
Stand Structure Management and Tree Diversity Conservation Based on Using Stand Factors: A Case Study in the Longwan National Nature Reserve


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Survey of Plant Communities
2.3. Measurement Indicators and Measurement Methods
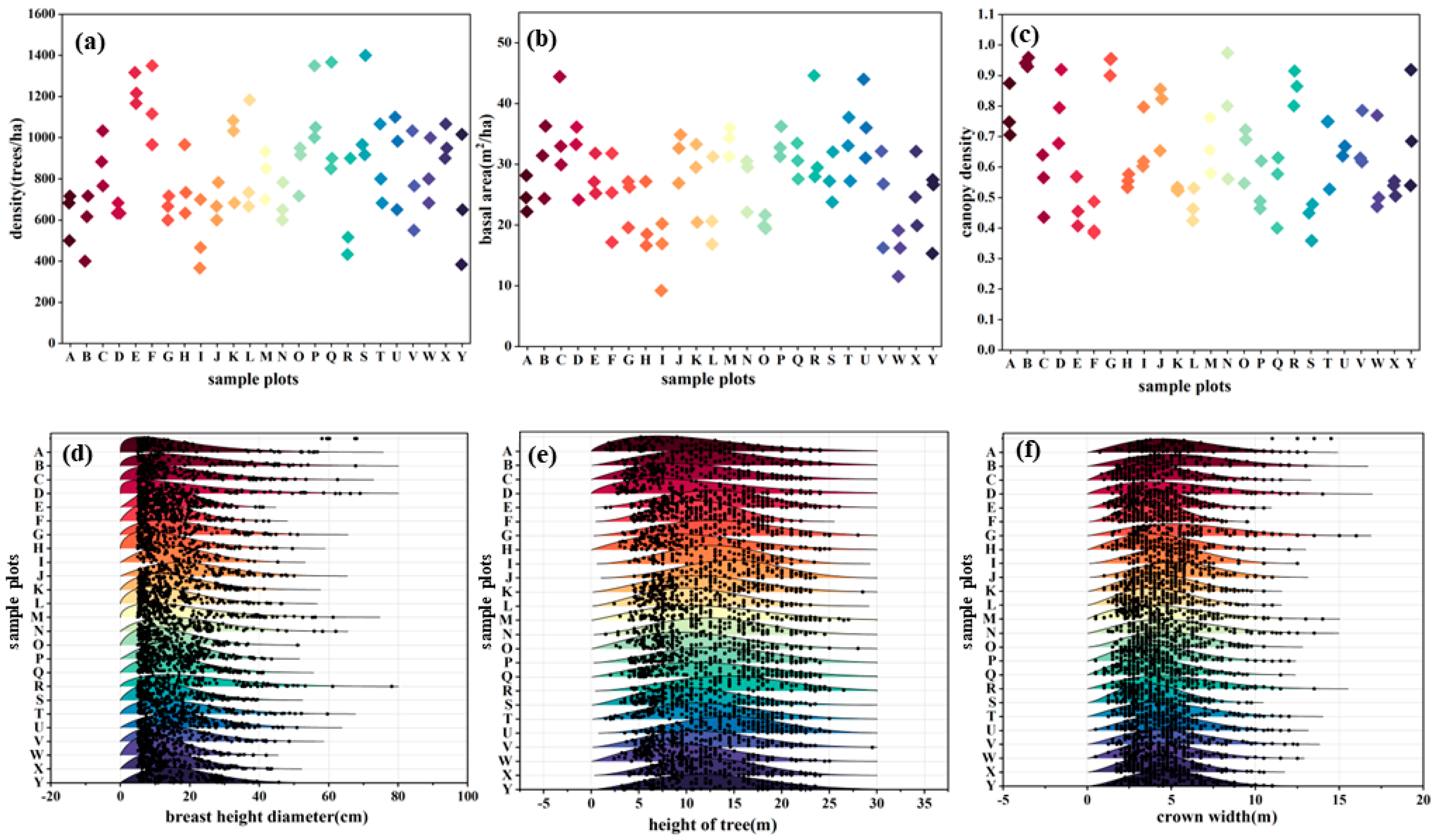
2.3.1. Stand Structure
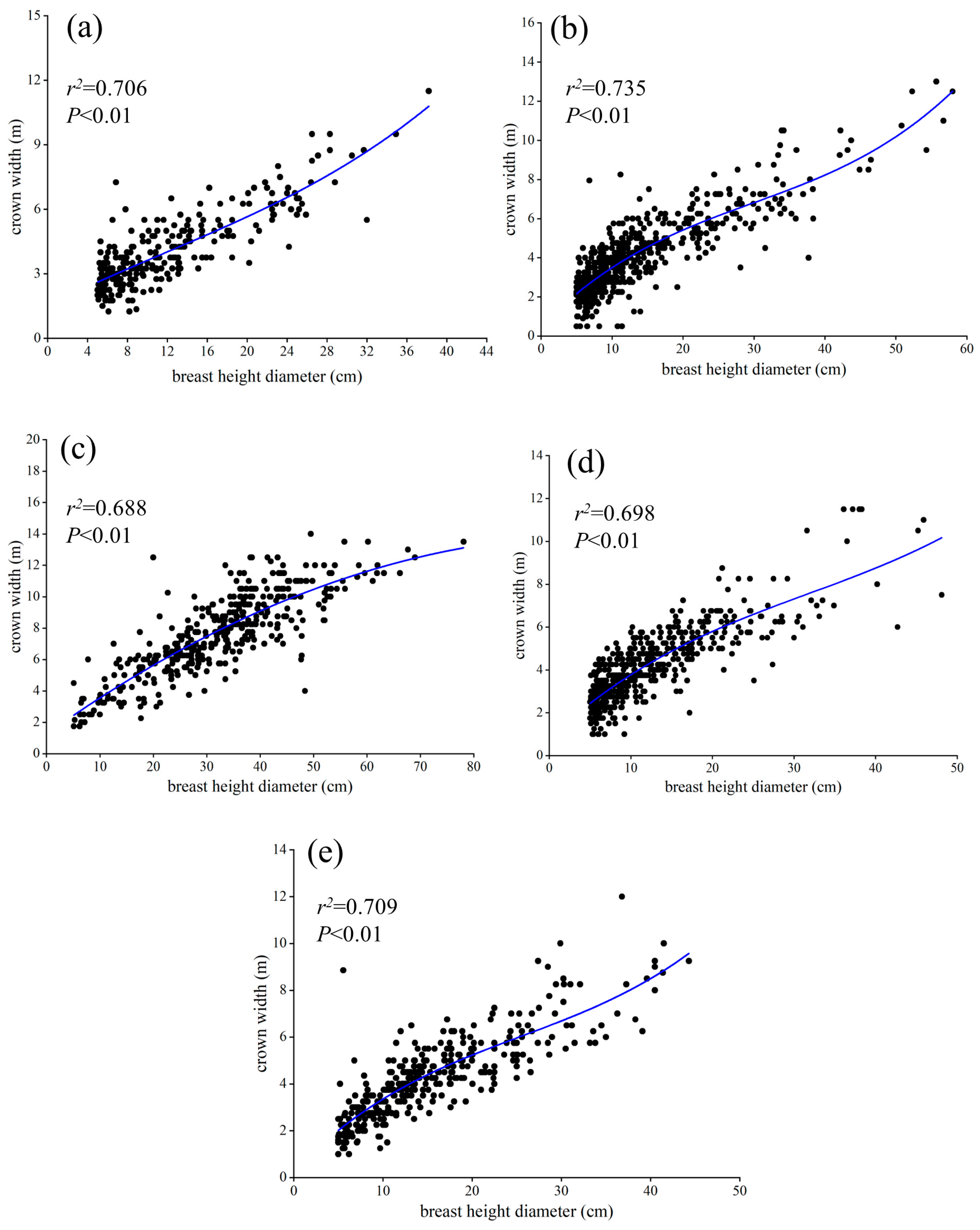
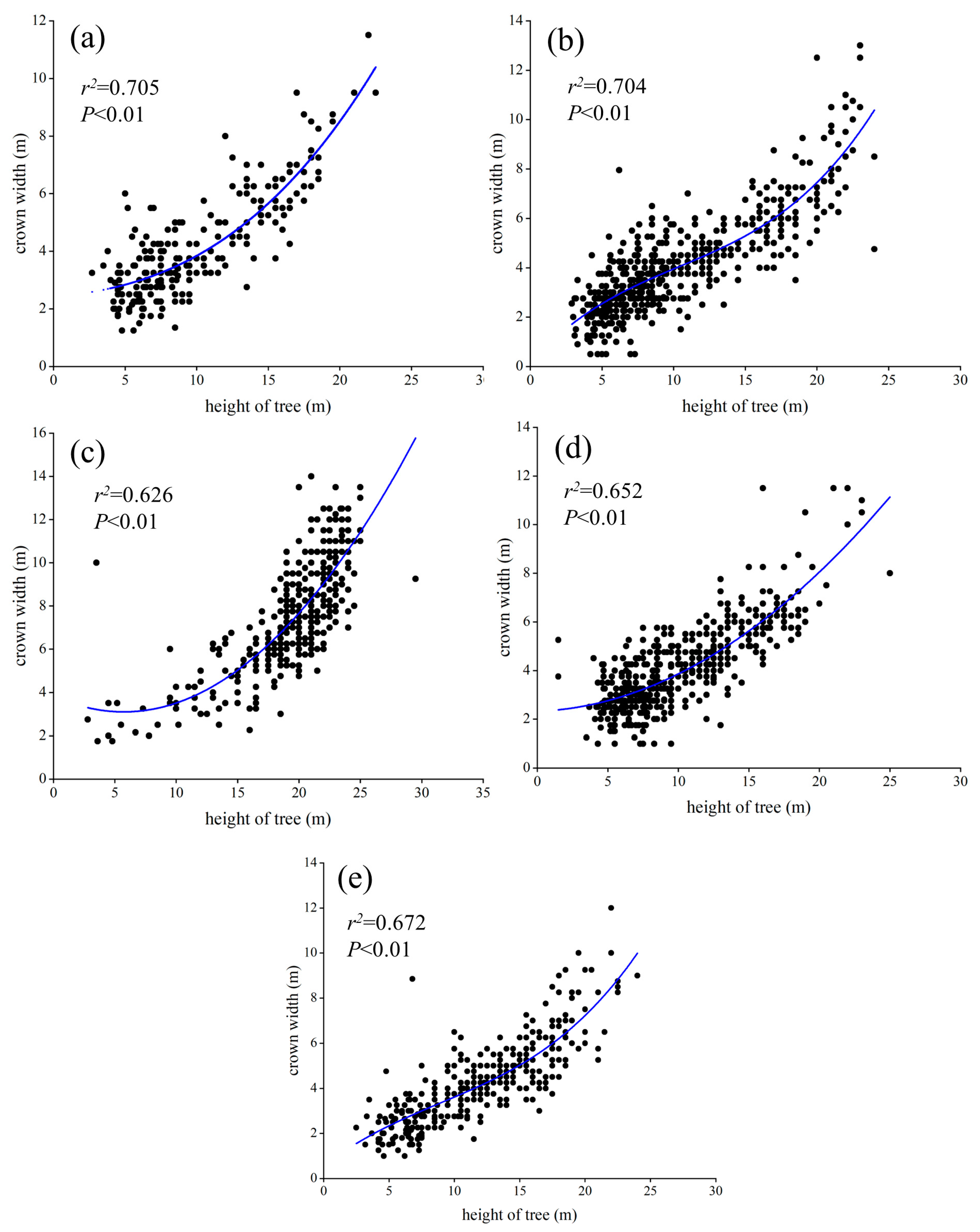
2.3.2. Construction of Allometric Growth Equation
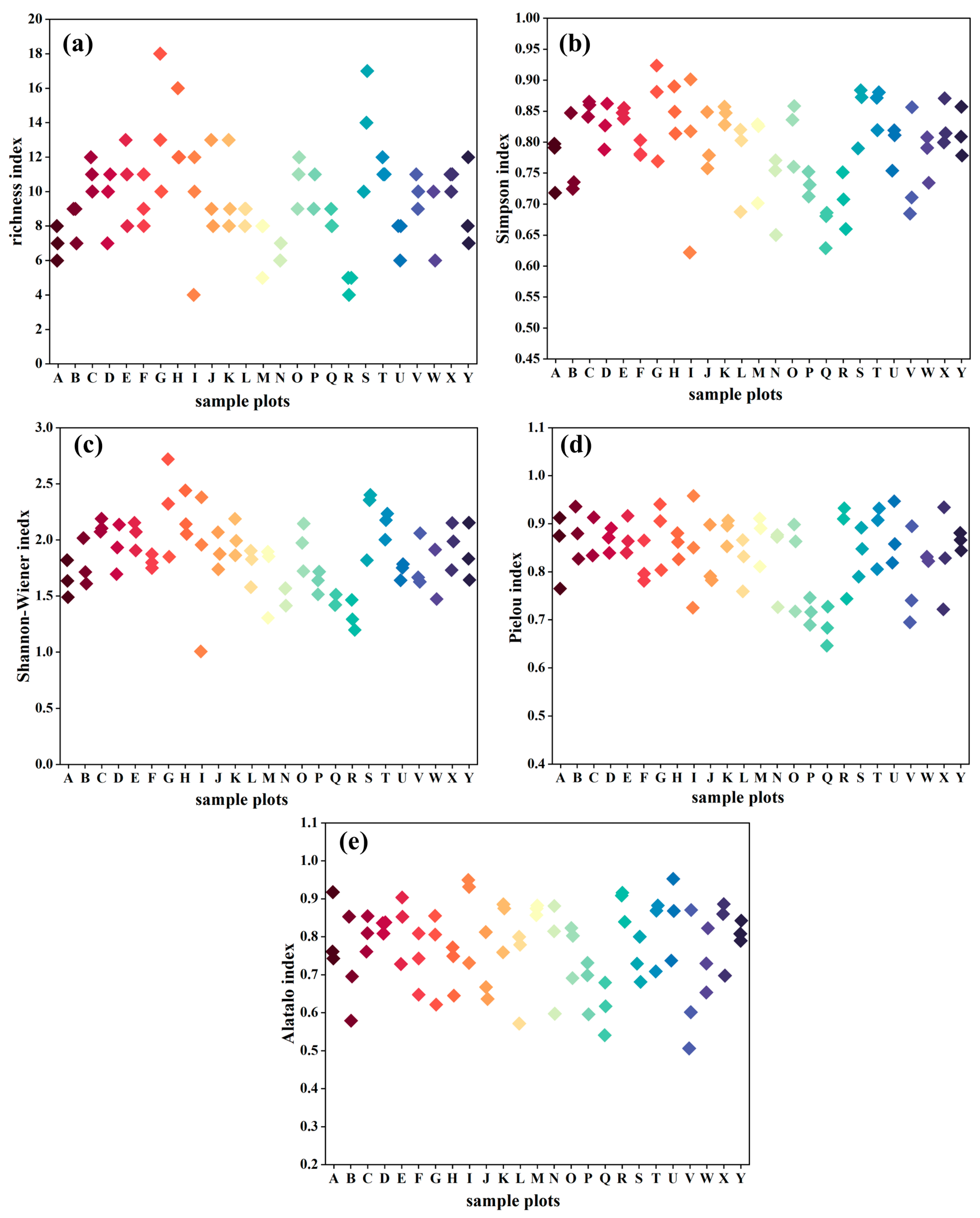
2.3.3. Plant Community Species Diversity
2.4. Data Analysis
3. Results
3.1. Species Composition and Stand Structure
3.2. Distribution Characteristics of Species Diversity
3.3. Allometric Growth Models of DBH, Tree Height, and Crown Width
3.4. Correlation Analysis of Community Structure and Species Diversity
4. Discussion
4.1. Tree-Layer Community Structure and Diversity
4.2. Allometric Growth Models of DBH, Tree Height, and Crown Width
4.3. Impact of Plant Community Structure on Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Symstad, A.J.; Chapin, F.S.; Wall, D.H.; Gross, K.L.; Huenneke, L.F.; Mittelbach, G.G.; Peters, D.P.; Tilman, D. Long-term and large-scale perspectives on the relationship between biodiversity and ecosystem functioning. Bioscience 2003, 53, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Ma, L.; Jia, Y.; Liu, M. Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China. PLoS ONE 2017, 12, e0174231. [Google Scholar] [CrossRef] [Green Version]
- Lasch, P.; Lindner, M.; Erhard, M.; Suckow, F.; Wenzel, A. Regional impact assessment on forest structure and functions under climate change—The Brandenburg case study. For. Ecol. Manag. 2002, 162, 73–86. [Google Scholar] [CrossRef]
- Pastorella, F.; Paletto, A. Stand structure indices as tools to support forest management: An application in Trentino forests (Italy). J. Trop. For. Sci. 2013, 59, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Ali, A. Forest stand structure and functioning: Current knowledge and future challenges. Ecol. Indic. 2019, 98, 665–677. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, Y.; Wang, W.; Chen, Y.; Liu, X. Characteristics of Plant Community and Its Relationship with Groundwater Depth of the Desert Riparian Zone in the Lower Reaches of the Ugan River, Northwest China. Water 2022, 14, 1663. [Google Scholar] [CrossRef]
- Li, J.; Ren, Z.; Zhou, Z. Ecosystem services and their values: A case study in the Qinba mountains of China. Ecol. Res. 2006, 21, 597–604. [Google Scholar] [CrossRef]
- Guo, Z.; Xiao, X.; Gan, Y.; Zheng, Y. Ecosystem functions, services and their values–a case study in Xingshan County of China. Ecol. Econ. 2001, 38, 141–154. [Google Scholar] [CrossRef]
- Mori, A.S.; Lertzman, K.P.; Gustafsson, L. Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest ecology. J. Appl. Ecol. 2017, 54, 12–27. [Google Scholar] [CrossRef]
- Staudhammer, C.L.; LeMay, V.M. Introduction and evaluation of possible indices of stand structural diversity. Can. J. For. Res 2001, 31, 1105–1115. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Gou, M.; Chen, L.; Lei, P.; Xiao, W.; Forrester, D.I. Stability in subtropical forests: The role of tree species diversity, stand structure, environmental and socio-economic conditions. Glob. Ecol. Biogeogr. 2021, 30, 500–513. [Google Scholar] [CrossRef]
- Cifuentes Jara, M.; Henry, M.; Réjou-Méchain, M.; Wayson, C.; Zapata-Cuartas, M.; Piotto, D.; Westfall, J. Guidelines for documenting and reporting tree allometric equations. Ann. For. Sci. 2015, 7, 763–768. [Google Scholar] [CrossRef] [Green Version]
