
Genome-Wide Identification of the Argonaute Protein Family and Its Expression Analysis under PEG6000, ABA and Heat Treatments in Populus alba × P. glandulosa
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of AGO genes in Poplar
2.2. Phylogenetic and Characteristic Analysis of the AGO gene Family
2.3. Conserved Motif, Exon, Intron and Chromosome Location of AGO Members in Poplar
2.4. Cis-Regulatory Element Prediction of AGO genes in Poplar
2.5. Plant Materials and Treatments
2.6. RNA Isolation and qRT-PCR
3. Results
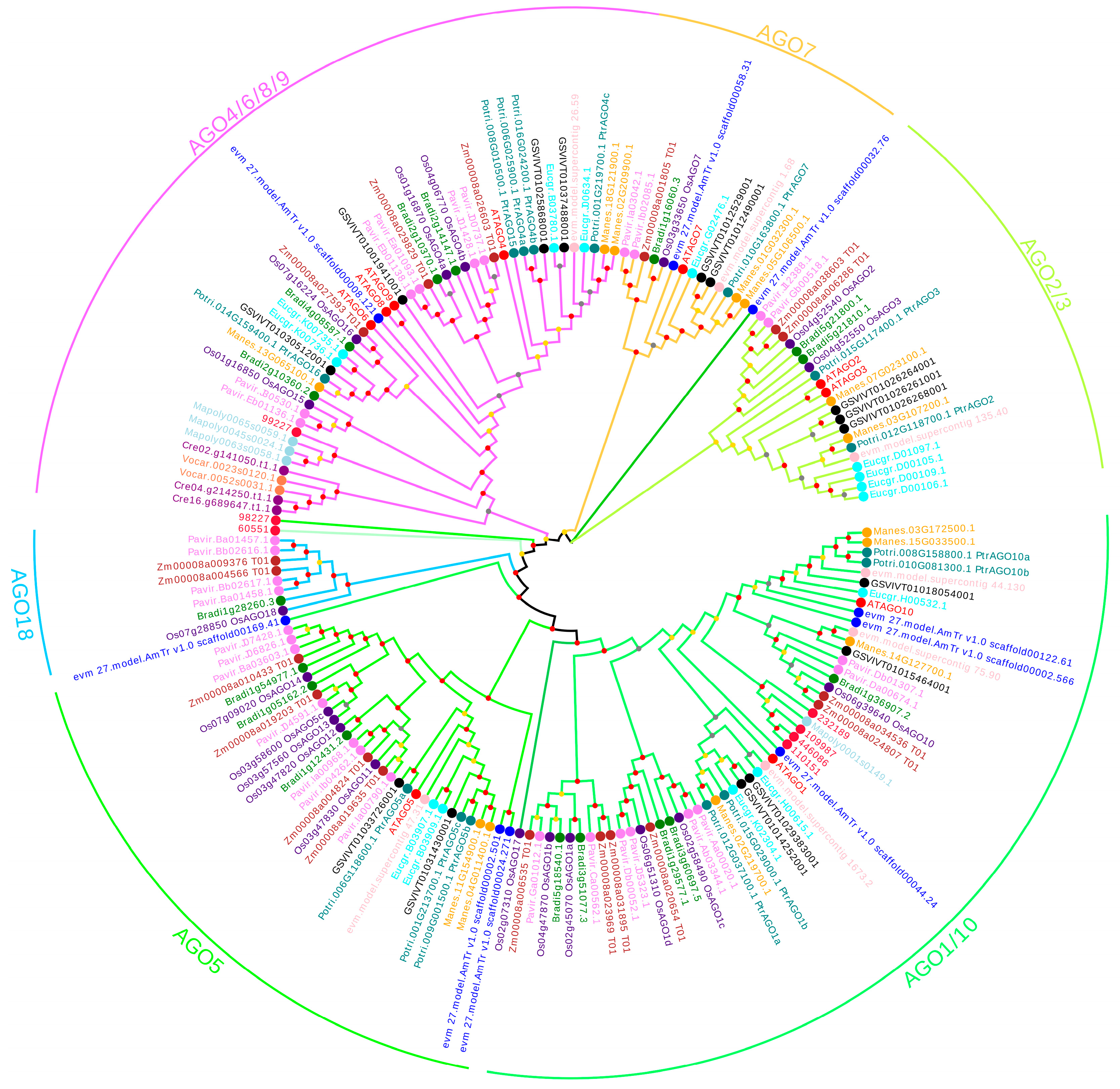
3.1. Identification and Evolution Analysis of AGO genes in Poplar
3.2. Characteristic Analysis of PtAGOs
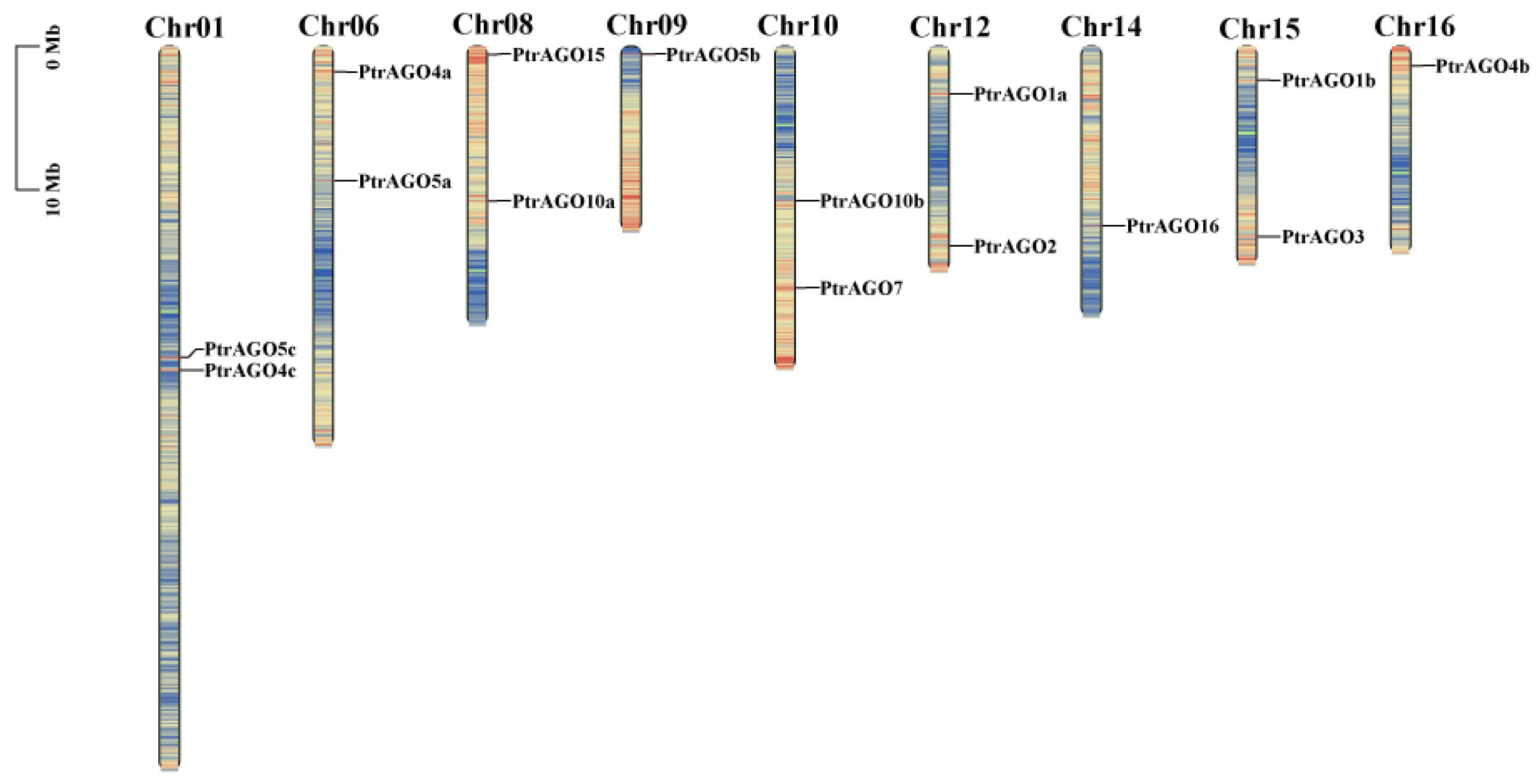
3.3. Location of the PtAGOs in Chromosomes
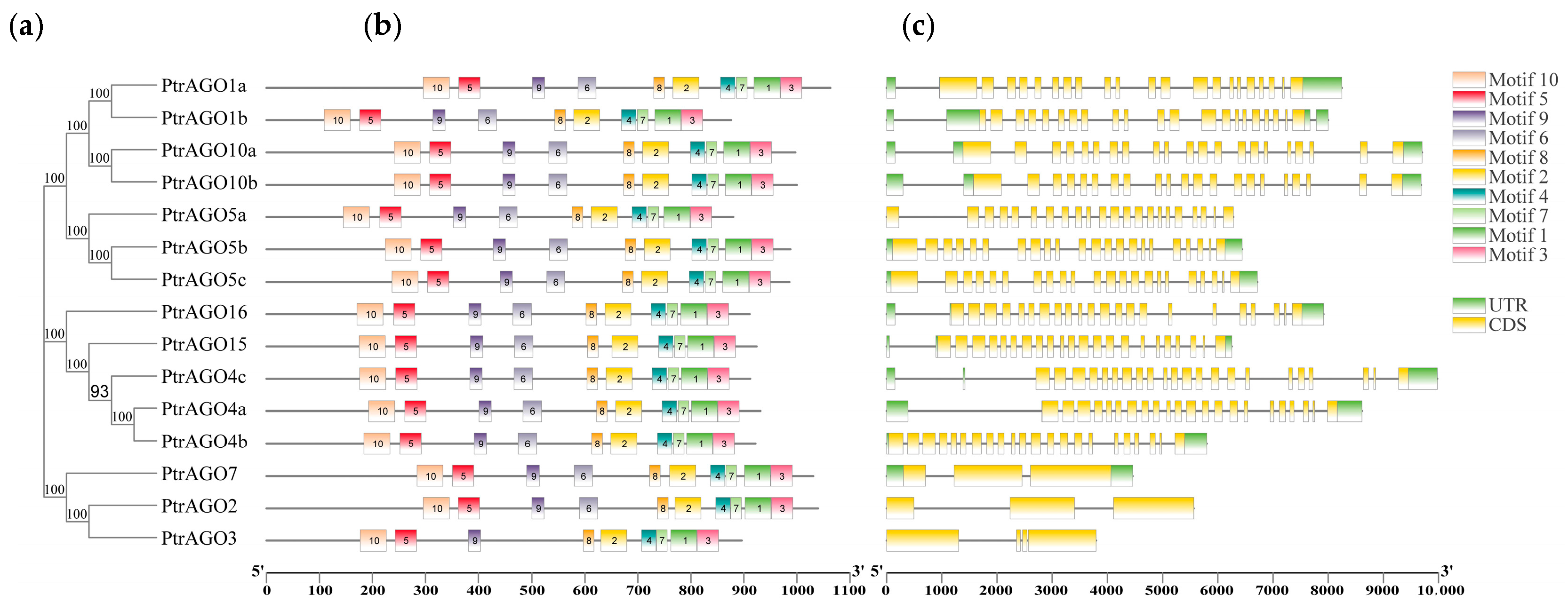
3.4. Conserved Motifs and Gene Structure Analysis of the PtAGO Family in Poplar
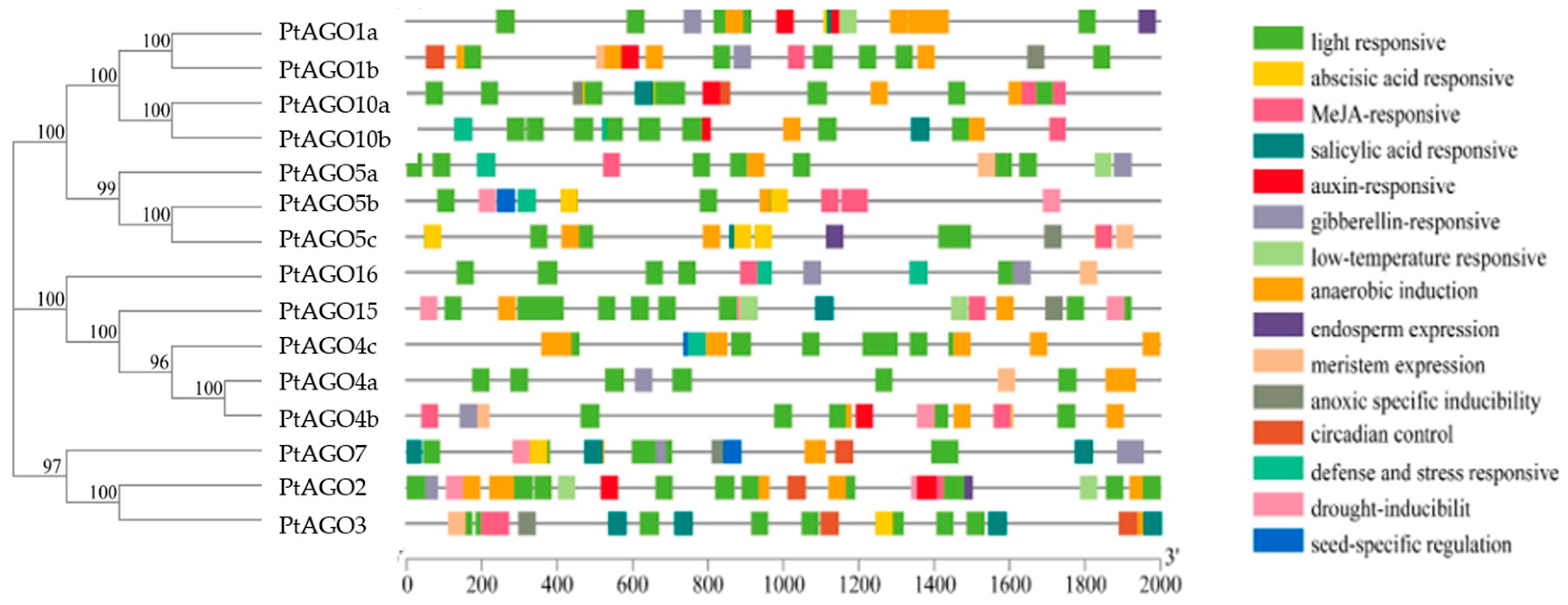
3.5. Cis-Regulatory Element Analysis of PtAGO Promoters
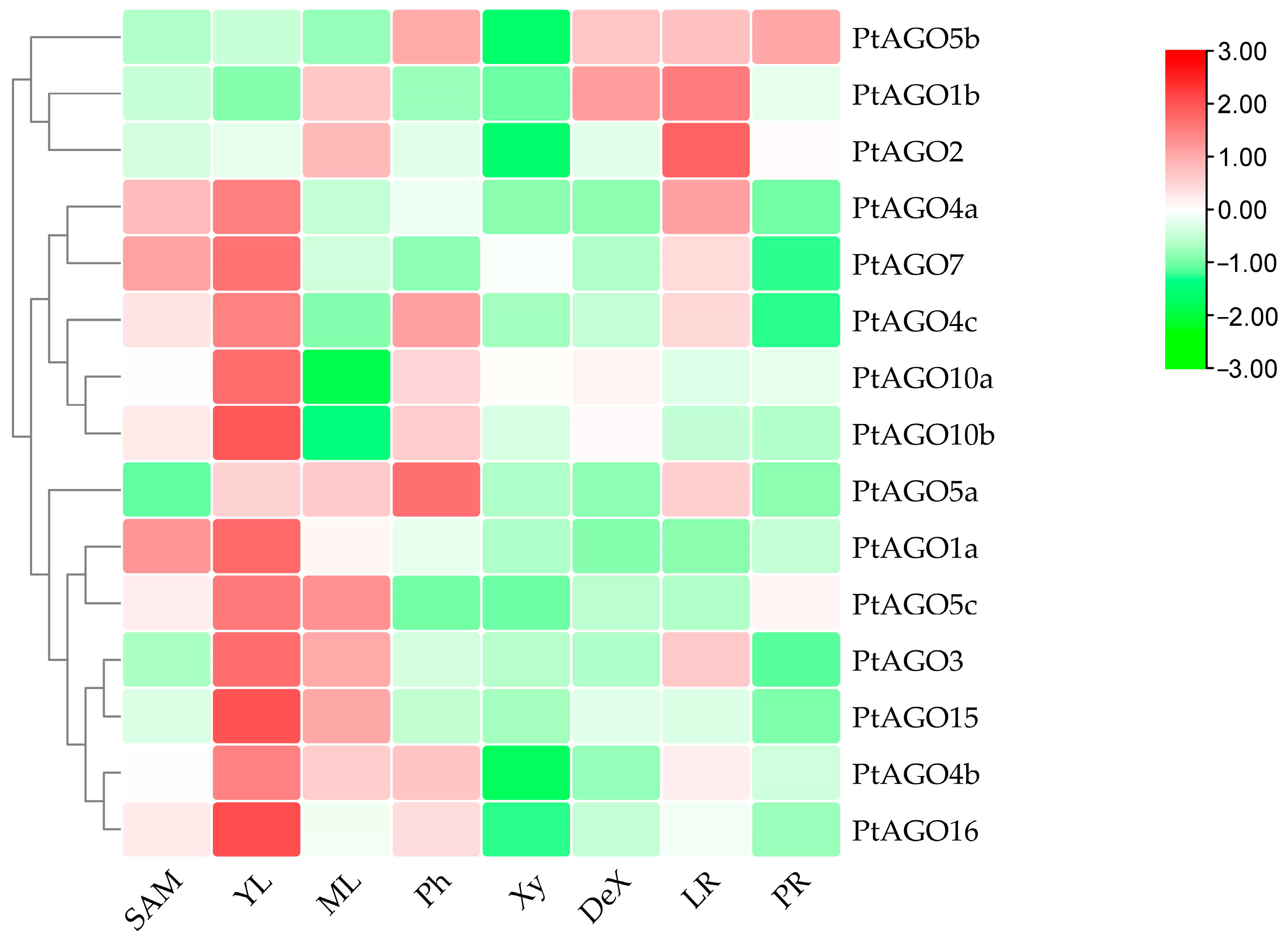
3.6. Expression Pattern of PtAGOs in Various Tissues
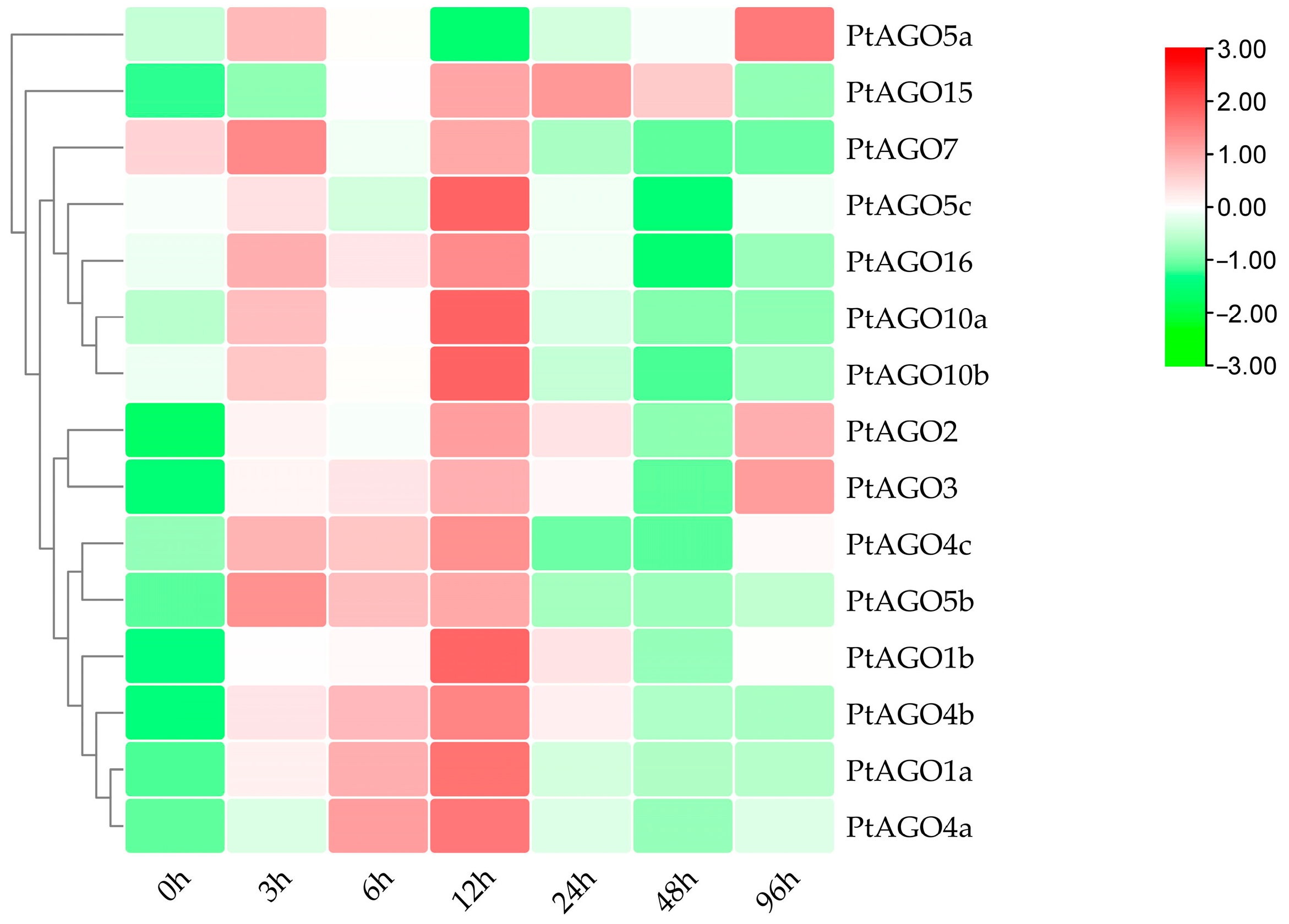
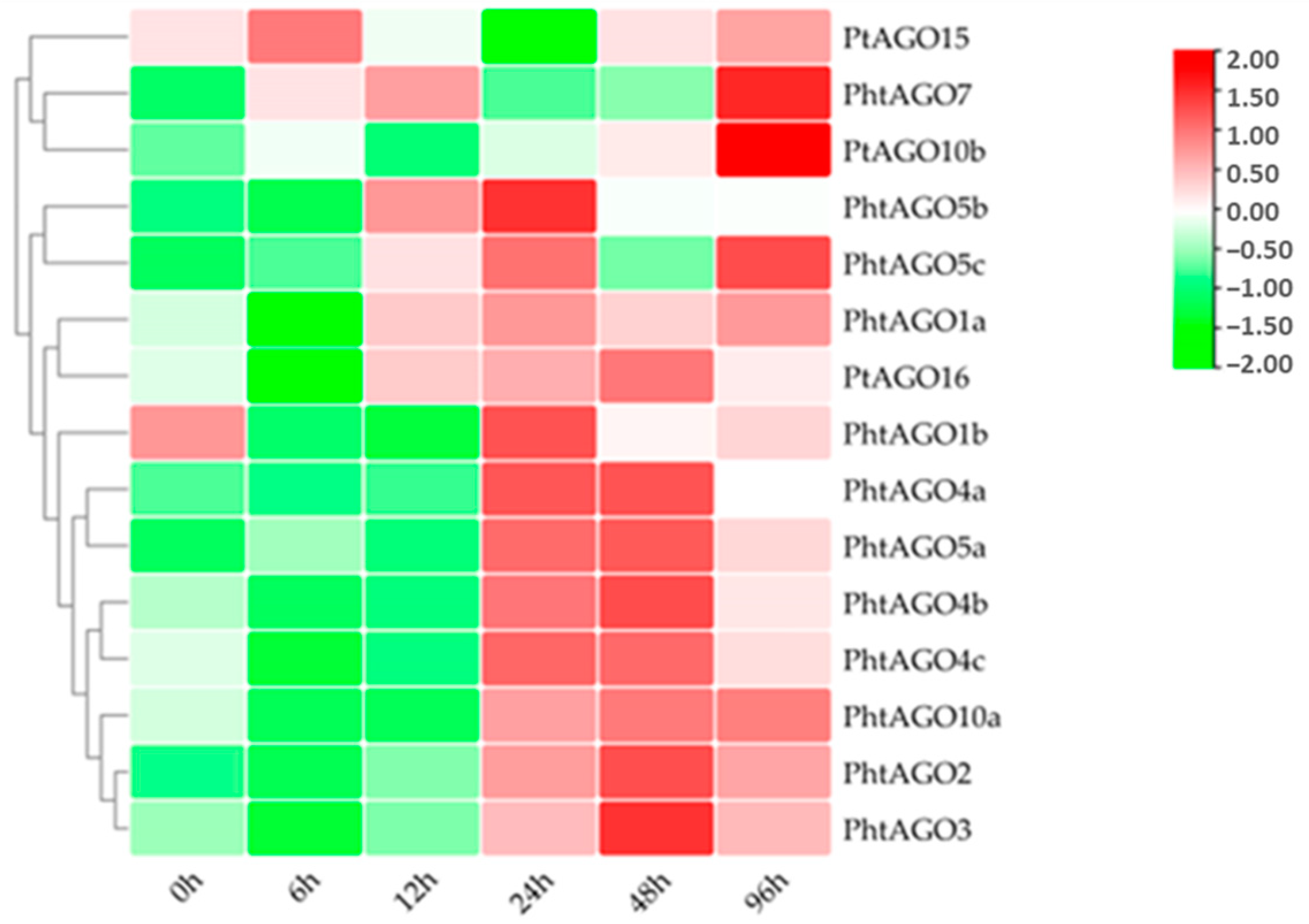
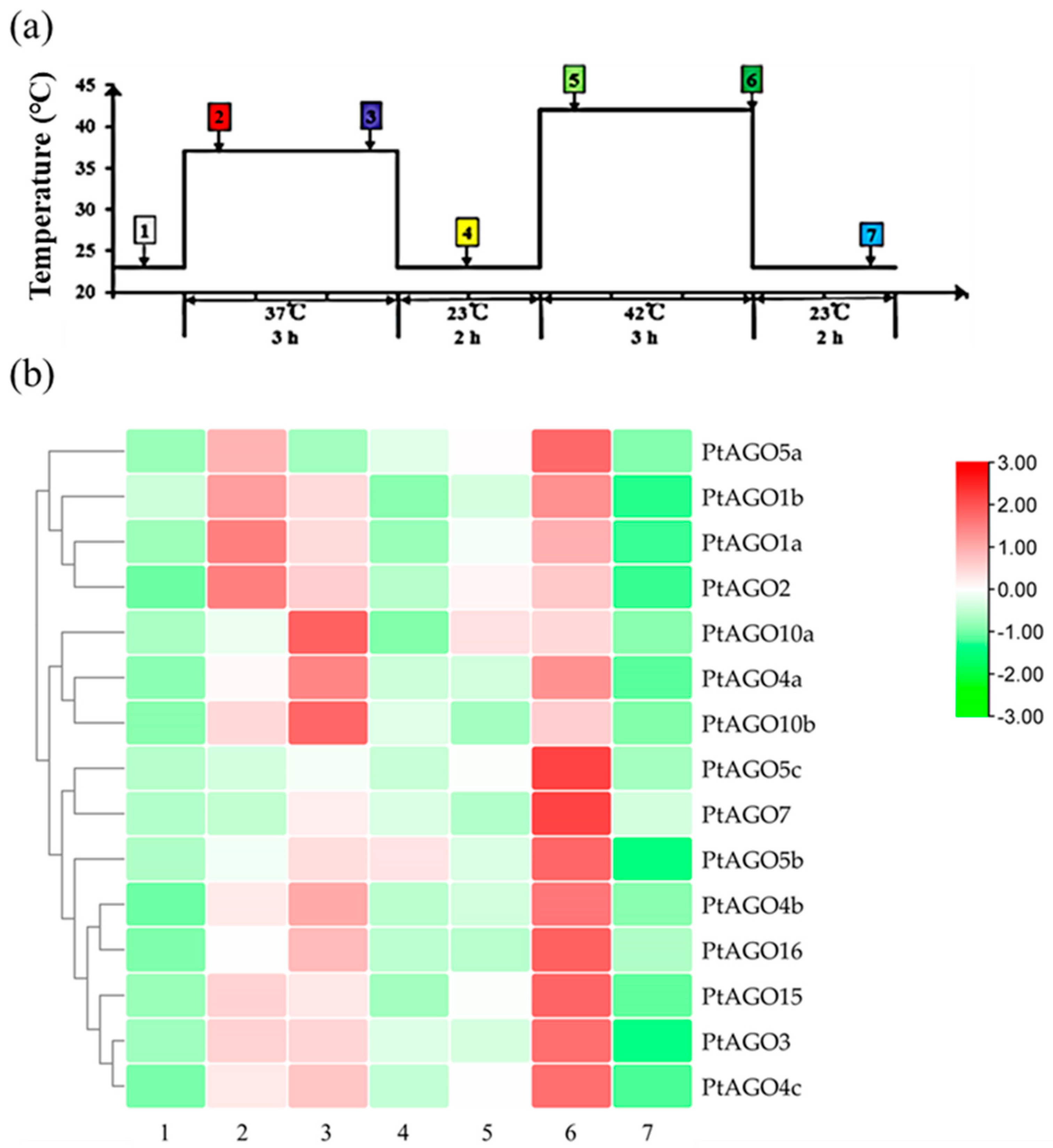
