
Molecular Ecological Network Structure and Potential Function of the Bacterial Community in the Soil Profile under Indigenous Tree Plantations in Subtropical China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Study Site and Collection of Soil Samples
2.2. Soil Physicochemical Characteristic Measurement
2.3. Isolation of Soil Bacterial DNA, PCR Amplification, and High-Throughput Sequencing (HTS)
2.4. Bioinformatic Analysis
2.5. Statistical Analysis
3. Results
3.1. Alterations of Soil Physicochemical Character in Native Tree Plantations
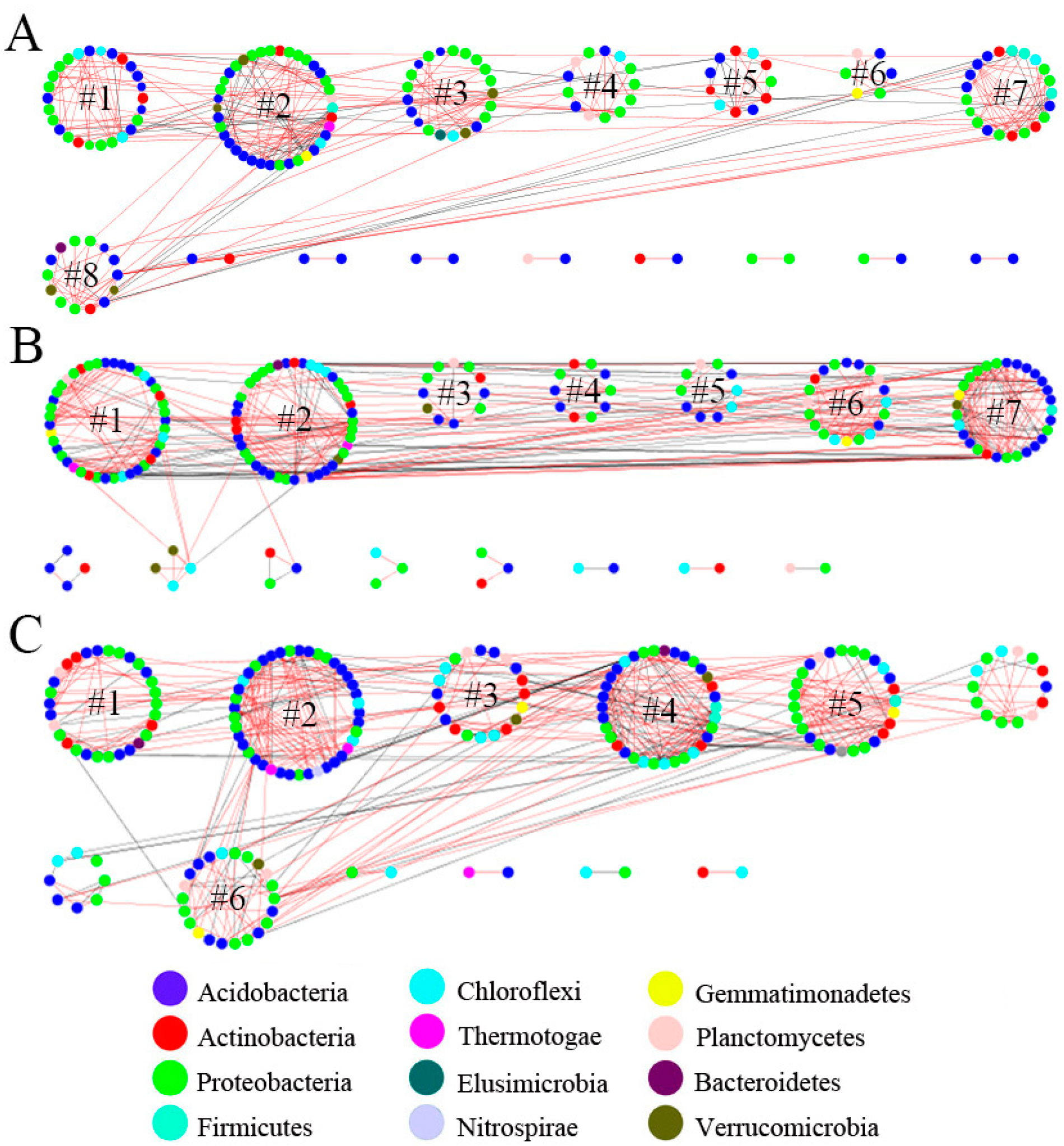
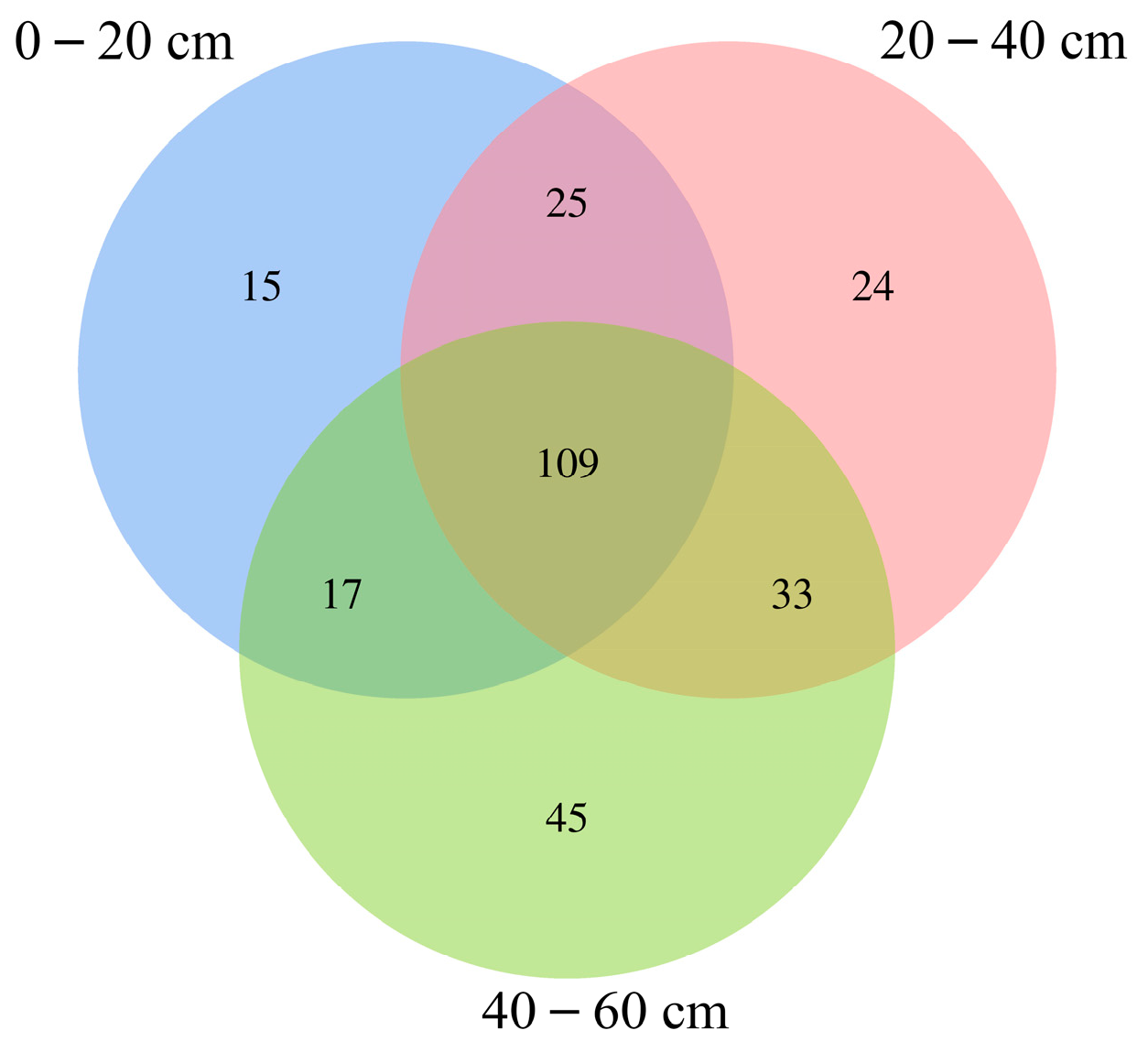
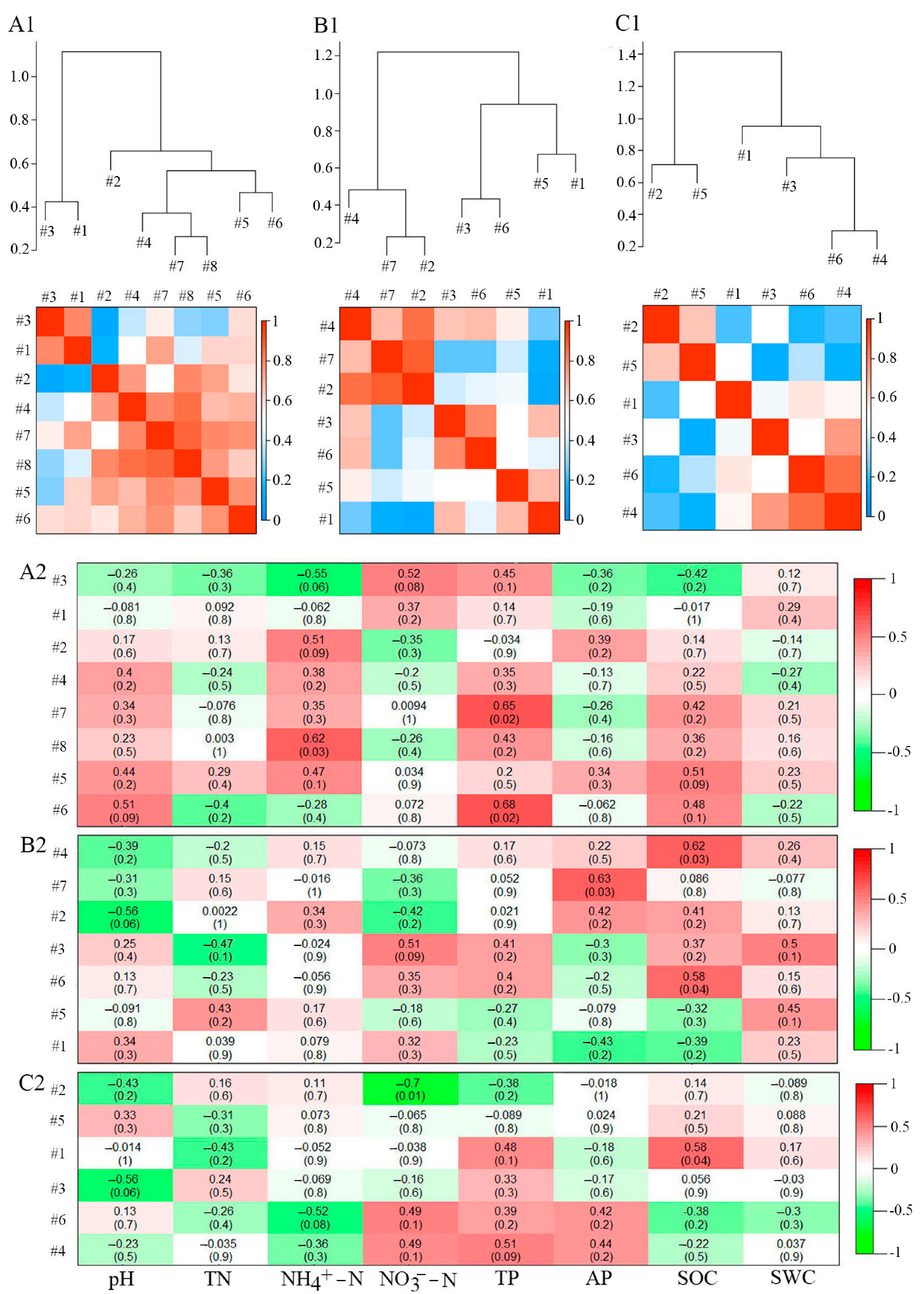
3.2. Characteristics of Bacterial Molecular Ecological Network Structure within Soil Profiles of Native Tree Plantations
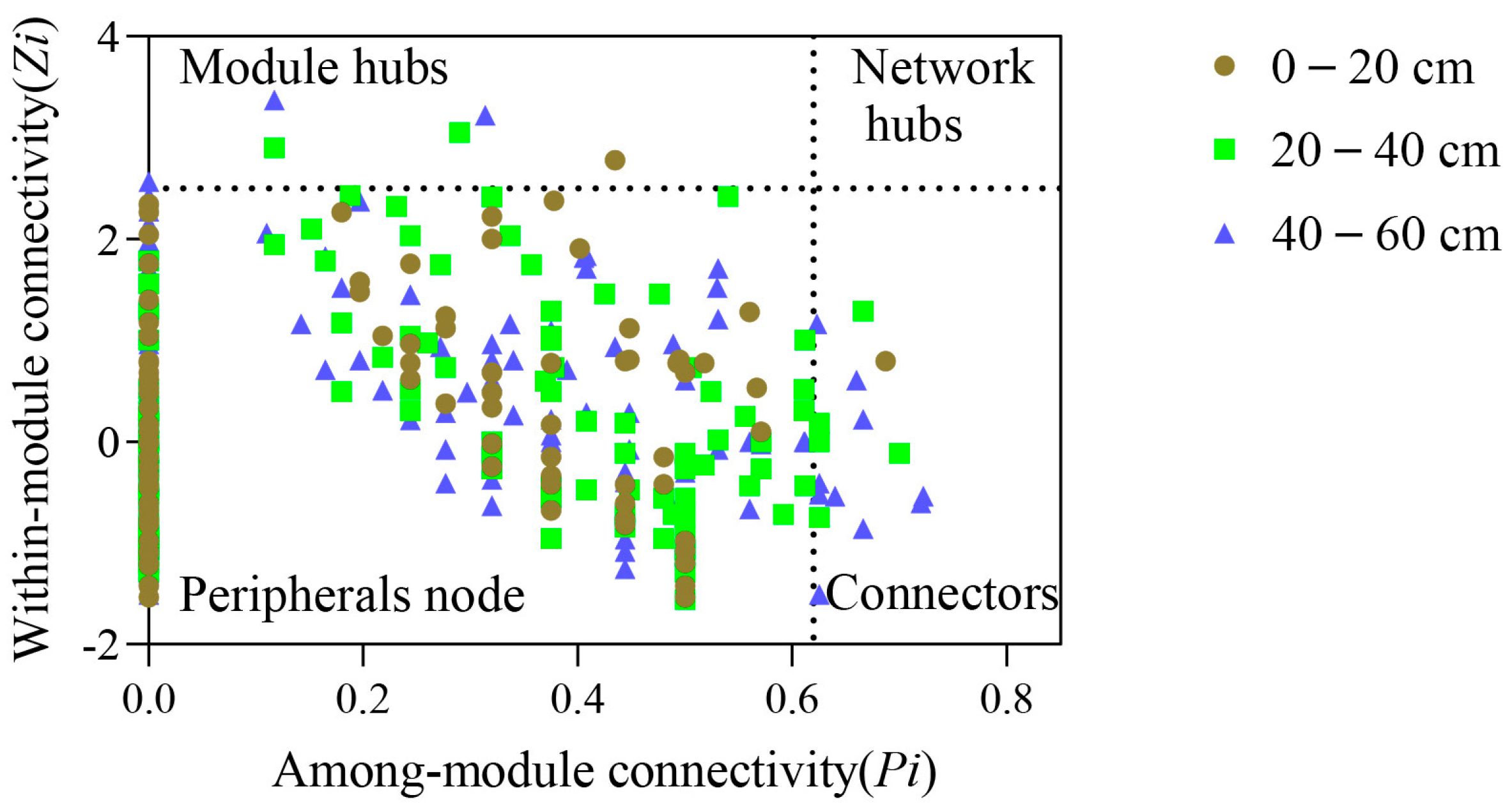
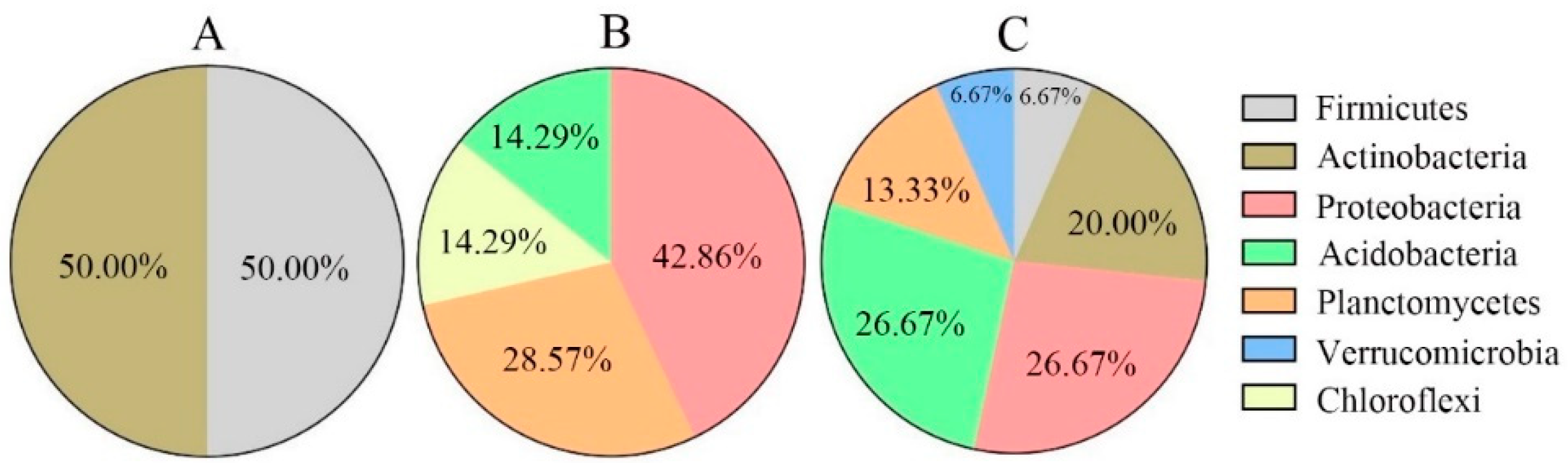
3.3. Topological Functions of Bacterial Molecular Ecological Network Nodes within Soil Profiles of Native Tree Plantations
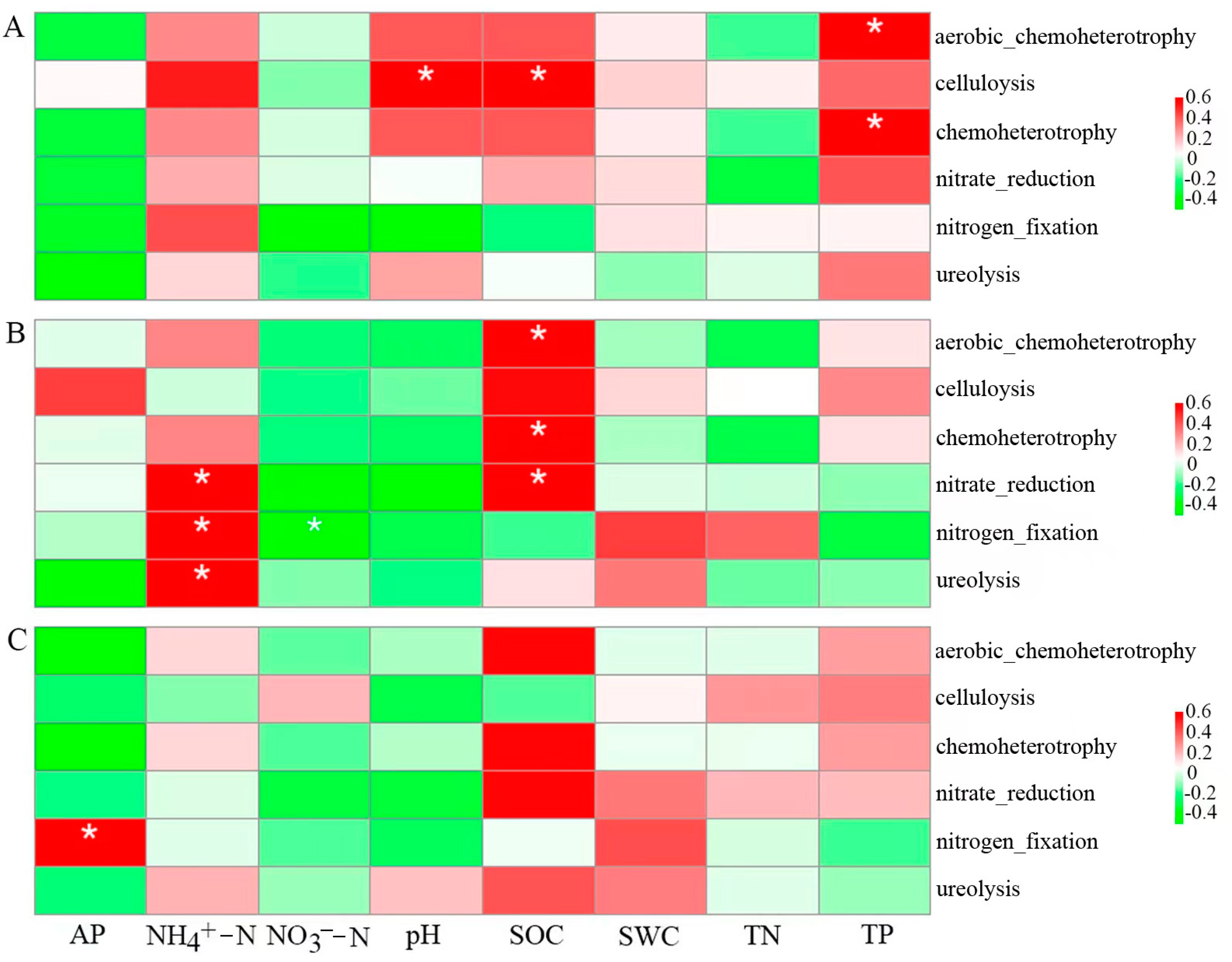
3.4. Functions of Soil Physicochemical Characteristics in Bacterial Molecular Ecological Network Structure
