
Deadwood-Dwelling Beetles (Coleoptera: Eucnemidae) in a Beech Reserve: A Case Study from the Czech Republic

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Deadwood Variables
2.3. Beetle Sampling and Determination
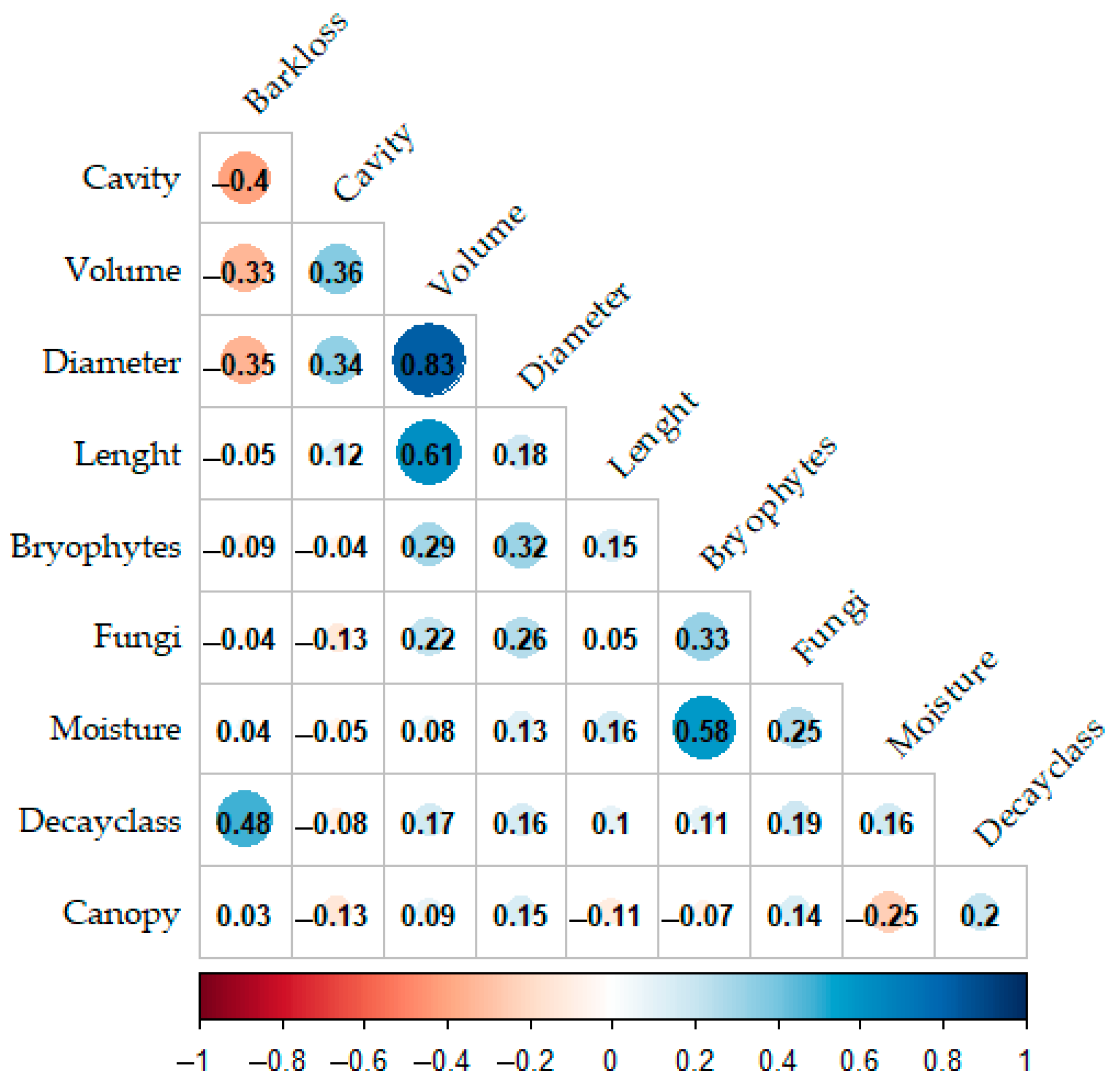
2.4. Data Analysis
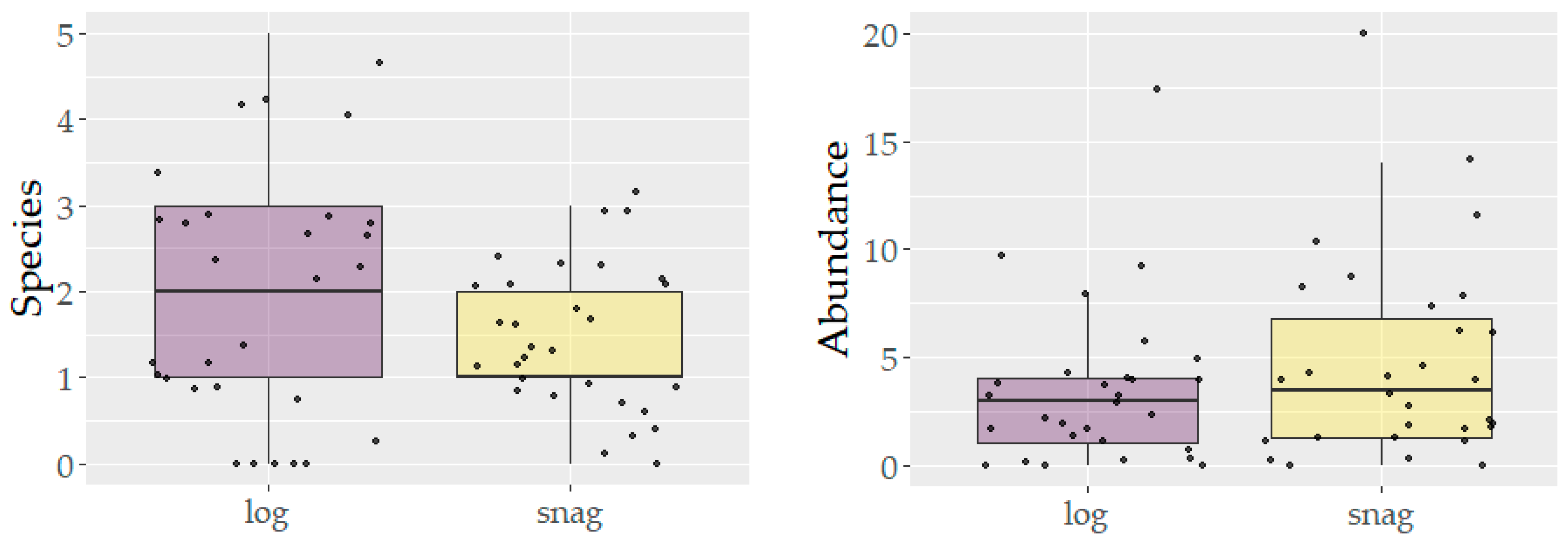
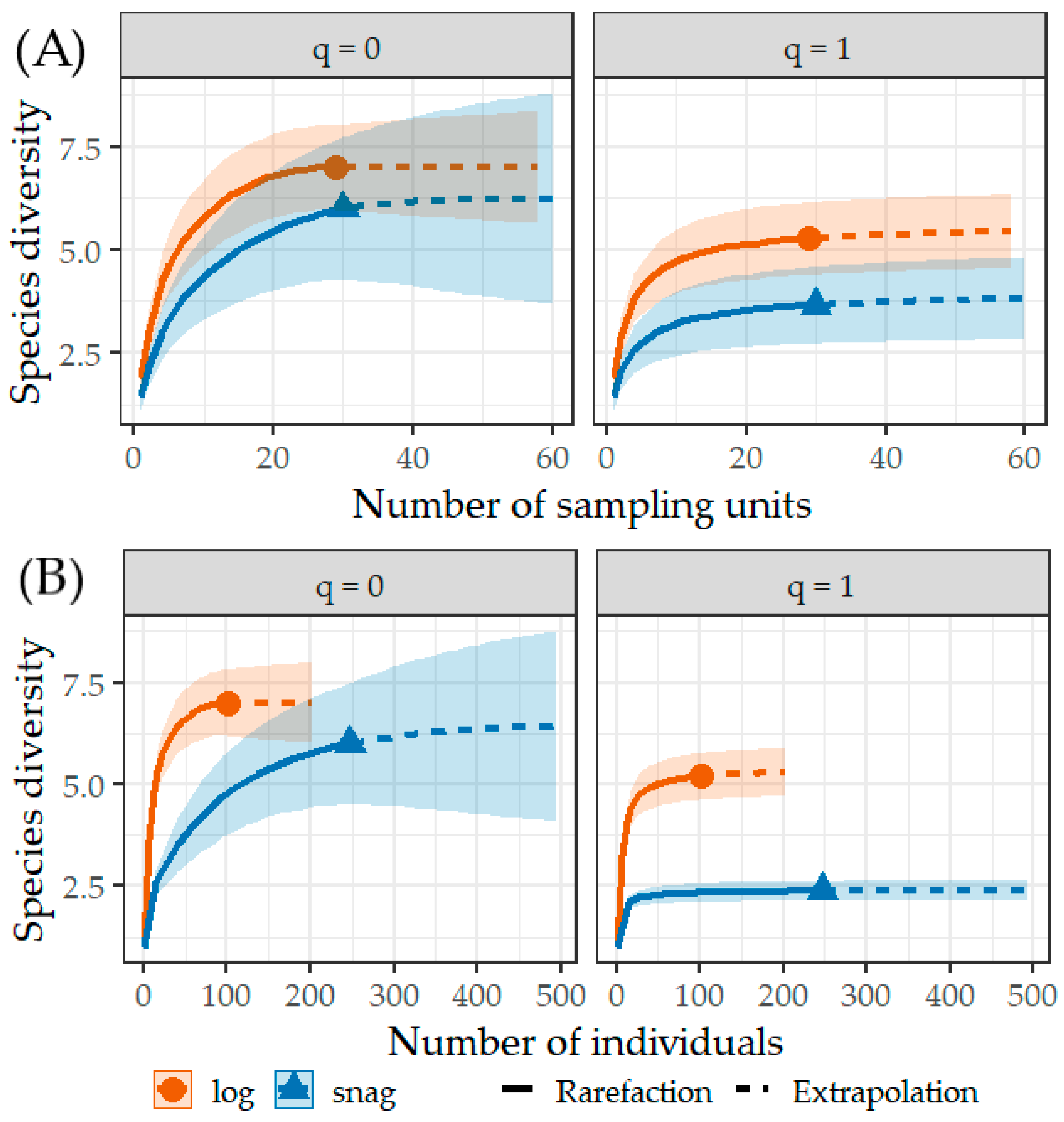
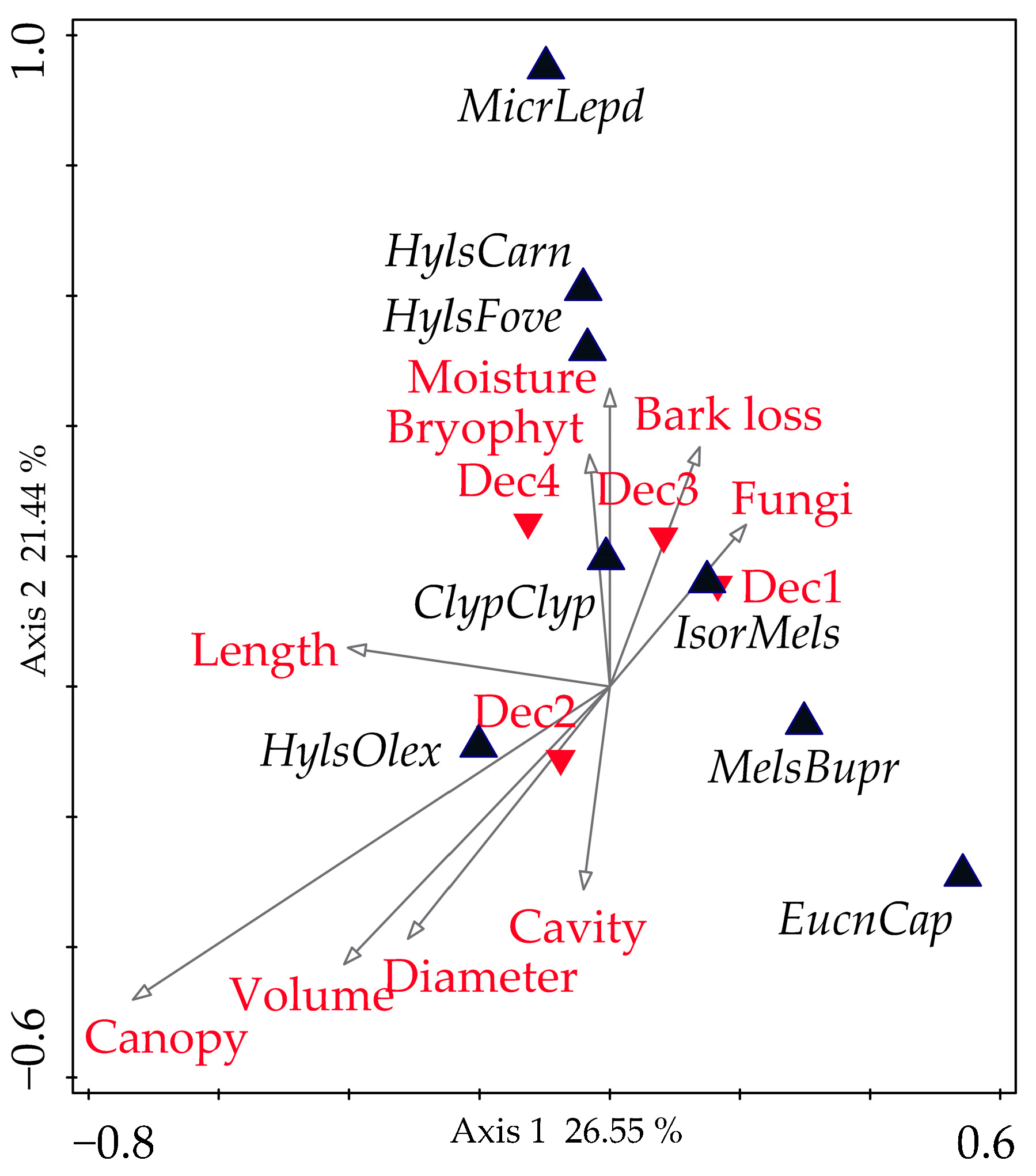
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oettel, J.; Lapin, K. Linking Forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol. Indic. 2021, 122, 107275. [Google Scholar] [CrossRef]
- Leidinger, J.; Blaschke, M.; Ehrhardt, M.; Fischer, A.; Gossner, M.M.; Jung, K.; Kienlein, S.; Kózak, J.; Michler, B.; Mosandl, R.; et al. Shifting tree species composition affects biodiversity of multiple taxa in Central European forests. For. Ecol. Manag. 2021, 498, 119552. [Google Scholar] [CrossRef]
- Schauer, B.; Thorn, S.; Blaschke, M.; Kudernatsch, T. Conversion of pure spruce to mixed spruce beech stands: Effects on alpha and beta diversity of multiple taxonomic groups. For. Ecol. Manag. 2023, 545, 121297. [Google Scholar] [CrossRef]