3.7. Expression Pattern of PtAGOs in Response to PEG6000, ABA and Heat Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Bohmert, K.; Camus, I.; Bellini, C.; Bouchez, D.; Caboche, M.; Benning, C. AGO1 defines a novel locus of Arabidopsis controlling leaf development. Embo J. 2014, 17, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Tolia, N.H.; Joshua-Tor, L. Slicer and the argonautes. Nat. Chem. Biol. 2007, 3, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Mallory, A.; Vaucheret, H. Form, function, and regulation of ARGONAUTE proteins. Plant Cell 2010, 22, 3879–3889. [Google Scholar] [CrossRef]
- Hutvagner, G.; Simard, M.J. Argonaute proteins: Key players in RNA silencing. Nat. Rev. Mol. Cell Biol. 2008, 9, 22–32. [Google Scholar] [CrossRef]
- Frank, F.; Hauver, J.; Sonenberg, N.; Nagar, B. Arabidopsis Argonaute MID domains use their nucleotide specificity loop to sort small RNAs. EMBO J. 2012, 31, 3588–3595. [Google Scholar] [CrossRef]
- Yuan, Y.; Pei, Y.; Ma, J.; Kuryavyi, V.; Zhadina, M.; Meister, G.; Chen, H.; Dauter, Z.; Tuschl, T.; Patel, D.J. Crystal structure of A. aeolicus argonaute, a site-specific DNA-guided endoribonuclease, provides insights into RISC-mediated mRNA cleavage. Mol. Cell 2005, 19, 405–419. [Google Scholar] [CrossRef]