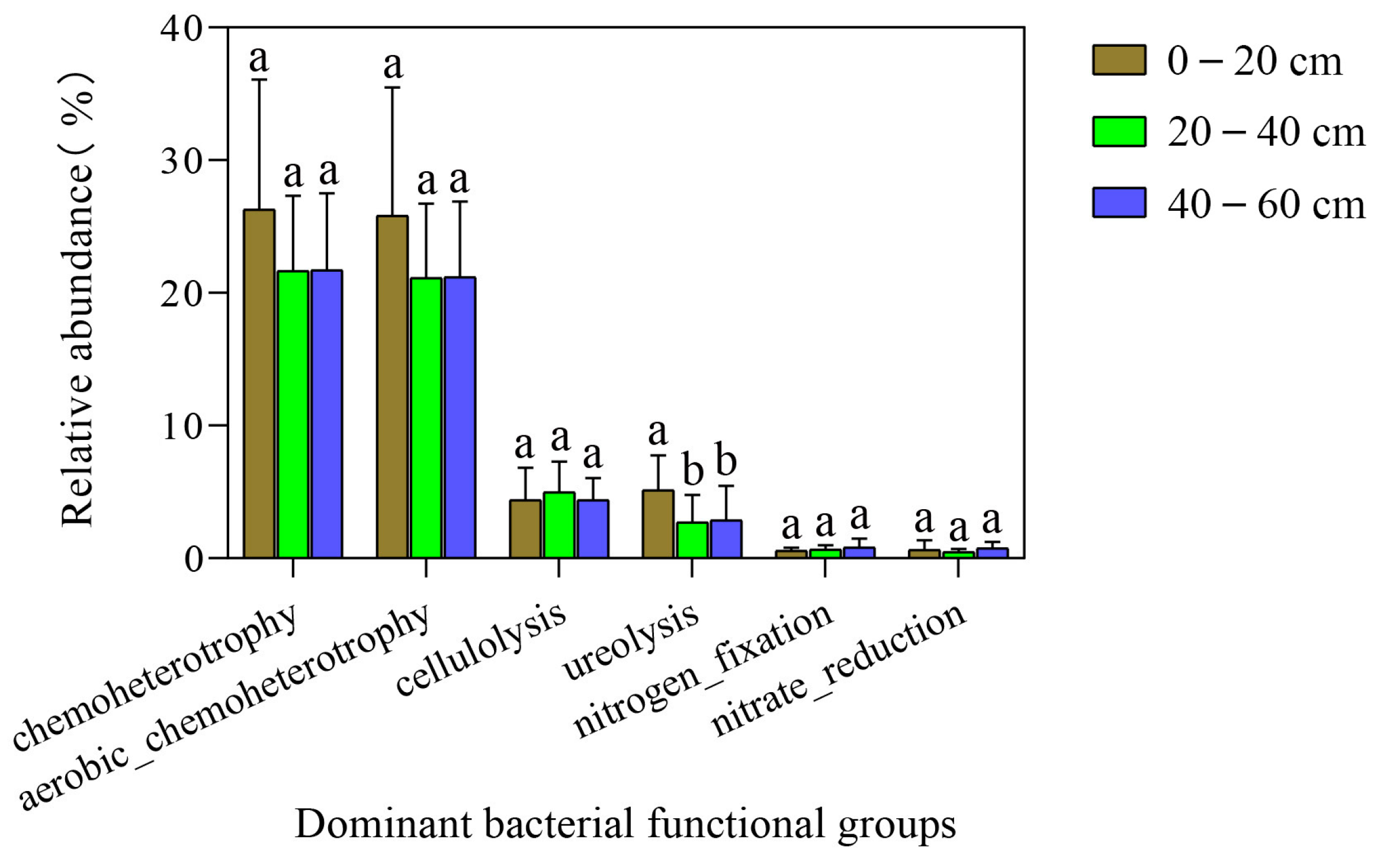
3.5. FAPROTAX Bacterial Functional Estimation within the Soil Profile in Native Tree Plantations
4. Discussion
4.1. Function of Soil Profile Depth in Bacterial Molecular Ecological Network Structure
4.2. Function of Soil Profile Depth in Soil Bacterial Community Function
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Eilers, K.G.; Debenport, S.; Anderson, S.; Fierer, N. Digging deeper to find unique microbial communities: The strong effect of depth on the structure of bacterial and archaeal communities in soil. Soil Biol. Biochem. 2012, 50, 58–65. [Google Scholar] [CrossRef]
- Li, X.; Sun, J.; Wang, H.; Li, X.; Wang, J.; Zhang, H. Changes in the soil microbial phospholipid fatty acid profile with depth in three soil types of paddy fields in China. Geoderma 2017, 290, 69–74. [Google Scholar] [CrossRef]
- Fierer, N.; Lef, J.W.; Adams, B.J.; Nielsen, U.N.; Bates, S.T.; Lauber, C.L.; Owens, S.; Gilbert, J.A.; Wall, D.H.; Caporaso, J.G. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proc. Natl. Acad. Sci. USA 2012, 109, 21390–21395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Yun, Y.; Wang, H.; Ma, L.; Tian, W.; Man, B.; Liu, C. Contrasting bacterial communities and their assembly processes in karst soils under different land use. Sci. Total Environ. 2021, 751, 142263. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Lee, M.J.; Jung, J.Y.; Hwang, C.Y.; Kim, M.; Ro, H.M.; Chun, J.; Lee, Y.K. Vertical distribution of bacterial community is associated with the degree of soil organic matter decomposition in the active layer of moist acidic tundra. J. Microbiol. 2016, 54, 713–723. [Google Scholar] [CrossRef]
- Wang, M.; Tian, J.; Bu, Z.; Lamit, L.J.; Chen, H.; Zhu, Q.; Peng, C. Structural and functional differentiation of the microbial community in the surface and subsurface peat of two minerotrophic fens in China. Plant Soil 2019, 437, 21–40. [Google Scholar] [CrossRef]
- Xu, T.; Chen, X.; Hou, Y.; Zhu, B. Changes in microbial biomass, community composition and diversity, and functioning with soil depth in two alpine ecosystems on the Tibetan plateau. Plant Soil 2020, 459, 137–153. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, L.; Yu, Z.; Peng, N.; Xiao, L.; Yin, D.; Qin, B. Characterization of depth-related microbial communities in lake sediment by denaturing gradient gel electrophoresis of amplified 16S rRNA fragments. J. Environ. Sci. 2008, 20, 224–230. [Google Scholar] [CrossRef]
- Tang, Y.; Yu, G.; Zhang, X.; Wang, Q.; Ge, J.; Liu, S. Changes in nitrogen-cycling microbial communities with depth in temperate and subtropical forest soils. Appl. Soil Ecol. 2018, 124, 218–228. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Y.; Hu, X.; Wan, S.; Wang, H.; Liang, C.; Chen, F. Litter manipulation effects on microbial communities and enzymatic activities vary with soil depth in a subtropical Chinese fir plantation. For. Ecol. Manag. 2021, 480, 118641. [Google Scholar] [CrossRef]
- Montoya, J.M.; Pimm, S.L.; Sole, R.V. Ecological networks and their fragility. Nature 2006, 442, 259–264. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Tu, Q.; Zhi, X. Functional molecular ecological networks. mBio 2010, 1, e00160–e00169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Jiang, Y.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinf. 2012, 13, 113. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Lu, Y.; Chen, F.; Li, X.; Xiao, D.; Wang, H. Molecular Ecological Network Complexity Drives Stand Resilience of Soil Bacteria to Mining Disturbances among Typical Damaged Ecosystems in China. Microorganisms 2020, 8, 433. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Xue, D.; Wang, Y.; Zheng, W.; Zhang, G.; Wang, Z.L. Molecular ecological network analysis of the response of soil microbial communities to depth gradients in farmland soils. Microbiol. Open 2020, 9, e983. [Google Scholar] [CrossRef]
- Liu, S.; Yang, Y.; Wang, H. Development strategy and management countermeasures of planted forests in China: Transforming from timber-centered single objective management towards multi-purpose management for enhancing quality and benefits of ecosystem services. Acta Ecol. Sin. 2018, 38, 1–10. [Google Scholar]
- State Forestry and Grassland Administration. China Forest Resources Rreport (2014–2018); China Forestry Press: Beijing, China, 2020.