- Kostanjsek, F.; Sebek, P.; Baranova, B.; Seric Jelaska, L.; Riedl, V.; Cizek, L.; Didham, R.; Müller, J. Size matters! Habitat preferences of the wrinkled bark beetle, Rhysodes sulcatus, the relict species of European primeval forests. Insect Conserv. Divers. 2018, 11, 545–553. [Google Scholar] [CrossRef]
- Graf, M.; Seibold, S.; Gossner, M.M.; Hagge, J.; Weiß, I.; Bässler, C.; Müller, J. Coverage based diversity estimates of facultative saproxylic species highlight the importance of deadwood for biodiversity. For. Ecol. Manag. 2022, 517, 120275. [Google Scholar] [CrossRef]
- Zumr, V.; Nakládal, O.; Remeš, J.; Brestovanská, T.; Zumr, V. Diversity of click beetles in managed nonnative coniferous and native beech stands: Consequences of changes in the structural and species composition of tree stands in Central Europe. For. Ecosyst. 2022, 9, 100057. [Google Scholar] [CrossRef]
- Seibold, S.; Bässler, C.; Brandl, R.; Gossner, M.M.; Thorn, S.; Ulyshen, M.D.; Müller, J. Experimental studies of dead-wood biodiversity—A review identifying global gaps in knowledge. Biol. Conserv. 2015, 191, 139–149. [Google Scholar] [CrossRef]
- Speight, M.C.D. Saproxylic invertebrates and their conservation. Nat. Environ. Ser. 1989, 42, 1–79. [Google Scholar]
- Seibold, S.; Brandl, R.; Buse, J.; Hothorn, T.; Schmidl, J.; Thorn, S.; Müller, J. Association of extinction risk of saproxylic beetles with ecological degradation of forests in Europe. Conserv. Biol. 2015, 29, 382–390. [Google Scholar] [CrossRef]