- Kapoor, M.; Arora, R.; Lama, T.; Nijhawan, A.; Khurana, J.P.; Tyagi, A.K.; Kapoor, S. Genome-wide identification, organization and phylogenetic analysis of Dicer-like, Argonaute and RNA-dependent RNA Polymerase gene families and their expression analysis during reproductive development and stress in rice. BMC Genom. 2008, 9, 451. [Google Scholar] [CrossRef]
- Bai, M.; Yang, G.; Chen, W.; Mao, Z.; Kang, H.; Chen, G.; Yang, Y.; Xie, B. Genome-wide identification of Dicer-like, Argonaute and RNA-dependent RNA polymerase gene families and their expression analyses in response to viral infection and abiotic stresses in Solanum lycopersicum. Gene 2012, 501, 52–62. [Google Scholar] [CrossRef]
- Liu, X.; Lu, T.; Dou, Y.; Yu, B.; Zhang, C. Identification of RNA silencing components in soybean and sorghum. BMC Bioinform. 2014, 15, 4. [Google Scholar] [CrossRef]
- Qin, L.; Mo, N.; Muhammad, T.; Liang, Y. Genome-wide analysis of DCL, AGO, and RDR gene families in pepper (Capsicum Annuum L.). Int. J. Mol. Sci. 2018, 19, 1038. [Google Scholar] [CrossRef] [PubMed]