- Forrester, D.I. Linking forest growth with stand structure: Tree size inequality, tree growth or resource partitioning and the asymmetry of competition. For. Ecol. Manag. 2019, 447, 139–157. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Rötzer, T.; Caldentey, J. Crown size and growing space requirement of common tree species in urban centres, parks, and forests—ScienceDirect. Urban For. Urban Green. 2015, 14, 466–479. [Google Scholar] [CrossRef] [Green Version]
- Subedi, T.; Bhandari, S.K.; Pandey, N. Form factor and volume equations for individual trees of Shorea robusta in Western low land of Nepal. Austrian J. For. Sci. 2021, 138, 143–166. [Google Scholar]
- Víquez, E.; Pérez, D. Effect of pruning on tree growth, yield, and wood properties of Tectona grandis plantations in Costa Rica. Silva Fenn. 2005, 39, 381. [Google Scholar] [CrossRef]
- Laubhann, D.; Sterba, H.; Reinds, G.J.; De Vries, W. The impact of atmospheric deposition and climate on forest growth in European monitoring plots: An individual tree growth model. For. Ecol. Manag. 2009, 258, 1751–1761. [Google Scholar] [CrossRef]
- Ruark, G.A.; Martin, G.L.; Bockheim, J.G. Comparison of constant and variable allometric ratios for estimating Populus tremuloides biomass. For. Sci. 1987, 33, 294–300. [Google Scholar] [CrossRef]
