660. [Google Scholar] [CrossRef]
- Matos, C.H.C.; Pallini, A.; Chaves, F.F.; Schoereder, J.H.; Janssen, A. Do Domatia Mediate Mutualistic Interactions between Coffee Plants and Predatory Mites? Entomol. Exp. Appl. 2006, 118, 185–192. [Google Scholar] [CrossRef]
- Loughner, R.; Goldman, K.; Loeb, G.; Nyrop, J. Influence of Leaf Trichomes on Predatory Mite (Typhlodromus Pyri) Abundance in Grape Varieties. Exp. Appl. Acarol. 2008, 45, 111–122. [Google Scholar] [CrossRef]
- Loughner, R.; Wentworth, K.; Loeb, G.; Nyrop, J. Leaf Trichomes Influence Predatory Mite Densities through Dispersal Behavior. Entomol. Exp. Appl. 2010, 134, 78–88. [Google Scholar] [CrossRef]
- Loughner, R.; Wentworth, K.; Loeb, G.; Nyrop, J. Influence of Leaf Trichomes on Predatory Mite Density and Distribution in Plant Assemblages and Implications for Biological Control. Biol. Control 2010, 54, 255–262. [Google Scholar] [CrossRef]
- O’Connell, D.M.; Lee, W.G.; Monks, A.; Dickinson, K.J.M. Does Microhabitat Structure Affect Foliar Mite Assemblages? Ecol. Entomol. 2010, 35, 317–328. [Google Scholar] [CrossRef]
- Schmidt, R.A. Leaf Structures Affect Predatory Mites (Acari: Phytoseiidae) and Biological Control: A Review. Exp. Appl. Acarol. 2014, 62, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Barba, P.; Loughner, R.; Wentworth, K.; Nyrop, J.P.; Loeb, G.M.; Reisch, B.I. A QTL Associated with Leaf Trichome Traits Has a Major Influence on the Abundance of the Predatory Mite Typhlodromus Pyri in a Hybrid Grapevine Population. Hortic. Res. 2019, 6, 87. [Google Scholar] [CrossRef] [PubMed]
- Roda, A.; Nyrop, J.; English-Loeb, G. Leaf Pubescence Mediates the Abundance of Non-Prey Food and the Density of the Predatory Mite Typhlodromus Pyri. Exp. Appl. Acarol. 2003, 29, 193–211. [Google Scholar] [CrossRef]
- Kugler, H. Blütenökologie; Gustav Fischer Verlag: Stuttgart, Germany, 1970. [Google Scholar]
- Goleva, I.; Zebitz, C.P.W. Suitability of Different Pollen as Alternative Food for the Predatory Mite Amblyseius Swirskii (Acari, Phytoseiidae). Exp. Appl. Acarol. 2013, 61, 259–283. [Google Scholar] [CrossRef] [PubMed]