- Morel, J.-B.; Godon, C.; Mourrain, P.; Béclin, C.; Boutet, S.; Feuerbach, F.; Proux, F.; Vaucheret, H. Fertile hypomorphic ARGONAUTE (AGO1) mutants impaired in post-transcriptional gene silencing and virus resistance. Plant Cell 2002, 14, 629–639. [Google Scholar] [CrossRef]
- Wang, M.; Qi, Z.; Pei, W.; Cheng, Y.; Mao, K.; Ma, F. The apple Argonaute gene MdAGO1 modulates salt tolerance. Environ. Exp. Bot. 2023, 207, 105202. [Google Scholar] [CrossRef]
- Xu, M.; Xie, W.; Pan, H.; Su, X.; Zhang, S.; Huang, M. Cloning and Characterization of ARGONAUTE Genes in Populus. Sci. Silvae Sin. 2011, 47, 46–51. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhang, J.; Li, J.; Liu, B.; Zhang, L.; Chen, J.; Lu, M. Genome-wide analysis of the Populus Hsp90 gene family reveals differential expression patterns, localization, and heat stress responses. BMC Genom. 2013, 14, 532. [Google Scholar] [CrossRef]
- Zhu, J. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, H.-O.; Tomari, Y. Molecular insights into microRNA-mediated translational repression in plants. Mol. Cell 2013, 52, 591–601. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, W.; Yan, D.; Du, B.; Zhang, J.; Liu, Q.; Chen, S. Characterization of a DRE-binding transcription factor from a halophyte Atriplex hortensis. Theor. Appl. Genet. 2003, 107, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H. Plant argonautes. Trends Plant Sci. 2008, 13, 350–358. [Google Scholar] [CrossRef]