- Parajuli, R.; Markwith, S.H. Quantity is foremost but quality matters: A global meta-analysis of correlations of dead wood volume and biodiversity in forest ecosystems. Biol. Conserv. 2023, 283, 110100. [Google Scholar] [CrossRef]
- Thorn, S.; Bässler, C.; Brandl, R.; Burton, P.J.; Cahall, R.; Campbell, J.L.; Castro, J.; Choi, C.-Y.; Cobb, T.; Donato, D.C.; et al. Impacts of salvage logging on biodiversity: A meta-analysis. J. Appl. Ecol. 2018, 55, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Muona, J. Eucnemidae. In Catalogue of Palaearctic Coleoptera: Elateroidea-Derodontoidea-Bostrichoidea-Lymexyloidea-Cleroidea-Cucujoidea; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2007; Volume 4, pp. 81–87. 935p. [Google Scholar]
- Vávra, J.; Škorpík, M. False click beetles (Coleoptera: Eucnemidae) in the Podyjí National Park and surrounding area, with notes to their bionomics. Thayensia 2013, 10, 53–90. [Google Scholar]
- Vávra, J. Eucnemidae. In Red List of Threatened Species of the Czech Republic; Hejda, R., Farkač, J., Chobot, K., Eds.; Agentura Ochrany Přírody a Krajiny České Republiky: Příroda, Praha, 2017; p. 355. [Google Scholar]