- Wan, X.; Huang, Z.; He, Z.; Yu, Z.; Wang, M.; David, R.M.; Yang, Y. Soil C: N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations. Plant Soil 2015, 387, 103–116. [Google Scholar] [CrossRef]
- You, Y.; Xu, H.; Wu, X.; Zhou, X.; Tan, X.; Li, M.; Wen, Y.; Zhu, H.; Cai, D.; Huang, X. Native broadleaf tree species stimulate topsoil nutrient transformation by changing microbial community composition and physiological function, but not biomass in subtropical plantations with low P status. For. Ecol. Manag. 2020, 477, 118491. [Google Scholar] [CrossRef]
- He, Y.; Qin, L.; Li, Z.; Liang, X.; Shao, M.; Tan, L. Carbon storage capacity of monoculture and mixed-species plantations in subtropical China. For. Ecol. Manag. 2013, 295, 193–198. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.R.; Mo, J.M.; Wang, J.X.; Makeschin, F.; Wolff, M. Soil organic carbon stock and chemical composition in four plantations of indigenous tree species in subtropical China. Ecol. Res. 2010, 25, 1071–1079. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Wang, J.; Shi, Z.; Lu, L.; Zeng, J.; Ming, A.; Tang, J.; Yu, H. Effects of tree species mixture on soil organic carbon stocks and greenhouse gas fluxes in subtropical plantations in China. For. Ecol. Manag. 2013, 300, 4–13. [Google Scholar] [CrossRef]
- Lu, R. Methods for Agrochemical Analysis of Soil; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Yang, Y. Phylogenetic molecular ecological network of soil microbial communities in response to elevated CO2. Mbio 2011, 2, e0012211. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Sansupa, C.; Wahdan, S.F.M.; Hossen, S.; Disayathanoowat, T.; Wubet, T.; Purahong, W. Can we use functional annotation of prokaryotic taxa (FAPROTAX) to assign the ecological functions of soil bacteria? Appl. Sci. 2021, 11, 688. [Google Scholar] [CrossRef]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling interactions in the microbiome: A network perspective. Trends Microbiol. 2017, 25, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Mundra, S.; Kjonaas, O.J.; Morgado, L.N.; Krabberod, A.K.; Ransedokken, Y.; Kauserud, H. Soil depth matters: Shift in composition and inter-kingdom co-occurrence patterns of microorganisms in forest soils. FEMS Microbiol. Ecol. 2021, 97, fiab022. [Google Scholar] [CrossRef]
- Morrien, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; de Hollander, M.; Soto, R.L.; Bouffaud, M.L.; Buee, M.; Dimmers, W.; et al. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef] [Green Version]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E.E.; van der Heijden, M.G.A. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nat. Commun. 2019, 10, 4841. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Wang, Y.; Lu, S.; Xiang, Q.; Yu, X.; Zhao, K.; Zou, L.; Chen, Q.; Tu, S.; Zhang, X. Long-term fertilization structures bacterial and archaeal communities along soil depth gradient in a paddy soil. Front. Microbiol. 2017, 8, 1516. [Google Scholar] [CrossRef] [Green Version]
- de Menezes, A.B.; Richardson, A.E.; Thrall, P.H. Linking fungal-bacterial co-occurrences to soil ecosystem function. Curr. Opin. Microbiol. 2017, 37, 135–141. [Google Scholar] [CrossRef]
- Bai, R.; Wang, J.T.; Deng, Y.; He, J.Z.; Feng, K.; Zhang, L.M. Microbial community and functional structure significantly varied among distinct types of paddy soils but responded differently along gradients of soil depth layers. Front. Microbiol. 2017, 8, 945. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Eldridge, D.J.; Hamonts, K.; Reich, P.B.; Singh, B.K. Experimentally testing the species-habitat size relationship on soil bacteria: A proof of concept. Soil Biol. Biochem. 2018, 123, 200–206. [Google Scholar] [CrossRef] [Green Version]
- An, Q.; Xu, M.; Zhang, X.; Jiao, K.; Zhang, C. Soil bacterial community composition and functional potentials along the vertical vegetation transection on Mount Segrila, Tibaet, China. Chin. J. Appl. Ecol. 2021, 32, 2147–2157. [Google Scholar]
- Ahamed Bakeri, S.; Thakib Maidin, M.S.; Mohd Masri, M.M. Soil bacterial biodiversity in development of secondary loggedover forest to oil palm plantation in mineral soil of Belaga, Sarawak. J. Oil Palm Res. 2019, 31, 394–411. [Google Scholar] [CrossRef]