- Dalmadi, Á.; Miloro, F.; Bálint, J.; Várallyay, É.; Havelda, Z. Controlled RISC loading efficiency of miR168 defined by miRNA duplex structure adjusts ARGONAUTE1 homeostasis. Nucleic Acids Res. 2021, 49, 12912–12928. [Google Scholar] [CrossRef] [PubMed]
- Sarkar Das, S.; Majee, M.; Nandi, A.K.; Karmakar, P. Sequestering miR165/166 enhances seed germination in Arabidopsis thaliana under normal condition and ABA treatment. J. Plant Biochem. Biotechnol. 2020, 29, 838–841. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Evolutionary Stage | Species | Group I (AtAGO1/5/10) | Group II (AtAGO2/3/7) | Group III (AtAGO4/6/8/9) | Number |
|---|---|---|---|---|---|
| Algae | Volvox carteri | 0 | 0 | 2 | 2 |
| Chlamydomonas reinhardtii | 0 | 0 | 3 | 3 | |
| Bryophyte | Marchantia polymorpha | 1 | 0 | 3 | 4 |
| Fern | Selaginella moellendorffii | 4 | 0 | 3 | 7 |
| Early angiosperm | Amborella trichopoda | 6 | 2 | 1 | 9 |
| Dicotyledon | Arabidopsis thaliana (L.) Heynh. | 3 | 3 | 4 | 10 |
| Manihot esculenta Crantz | 6 | 4 | 3 | 13 | |
| Vitis vinifera L. | 4 | 4 | 5 | 13 | |