- Eckelt, A.; Müller, J.; Bense, U.; Brustel, H.; Bußler, H.; Chittaro, Y.; Cizek, L.; Frei, A.; Holzer, E.; Kadej, M.; et al. “Primeval Forest Relict Beetles” of Central Europe: A Set of 168 Umbrella Species for The Protection of Primeval Forest Remnants. J. Insect Conserv. 2018, 22, 15–28. [Google Scholar] [CrossRef]
- Nieto, A.; Alexander, K.N.A. European Red List of Saproxylic Beetles; Publications Office of the European Union: Luxembourg, 2010; p. 45. [Google Scholar]
- Mertlik, J. Faunistic Maps of Species of the Families Cerophytidae, Elateridae, Eucnemidae and Throscidae (Coleoptera: Elateroidea) of the Czech Republic and Slovakia.—Permanent Electronic Publication 2007–2023 (akt. 1.1.2023). 2023. Available online: https://www.elateridae.com/pag_uni.php?idp=19 (accessed on 6 January 2024).
- Hejda, R.; FarkaČ, J.; Chobot, K. Red List of Threatened Species of the Czech Republic; Agentura Ochrany Přírody a Krajiny České Republiky: Praha, Czech Republic, 2017; Volume 36, pp. 1–612. ISBN 978-80-88076-53-7. [Google Scholar]
- Foit, J. Faunistic records from the Czech Republic—436. Coleoptera: Eucnemidae. Klapalekiana 2018, 54, 4. [Google Scholar]
- Nakládal, O.; Vávra, C.J. Faunistic records from the Czech Republic—454. Coleoptera: Eucnemidae. Klapalekiana 2018, 54, 290. [Google Scholar]