- Goleva, I.; Rubio Cadena, E.C.; Ranabhat, N.B.; Beckereit, C.; Zebitz, C.P.W. Dietary Effects on Body Weight of Predatory Mites (Acari, Phytoseiidae). Exp. Appl. Acarol. 2015, 66, 541–553. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Tietze, M.; Michońska, M. Ecological Features of the Flowers of Aesculus Hippocastanum L. and Characteristics of Aesculus L. Pollen Seasons under the Conditions of Central-Eastern Poland. Acta Agrobot. 2012, 65, 61–68. [Google Scholar] [CrossRef]
- Pekas, A.; Wäckers, F.L. Multiple Resource Supplements Synergistically Enhance Predatory Mite Populations. Oecologia 2017, 184, 479–484. [Google Scholar] [CrossRef]
- Pozzebon, A.; Loeb, G.M.; Duso, C. Role of Supplemental Foods and Habitat Structural Complexity in Persistence and Coexistence of Generalist Predatory Mites. Sci. Rep. 2015, 5, 14997. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, R.; Sakcali, S.; Yarci, C.; Aksoy, A.; Ozturk, M. Use of Aesculus Hippocastanum L. as a Biomonitor of Heavy Metal Pollution. Pak. J. Bot. 2006, 38, 1519–1527. [Google Scholar]
- Godinho, D.P.; Branquinho, C.; Magalhães, S. Intraspecific Variability in Herbivore Response to Elemental Defences Is Caused by the Metal Itself. J. Pest. Sci. 2023, 96, 797–806. [Google Scholar] [CrossRef]
- Raupp, M.J.; Shrewsbury, P.M.; Herms, D.A. Ecology of Herbivorous Arthropods in Urban Landscapes. Annu. Rev. Entomol. 2010, 55, 19–38. [Google Scholar] [CrossRef]
- Kropczynska, D.; Van De Vrie, M.; Tomczyk, A. Bionomics of Eotetranychus Tiliarium and Its Phytoseiid Predators. Exp. Appl. Acarol. 1988, 5, 65–81. [Google Scholar] [CrossRef]
- Kehrli, P.; Bacher, S. Date of Leaf Litter Removal to Prevent Emergence of Cameraria ohridella in the Following Spring. Entomol. Exp. Appl. 2003, 107, 159–162. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Geographical Coordinates of Geometric Centre | Greenery | Distance to Water 2 (m) | A. hippocastanum | ||
|---|---|---|---|---|---|---|
| Area (%) | Care 1 | Density (ha−1) | Age 3 (years; x̄ ± SE) | |||
| City center | 48.9744136 N, 14.4770594 E | 9.02 | 3 | 69 | 1.40 | 74.99 ± 1.86 |
| Šumava and Máj estate | 48.9841631 N, 14.4407714 E | 31.69 | 2 | 660 | 0.22 | 39.02 ± 3.19 |
| Vltava estate | 48.9962106 N, 14.4519647 E | 36.96 | 2 | 360 | 0.43 | 41.79 ± 3.49 |