| Eucalyptus grandis Hill | 5 | 5 | 4 | 14 | |
| Populus trichocarpa | 8 | 3 | 4 | 15 | |
| Monocotyledon | Brachypodium distachyon (L.) Beauv. | 10 | 3 | 3 | 16 |
| Zea mays L. | 12 | 3 | 3 | 18 | |
| Oryza sativa L. | 12 | 3 | 4 | 19 |
| Gene Symbol | Gene Identifier | Chr. Location | Start | End | No. of Exons | No. of Introns | Protein Length (a.a.) | CDS Length (bp) | Amino Acids MW (Da) | Isoelectric Point (pI) | Number of Amino Acids | Protein Stability | Cellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PtAGO1a | Potri.012G037100.1 | Chr.12 | 3,306,907 | 3,315,154 | 22 | 21 | 1063 | 3189 | 117,666.6 | 9.41 | 1062 | stable | Nucleus |
| PtAGO1b | Potri.015G029000.1 | Chr.15 | 2,345,768 | 2,353,767 | 23 | 22 | 876 | 2628 | 98,739.23 | 9.31 | 875 | unstable | Nucleus |
| PtAGO2 | Potri.012G118700.1 | Chr.12 | 13,901,935 | 13,907,501 | 3 | 2 | 1040 | 3120 | 113,808.1 | 9.29 | 1039 | stable | Cell membrane/Chloroplast/Nucleus |
| PtAGO3 | Potri.015G117400.1 | Chr.15 | 13,254,772 | 13,258,567 | 4 | 3 | 896 | 2688 | 101,169.4 | 9.21 | 895 | stable | Chloroplast |
| PtAGO4a | Potri.006G025900.1 | Chr.06 | 1,764,851 | 1,773,462 | 23 | 22 | 931 | 2793 | 103,746.4 | 8.91 | 930 | unstable | Cell membrane/Nucleus |