- Jurado, M.; Lopez, M.J.; Suarez-Estrella, F.; Vargas-Garcia, M.C.; Lopez-Gonzalez, J.A.; Moreno, J. Exploiting composting biodiversity: Study of the persistent and biotechnologically relevant microorganisms from lignocellulose-based composting. Bioresour. Technol. 2014, 162, 283–293. [Google Scholar] [CrossRef]
- Kirby, R. Actinomycetes and lignin degradation. Adv. Appl. Microbiol. 2005, 58, 125–168. [Google Scholar]
- Kramer, C.; Gleixner, G. Soil organic matter in soil depth profiles: Distinct carbon preferences of microbial groups during carbon transformation. Soil Biol. Biochem. 2008, 40, 425–433. [Google Scholar] [CrossRef]
- Yang, L.; Barnard, R.; Kuzyakov, Y.; Tian, J. Bacterial communities drive the resistance of soil multifunctionality to land-use change in karst soils. Eur. J. Soil Biol. 2021, 104, 103313. [Google Scholar] [CrossRef]
- Øvreås, L.; Torsvik, V. Microbial diversity and community structure in two different agricultural soil communities. Microb. Ecol. 1998, 36, 303–315. [Google Scholar] [CrossRef]
- Chu, H.; Sun, H.; Tripathi, B.M.; Adams, J.M.; Huang, R.; Zhang, Y.; Shi, Y. Bacterial community dissimilarity between the surface and subsurface soils equals horizontal differences over several kilometers in the western Tibetan Plateau. Environ. Microbiol. 2016, 18, 1523–1533. [Google Scholar] [CrossRef]
- Ma, Y.; Feng, C.; Wang, Z.; Huang, C.; Huang, X.; Wang, W.; Yang, S.; Fu, S.; Chen, H. Restoration in degraded subtropical broadleaved forests induces changes in soil bacterial communities. Glob. Ecol. Conserv. 2021, 30, e01775. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Q.; Ma, J.; Yang, Y.; Wang, Y.; Fu, C. Flooding irrigation weakens the molecular ecological network complexity of soil microbes during the process of dryland-to-paddy conversion. Int. J. Environ. Res. Public Health 2020, 17, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Zhao, M.; Cong, J.; Qi, Q.; Xiao, Y.; Cong, W.; Deng, Y.; Zhou, J.; Zhang, Y. Soil pH exerts stronger impacts than vegetation type and plant diversity on soil bacterial community composition in subtropical broad-leaved forests. Plant Soil 2020, 450, 273–286. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéja, R.; Baldria, P. Forest soil bacteria: Diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [Green Version]
- Almonacid-Muñoz, L.; Herrera, H.; Fuentes-Ramírez, A.; Vargas-Gaete, R.; Larama, G.; Jara, R.; Fernández-Urrutia, C.; da Silva Valadares, R.B. Tree cover species modify the diversity of rhizosphere-associated microorganisms in nothofagus obliqua (Mirb.) oerst temperate forests in south-central Chile. Forests 2022, 13, 756. [Google Scholar] [CrossRef]
- Sun, W.; Li, Z.; Lei, J.; Liu, X. Bacterial communities of forest soils along different elevations: Diversity, structure, and functional composition with potential impacts on CO2 emission. Microorganisms 2022, 10, 766. [Google Scholar] [CrossRef]
- O’Donnell, A.G.; Seasman, M.; Macrae, A.; Waite, I.; Davies, J.T. Plants and fertilisers as drivers of change in microbial community structure and function in soils. Plant Soil 2001, 232, 135–145. [Google Scholar] [CrossRef]
- Liang, S.; Deng, J.; Jiang, Y.; Wu, S.; Zhou, Y.; Zhu, W. Functional distribution of bacterial community under different land use patterns based on FaProTax function prediction. Pol. J. Environ. Stud. 2020, 29, 1245–1261. [Google Scholar] [CrossRef] [PubMed]
- Brackin, R.; Robinson, N.; Lakshmanan, P.; Schmidt, S. Microbial function in adjacent subtropical forest and agricultural soil. Soil Biol. Biochem. 2013, 57, 68–77. [Google Scholar] [CrossRef]
- Martin, J.F.; Reddy, K.R. Interaction and spatial distribution of wetland nitrogen processes. Ecol. Modell. 1997, 105, 1–21. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Layer | SWC (%) | pH | SOC (g/kg) | TN (g/kg) | NH4+-N (mg/kg) | NO3−-N (mg/kg) | TP (g/kg) | AP (mg/kg) |
|---|---|---|---|---|---|---|---|---|
| 0–20 cm | 31.32 ± 2.56 a | 4.17 ± 0.24 a | 21.17 ± 2.11 a | 1.43 ± 0.34 a | 19.38 ± 5.49 a | 4.42 ± 0.63 a | 1.26 ± 0.49 a | 13.85 ± 3.64 a |
| 20–40 cm | 28.02 ± 2.92 b | 4.23 ± 0.31 a | 12.05 ± 1.87 b | 1.10 ± 0.20 b | 15.07 ± 7.09 ab | 3.20 ± 1.06 b | 0.96 ± 0.44 a | 11.76 ± 4.98 a |
| 40–60 cm | 25.86 ± 3.69 b | 4.27 ± 0.25 a | 9.58 ± 1.62 c | 0.74 ± 0.17 c | 10.07 ± 6.37 b | 1.93 ± 0.87 c | 0.43 ± 0.22 b | 9.99 ± 5.27 a |