- Bednařík, M. Otho Sphondyloides (Germar, 1818) (Coleoptera, Eucnemidae), a New Species for the Fauna of the Czech Republic. 2021. Available online: https://www.elateridae.com/elateridarium/page.php?idcl=335 (accessed on 3 January 2024).
- Müller, J.; Brunet, J.; Brin, A.; Bouget, C.; Brustel, H.; Bussler, H.; Foerster, B.; Isacsson, G.; Koehler, F.; Lachat, T.; et al. Implications from large-scale spatial diversity patterns of saproxylic beetles for the conservation of European Beech forests. Insect Conserv. Divers. 2013, 6, 162–169. [Google Scholar] [CrossRef]
- Nakládal, O.; Synek, J.; Zumr, V. Diurnal and Sex Ratio Flight Activity of Rare Cavity-Dweller Eucnemis capucina Ahrens, 1812 (Coleoptera: Eucnemidae) in Lowland Deciduous Forest. Forests 2023, 14, 720. [Google Scholar] [CrossRef]
- NCA CR. Species Occurrence Database; Nature Conservation Agency of the Czech Republic, NCA CR: Prague, Czech Republic, 2024; Available online: https://portal.nature.cz/publik_syst/ctihtmlpage.php?what=3&nabidka=hlavni (accessed on 19 January 2024).
- Care Plan for NPR Voděradské Bučiny 2021–2029; Agency for Nature Conservation and Landscape Protection of the Czech Republic: Prague, Czech Republic, 2023.