| PtAGO4b | Potri.016G024200.1 | Chr.16 | 1,357,308 | 1,363,107 | 22 | 21 | 922 | 2766 | 102,818.4 | 9.03 | 921 | unstable | Chloroplast |
| PtAGO4c | Potri.001G219700.1 | Chr.01 | 22,549,541 | 22,559,519 | 24 | 23 | 912 | 2736 | 101,826.6 | 8.97 | 911 | unstable | Chloroplast |
| PtAGO5a | Potri.006G118600.1 | Chr.06 | 9,371,285 | 9,377,566 | 22 | 21 | 880 | 2640 | 98,054.72 | 9.31 | 879 | unstable | Nucleus |
| PtAGO5b | Potri.009G001500.1 | Chr.09 | 558,579 | 565,021 | 22 | 21 | 988 | 2964 | 110,387.1 | 9.46 | 987 | unstable | Cell membrane/Nucleus |
| PtAGO5c | Potri.001G213700.1 | Chr.01 | 21,655,182 | 21,661,898 | 22 | 21 | 986 | 2958 | 110,132.4 | 9.47 | 985 | unstable | Cell membrane/Nucleus |
| PtAGO7 | Potri.010G163800.1 | Chr.10 | 16,820,282 | 16,824,742 | 3 | 2 | 1031 | 3093 | 117,405.8 | 9.4 | 1030 | unstable | Cell membrane/Nucleus |
| PtAGO10a | Potri.008G158800.1 | Chr.08 | 10,768,378 | 10,778,079 | 22 | 21 | 997 | 2991 | 111,732 | 9.23 | 996 | unstable | Nucleus |
| PtAGO10b | Potri.010G081300.1 | Chr.10 | 10,737,045 | 10,746,728 | 22 | 21 | 1000 | 3000 | 111,938.4 | 9.33 | 999 | unstable | Cell membrane/Chloroplast |