| Soil Layer | Molecular Ecological Networks | Random Networks | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cutoff | Total Nodes | Total Links | R2 of Power Law | The Number of Positive Connections | The Number of Negative Connections | Average Degree | Average Clustering Coefficient | Average Path Dis-tance | Modularity Index (Module Number) | Average Clustering Coefficient | Average Path Distance | Modularity Index | |
| 0–20 cm | 0.76 | 166 | 333 | 0.752 | 263 | 70 | 4.012 | 0.203 | 4.451 | 0.669 (16) | 0.029 ± 0.009 | 3.665 ± 0.051 | 0.475 ± 0.009 |
| 20–40 cm | 0.76 | 191 | 488 | 0.813 | 345 | 143 | 5.11 | 0.225 | 4.148 | 0.586 (15) | 0.049 ± 0.009 | 3.308 ± 0.043 | 0.400 ± 0.007 |
| 40–60 cm | 0.76 | 204 | 519 | 0.796 | 344 | 175 | 5.088 | 0.235 | 4.204 | 0.623 (12) | 0.042 ± 0.008 | 3.359 ± 0.035 | 0.406 ± 0.007 |
| Bacterial Functional Group | Relative Abundance (%) | ||
|---|---|---|---|
| 0–20 cm | 20–40 cm | 40–60 cm | |
| chemoheterotrophy | 26.3037 | 21.6669 | 21.7211 |
| aerobic chemoheterotrophy | 25.8136 | 21.1417 | 21.2079 |
| cellulolysis | 4.3823 | 4.9789 | 4.3766 |
| ureolysis | 5.1485 | 2.7107 | 2.8567 |
| nitrogen fixation | 0.5692 | 0.6592 | 0.8078 |
| nitrate reduction | 0.6468 | 0.5049 | 0.7700 |
| methanotrophy | 0.0115 | 0.0199 | 0.0178 |
| methanol_oxidation | 0.0191 | 0.0444 | 0.0738 |
| methylotrophy | 0.0306 | 0.0643 | 0.0916 |
| aerobic_ammonia_oxidation | 0.0011 | 0.0010 | 0.0022 |
| aerobic_nitrite_oxidation | 0.0146 | 0.0979 | 0.2692 |
| nitrification | 0.0157 | 0.0989 | 0.2714 |
| sulfur_respiration | 0.0000 | 0.0006 | 0.0015 |
| respiration_of_sulfur_compounds | 0.0000 | 0.0006 | 0.0015 |
| chitinolysis | 0.0004 | 0.0002 | 0.0002 |
| nitrite_ammonification | 0.0938 | 0.2136 | 0.3710 |
| nitrite_respiration | 0.0938 | 0.2136 | 0.3710 |
| dark_thiosulfate_oxidation | 0.0731 | 0.0210 | 0.0135 |
| dark_oxidation_of_sulfur_compounds | 0.0736 | 0.0212 | 0.0135 |
| manganese_oxidation | 0.0086 | 0.0119 | 0.0175 |
| fermentation | 0.2850 | 0.3708 | 0.5907 |
| invertebrate_parasites | 0.0017 | 0.0021 | 0.0038 |
| human_pathogens_pneumonia | 0.0000 | 0.0012 | 0.0013 |
| human_pathogens_gastroenteritis | 0.0938 | 0.2136 | 0.3710 |
| human_pathogens_diarrhea | 0.0938 | 0.2136 | 0.3710 |
| human_pathogens_all | 0.1186 | 0.2504 | 0.4334 |
| human_gut | 0.0946 | 0.2136 | 0.3724 |
| human_associated | 0.1195 | 0.2504 | 0.4349 |
| mammal_gut | 0.0946 | 0.2136 | 0.3724 |
| animal_parasites_or_symbionts | 0.2356 | 0.3761 | 0.5379 |
| plant_pathogen | 0.0570 | 0.0292 | 0.0433 |
| aromatic_hydrocarbon_degradation | 0.0006 | 0.0014 | 0.0021 |
| aromatic_compound_degradation | 0.0336 | 0.0665 | 0.1346 |
| liphatic_non_methane_hydrocarbon_degradation | 0.0006 | 0.0014 | 0.0021 |
| hydrocarbon_degradation | 0.0121 | 0.0214 | 0.0199 |
| iron_respiration | 0.0304 | 0.0455 | 0.0482 |
| nitrate_respiration | 0.1117 | 0.2414 | 0.4375 |
| nitrogen_respiration | 0.1117 | 0.2414 | 0.4375 |
| fumarate_respiration | 0.0938 | 0.2136 | 0.3710 |
| intracellular_parasites | 0.1295 | 0.1084 | 0.0770 |
| predatory_or_exoparasitic | 0.0368 | 0.0438 | 0.0694 |
| chloroplasts | 0.0045 | 0.0103 | 0.0084 |
| nonphotosynthetic_cyanobacteria | 0.0382 | 0.0210 | 0.0239 |
| aerobic_anoxygenic_phototrophy | 0.0010 | 0.0006 | 0.0051 |
| photoheterotrophy | 0.0219 | 0.0287 | 0.0393 |
| phototrophy | 0.0219 | 0.0287 | 0.0393 |
| Total | 65.1426 | 55.6798 | 58.5035 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, L.; Wang, Y.; Ming, A.; Xi, S.; Xiao, Z.; Teng, J.; Tan, L. Molecular Ecological Network Structure and Potential Function of the Bacterial Community in the Soil Profile under Indigenous Tree Plantations in Subtropical China. Forests 2023, 14, 803. https://doi.org/10.3390/f14040803
Qin L, Wang Y, Ming A, Xi S, Xiao Z, Teng J, Tan L. Molecular Ecological Network Structure and Potential Function of the Bacterial Community in the Soil Profile under Indigenous Tree Plantations in Subtropical China. Forests. 2023; 14(4):803. https://doi.org/10.3390/f14040803
Chicago/Turabian StyleQin, Lin, Yufeng Wang, Angang Ming, Shouhong Xi, Zhirou Xiao, Jinqian Teng, and Ling Tan. 2023. "Molecular Ecological Network Structure and Potential Function of the Bacterial Community in the Soil Profile under Indigenous Tree Plantations in Subtropical China" Forests 14, no. 4: 803. https://doi.org/10.3390/f14040803