- Bílek, L.; Remeš, J.; Podrázský, V.; Rozenbergar, D.; Diaci, J.; Zahradník, D. Gap Regeneration in Near-Natural European Beech Forest Stands in Central Bohemia—The Role of Heterogeneity and Micro-Habitat Factors. Dendrobiology 2013, 71, 59–71. [Google Scholar] [CrossRef]
- Zumr, V.; Nakládal, O.; Bílek, L.; Remeš, J. The diameter of beech snags is an important factor for saproxylic beetle richness: Implications for forest management and conservation. For. Ecosyst. 2023, 10, 100143. [Google Scholar] [CrossRef]
- Šrámek, O. SPR Voděradské bučiny I. a II. Památky A Příroda 1983, 166–171, 241–248. [Google Scholar]
- Pokorný, J. Vývoj a druhová skladba černokosteleckých lesů do konce 18. století, Část II. Vývoj a obraz druhové skladby revíru Bohumile, Voděrady a Oplany. Sborník Lesn. Fak. Vysok. Školy Zemědělské V Praze 1963, 6, 31–107. [Google Scholar]
- Christensen, M.; Hahn, K.; Mountford, E.P.; Ódor, P.; Standovár, T.; Rozenbergar, D.; Diaci, J.; Wijdeven, S.; Meyer, P.; Winter, S.; et al. Dead wood in European beech (Fagus sylvatica) forest reserves. For. Ecol. Manag. 2005, 210, 267–282. [Google Scholar] [CrossRef]
- Bilek, L.; Remes, J.; Zahradnik, D. Managed vs. unmanaged. Structure of beech forest stands (Fagus sylvatica L.) after 50 years of development, Central Bohemia. For. Syst. 2011, 20, 122–138. [Google Scholar] [CrossRef]
- Brunet, J.; Isacsson, G. Influence of snag characteristics on saproxylic beetle assemblages in a south Swedish beech forest. J. Insect Conserv. 2009, 13, 515–528. [Google Scholar] [CrossRef]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap Light Analyzer (GLA), Version 2.0: Imaging Software to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye Photographs, User’s Manual and Program Documentation; Simon Fraser University/Institute of Ecosystem Studies: Burnaby/Millbrook, UK, 1999. [Google Scholar]
- Winter, S.; Möller, G.C. Microhabitats in lowland beech forests as monitoring tool for nature conservation. For. Ecol. Manag. 2008, 255, 1251–1261. [Google Scholar] [CrossRef]
- Gossner, M.; Lachat, T.; Brunet, J.; Isacsson, G.; Bouget, C.; Brustel, H.; Brandl, R.; Weisser, W.W.; Müller, J. Current near-to-nature forest management effects on functional trait composition of saproxylic beetles in beech forests. Conserv. Biol. 2013, 27, 605–614. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package ‘corrplot’: Visualization of a Correlation Matrix (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 5 January 2024).
- Wikars, L.-O.; Sahlin, E.; Ranius, T. A Comparison of Three Methods to Estimate Species Richness of Saproxylic Beetles (Coleoptera) in Logs and High Stumps of Norway Spruce. Can. Entomol. 2005, 137, 304–324. [Google Scholar] [CrossRef]
- Alinvi, O.; Ball, J.P.; Danell, K.; Hjältén, J.; Pettersson, R.B. Sampling Saproxylic Beetle Assemblages in Dead Wood Logs: Comparing Window and Eclector Traps to Traditional Bark Sieving and A Refinement. J. Insect Conserv. 2007, 11, 99–112. [Google Scholar] [CrossRef]
- Mertlik, J. The species of the family Melasidae (Coleoptera: Elateroidea) Czech and Slovak Republics. Elateridarium 2008, 2, 69–137. Available online: https://www.elateridae.com/elateridarium/page.php?idcl=88 (accessed on 1 September 2023).
- Schmidl, J.; Sausage, C.; Bussler, H. Red list and complete list of varieties of the “Diversicornia” (Coleoptera) of Germany. In Red List of Endangered Animals, Plants and Fungi of Germany, Volume 5: Invertebrates (Part 3); Nature Protection and Biodiversity; Ries, M., Balzer, S., Gruttke, H., Haupt, H., Hofbauer, N., Ludwig, G., Matzke-Hajek, G., Eds.; Agriculture Publisher: Münster, Germany, 2021; Volume 70, pp. 99–124. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 5 June 2023).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. Glmmtmb Balances Speed And Flexibility Among Packages For Zero-Inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: iNterpolation and EXTrapolation for Species Diversity. R Package Version 2022, 3.0.0. Available online: http://chao.stat.nthu.edu.tw/wordpress/software-download/ (accessed on 20 January 2024).