| Třebotovice and Kaliště village | 48.9612606 N, 14.5651828 E | 6.67 | 1 | 700 | 0.17 | 17. 82 ± 6.86 |
| Rožnov estate | 48.9601436 N, 14.4763944 E | 9.86 | 2 | 110 | 0.19 | 70.49 ± 4.43 |
| Pražské předměstí estate | 48.9860569 N, 14.4674681 E | 29.84 | 2 | 338 | 0.40 | 49.74 ± 2.75 |
| Stromovka park | 48.9670208 N, 14.4551158 E | 40.00 | 2 | 102 | 0.30 | 43.53 ± 4.13 |
| Nádražní street | 48.9779942 N, 14.4866511 E | 7.69 | 0 | 920 | 1.38 | 77.74 ± 3.02 |
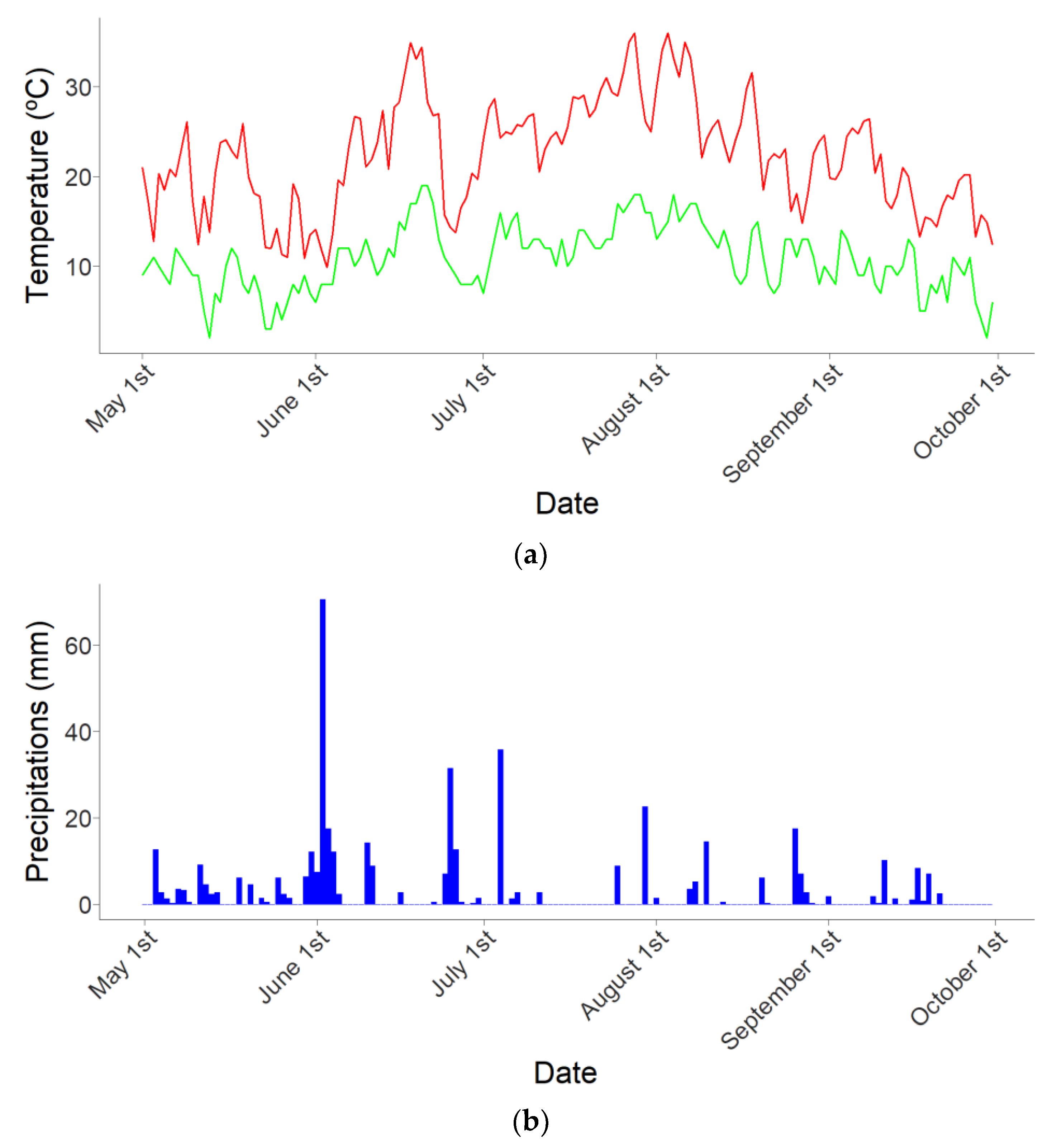
| Month | Temperature in °C (Mean ± SE) | Precipitations in mm |
|---|---|---|
| May | 12.95 ± 3.17 | 84.5 |
| June | 16.95 ± 4.96 | 190.0 |
| July | 20.35 ± 2.53 | 74.2 |
| August | 18.93 ± 3.77 | 59.5 |
| September | 13.80 ± 2.98 | 35.2 |
| Parameter | Estimate | Standard Error | 95% Confidence Limits | Z | p | |
|---|---|---|---|---|---|---|
| Intercept | −4.9278 | 0.5132 | −5.9336 | −3.9220 | −9.60 | <0.0001 |
| Greenery area | −0.0462 | 0.0026 | −0.0512 | −0.0411 | −17.93 | <0.0001 |
| Care intensity | −0.1688 | 0.0208 | −0.2096 | −0.1280 | −8.11 | <0.0001 |
| Distance to water | −0.0002 | 0.0001 | −0.0003 | −0.0000 | −2.26 | 00.0239 |
| Tree density | −0.5152 | 0.0160 | −0.5465 | −0.4838 | −32.20 | <0.0001 |
| Tree age | −0.0317 | 0.0030 | −0.0375 | −0.0258 | −10.64 | <0.0001 |
| Temperature | 0.1941 | 0.0183 | 0.1583 | 0.2300 | 10.62 | <0.0001 |
| Precipitations | −0.0009 | 0.0014 | −0.0037 | 0.0019 | −0.63 | 00.5257 |
| Air pollution index | 1.5798 | 0.0978 | 1.3881 | 1.7716 | 16.15 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopačka, M.; Zemek, R. Species Composition and Seasonal Abundance of Predatory Mites (Acari: Phytoseiidae) Inhabiting Aesculus hippocastanum (Sapindaceae). Forests 2023, 14, 942. https://doi.org/10.3390/f14050942
Kopačka M, Zemek R. Species Composition and Seasonal Abundance of Predatory Mites (Acari: Phytoseiidae) Inhabiting Aesculus hippocastanum (Sapindaceae). Forests. 2023; 14(5):942. https://doi.org/10.3390/f14050942
Chicago/Turabian StyleKopačka, Michal, and Rostislav Zemek. 2023. "Species Composition and Seasonal Abundance of Predatory Mites (Acari: Phytoseiidae) Inhabiting Aesculus hippocastanum (Sapindaceae)" Forests 14, no. 5: 942. https://doi.org/10.3390/f14050942