| PtAGO15 | Potri.008G010500.1 | Chr.08 | 569,178 | 575,431 | 23 | 22 | 924 | 2772 | 103,480.1 | 9.02 | 923 | unstable | Cell membrane/Chloroplast/Nucleus |
| PtAGO16 | Potri.014G159400.1 | Chr.14 | 12,487,183 | 12,495,100 | 23 | 22 | 911 | 2733 | 101,656.6 | 9.18 | 910 | unstable | Chloroplast/Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Wang, Z.; Gao, Y.; Chen, M.; Hao, Z.; Chen, X.; Guo, L.; Fan, C.; Zhao, S. Genome-Wide Identification of the Argonaute Protein Family and Its Expression Analysis under PEG6000, ABA and Heat Treatments in Populus alba × P. glandulosa. Forests 2023, 14, 1015. https://doi.org/10.3390/f14051015
Li H, Wang Z, Gao Y, Chen M, Hao Z, Chen X, Guo L, Fan C, Zhao S. Genome-Wide Identification of the Argonaute Protein Family and Its Expression Analysis under PEG6000, ABA and Heat Treatments in Populus alba × P. glandulosa. Forests. 2023; 14(5):1015. https://doi.org/10.3390/f14051015
Chicago/Turabian StyleLi, Hongying, Zhengbo Wang, Yanwu Gao, Mengdi Chen, Ziyuan Hao, Xi Chen, Lili Guo, Chunjie Fan, and Shutang Zhao. 2023. "Genome-Wide Identification of the Argonaute Protein Family and Its Expression Analysis under PEG6000, ABA and Heat Treatments in Populus alba × P. glandulosa" Forests 14, no. 5: 1015. https://doi.org/10.3390/f14051015