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef]
- Akaike, H. A Bayesian analysis of the minimum AIC procedure. Ann. Inst. Stat. Math. 1978, 30, 9–14. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where Are Europe’s Last Primary Forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef]
- Procházka, J.; Schlaghamerský, J. Does dead wood volume affect saproxylic beetles in montane beech-fir forests of Central Europe? J. Insect Conserv. 2019, 23, 157–173. [Google Scholar] [CrossRef]
- Müller, J.; Bußler, H.; Kneib, T. Saproxylic Beetle Assemblages Related to Silvicultural Management Intensity And Stand Structures In A Beech Forest In Southern Germany. J. Insect Conserv. 2008, 12, 107–124. [Google Scholar] [CrossRef]
- Müller, J.; Jarzabek-Müller, A.; Bussler, H.; Gossner, M.M. Hollow beech trees identified as keystone structures for saproxylic beetles by analyses of functional and phylogenetic diversity. Anim. Conserv. 2014, 17, 154–162. [Google Scholar] [CrossRef]
- Nakládal, O.; Zumr, V. Faunistic Records from the Czech Republic—526: Coleoptera: Eucnemidae. Klapalekiana 2022, 28, 112. [Google Scholar]
- Mertlik, J.; Pelikán, J. New data about Hylis olexai (Coleoptera: Eucnemidae) for the area of the Czech Republic and Slovakia. Elateridarium 2013, 7, 45–54. Available online: https://www.elateridae.com/elateridarium/page.php?idcl=207 (accessed on 2 January 2024).
- Lindman, L.; Öckinger, E.; Ranius, T. Microclimatic conditions mediate the effect of deadwood and forest characteristics on a threatened beetle species, Tragosoma depsarium. Oecologia 2022, 199, 737–752. [Google Scholar] [CrossRef] [PubMed]
- Bouget, C.; Nusillard, B.; Pineau, X.; Ricou, C. Effect of deadwood position on saproxylic beetles in temperate forests and conservation interest of oak snags. Insect Conserv. Divers. 2012, 5, 264–278. [Google Scholar] [CrossRef]
- Franc, N. Standing or downed dead trees—Does it matter for saproxylic beetles in temperate oak-rich forest? Can. J. For. Res. 2007, 37, 2494–2507. [Google Scholar] [CrossRef]
- Kappes, H.; Topp, W. Emergence of Coleoptera from deadwood in a managed broadleaved forest in central Europe. Biodivers. Conserv. 2004, 13, 1905–1924. [Google Scholar] [CrossRef]
- Hararuk, O.; Kurz, W.A.; Didion, M. Dynamics of dead wood decay in Swiss forests. For. Ecosyst. 2020, 7, 36. [Google Scholar] [CrossRef]
- Jonsell, M.; Weslien, J.; Ehnström, B. Substrate requirements of red-listed saproxylic invertebrates in Sweden. Biodivers. Conserv. 1998, 7, 749–764. [Google Scholar] [CrossRef]
- Floriancic, M.G.; Allen, S.T.; Meier, R.; Truniger, L.; Kirchner, J.W.; Molnar, P. Potential for Significant Precipitation Cycling by Forest-Floor Litter and Deadwood. Ecohydrology 2023, 16, e2493. [Google Scholar] [CrossRef]
- Bartlett, B. The Afterlife of a Tree; WWF Poland: Warszawa-Hajnówka, Poland, 2005; ISBN 83-920712-1-2. [Google Scholar]
- Oettel, J.; Zolles, A.; Gschwantner, T.; Lapin, K.; Kindermann, G.; Schweinzer, K.-M.; Gossner, M.M.; Essl, F. Dynamics of standing deadwood in Austrian forests under varying forest management and climatic conditions. J. Appl. Ecol. 2023, 60, 696–713. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Green, M.B.; Fraver, S.; Lutz, D.A.; Woodall, C.W.; D’Amato, A.W.; Evans, D.M. Does Deadwood Moisture Vary Jointly with Surface Soil Water Content? Soil Sci. Soc. Am. J. 2022, 86, 1113–1121. [Google Scholar] [CrossRef]