- Kusmana, C.; Hidayat, T.; Tiryana, T.; Rusdiana, O. Allometric models for above-and below-ground biomass of Sonneratia spp. Glob. Ecol. Conserv. 2018, 15, e00417. [Google Scholar] [CrossRef]
- Curtis, R.O.; Reukema, D.L. Crown Development and Site Estimates in a Douglas-Fir Plantation Spacing Test. For. Sci. 1970, 16, 287–301. [Google Scholar] [CrossRef]
- Sharma, R.P.; Bílek, L.; Vacek, Z.; Vacek, S. Modelling crown width–diameter relationship for Scots pine in the central Europe. Trees 2017, 31, 1875–1889. [Google Scholar] [CrossRef]
- Tumwebaze, R. Allometric biomass equations for tree species used in agroforestry systems in Uganda. Agrofor. Syst. 2013, 87, 781–795. [Google Scholar] [CrossRef]
- Troxel, B.; Piana, M.; Ashton, M.S.; Murphy-Dunning, C. Relationships between bole and crown size for young urban trees in the northeastern USA. Urban For. Urban Green. 2013, 12, 144–153. [Google Scholar] [CrossRef]
- Semenzato, P.; Cattaneo, D.; Dainese, M. Growth prediction for five tree species in an Italian urban forest. Urban For. Urban Green. 2011, 10, 169–176. [Google Scholar] [CrossRef]
- Rametsteiner, E.; Simula, M. Forest certification—An instrument to promote sustainable forest management? J. Environ. Manag. 2003, 67, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.L.; Puettmann, K.J. Overstory composition and stand structure influence herbaceous plant diversity in the mixed aspen forest of northern Minnesota. Am. Midl. Nat. 2000, 143, 111–125. [Google Scholar] [CrossRef]