- Park Williams, A.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Units | Log | Snag | |
|---|---|---|---|---|
| moisture | % | 23.2 (13–59) | 14.1 (12–21) | *** |
| diameter | cm | 43.9 (9–110) | 55.3 (21–131 | n.s. |
| length | m | 12 (4–21) | 11.1 (3.6–22.2) | n.s. |
| volume | m3 | 2.7 (0.03–12.7) | 3.3 (0.6–9.1) | n.s. |
| canopy | % | 8.3 (4.3–21.7) | 11.4 (4.7–42.5) | * |
| cavity | pcs | 0.1 (0–3) | 0.9 (0–7) | * |
| fungi | pcs | 3.2 (0–20) | 5.3 (0–78) | n.s. |
| bark loss | % | 54.3 (0–100) | 42.5 (5–100) | n.s. |
| decay class | class | 1st:15; 2nd:6; 3rd:5; 4th:3 | 1st:9; 2nd:14; 3rd:7 | |
| bryophytes | % | 10.9 (0–75) | 0.2 (0–5) | ** |
| Species | Abbreviation | Czechia RL | German RL | Log | Snag | Sum |
|---|---|---|---|---|---|---|
| Hylis olexai | HylsOlex | EN | NT | 30 (17) | 149 (10) | 179 |
| Melasis buprestoides | MelsBupr | 24 (13) | 87 (23) | 111 | ||
| Hylis foveicollis | HylsFove | EN | NT | 25 (13) | 0 | 25 |
| Hylis cariniceps | HylsCarn | CR | EN | 9 (5) | 2 (2) | 11 |
| Isorhipis melasoides | IsorMels | EN | EN | 8 (3) | 3 (2) | 11 |
| Eucnemis capucina | EucnCapc | EN | EN | 0 | 5 (5) | 5 |
| Clypeorhagus clypeatus | ClypClyp | * | - | 3 (2) | 1 (1) | 4 |
| Microrhagus lepidus | MicrLepd | EN | 2 (2) | 0 | 2 |
| Logs | ||||
|---|---|---|---|---|
| Stat | p-Value | Stat | p-Value | |
| incidence | abundance | |||
| Hylis foveicollis | 0.537 | 0.0001 *** | 0.438 | 0.0001 *** |
| Hylis olexai | 0.254 | 0.0719 | ||
| Microrhagus lepidus | 0.189 | 0.2387 | 0.189 | 0.2340 |
| Hylis cariniceps | 0.163 | 0.2559 | 0.214 | 0.1201 |
| Clypeorhagus clypeatus | 0.081 | 0.6115 | 0.112 | 0.4847 |
| Isorhipis melasoides | 0.066 | 0.6695 | 0.105 | 0.5720 |
| Snags | ||||
| Melasis buprestoides | 0.326 | 0.0189 * | 0.439 | 0.0004 *** |
| Eucnemis capucina | 0.302 | 0.0539 | 0.302 | 0.0512 |
| Hylis olexai | 0.125 | 0.6061 | ||
| Moisture | Diameter | Length | Volume | Canopy | Cavity | Fungi | Bark Loss | Decay Class | Bryophytes | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Clypeorhagus clypeatus | Abundance | +0.008 | +0.054 | ||||||||
| Incidence | +0.068 | ||||||||||
| Eucnemis capucina | Abundance | −0.004 | |||||||||
| Incidence | +0.062 | −0.018 | |||||||||
| Hylis cariniceps | Abundance | ||||||||||
| Incidence | |||||||||||
| Hylis foveicollis | Abundance | −0.047 | −0.058 | −0.022 | |||||||
| Incidence | +0.022 | −0.020 | −0.046 | −0.036 | −0.016 | +0.018 | |||||
| Hylis olexai | Abundance | +0.001 | |||||||||
| Incidence | −0.008 | ||||||||||
| Isorhipis melasoides | Abundance | ||||||||||
| Incidence | |||||||||||
| Melasis buprestoides | Abundance | −0.002 | −0.025 | ||||||||
| Incidence | −0.064 | −0.013 | −0.035 | −0.002 | |||||||
| Microrhagus lepidus | Abundance | +0.002 | |||||||||
| Incidence | +0.023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zumr, V.; Nakládal, O.; Remeš, J. Deadwood-Dwelling Beetles (Coleoptera: Eucnemidae) in a Beech Reserve: A Case Study from the Czech Republic. Forests 2024, 15, 469. https://doi.org/10.3390/f15030469
Zumr V, Nakládal O, Remeš J. Deadwood-Dwelling Beetles (Coleoptera: Eucnemidae) in a Beech Reserve: A Case Study from the Czech Republic. Forests. 2024; 15(3):469. https://doi.org/10.3390/f15030469
Chicago/Turabian StyleZumr, Václav, Oto Nakládal, and Jiří Remeš. 2024. "Deadwood-Dwelling Beetles (Coleoptera: Eucnemidae) in a Beech Reserve: A Case Study from the Czech Republic" Forests 15, no. 3: 469. https://doi.org/10.3390/f15030469