- Martins, A.C.; Willig, M.R.; Presley, S.J.; Marinho-Filho, J. Effects of forest height and vertical complexity on abundance and biodiversity of bats in Amazonia. For. Ecol. Manag. 2017, 391, 427–435. [Google Scholar] [CrossRef]
- Ishii, H.T.; Tanabe, S.I.; Hiura, T. Exploring the relationships among canopy structure, stand productivity, and biodiversity of temperate forest ecosystems. For. Sci. 2004, 50, 342–355. [Google Scholar] [CrossRef]
- Kern, C.C.; Montgomery, R.A.; Reich, P.B.; Strong, T.F. Harvest-created canopy gaps increase species and functional trait diversity of the forest ground-layer community. For. Sci. 2014, 60, 335–344. [Google Scholar] [CrossRef]
- Brokaw, N.; Busing, R.T. Niche versus chance and tree diversity in forest gaps. Trends Ecol. Evol. 2000, 15, 183–188. [Google Scholar] [CrossRef]
- Spehn, E.M.; Joshi, J.; Schmid, B.; Diemer, M.; Körner, C. Above-ground resource use increases with plant species richness in experimental grassland ecosystems. Funct. Ecol. 2000, 14, 326–337. [Google Scholar] [CrossRef]
- Auclair, A.N.; Goff, F.G. Diversity relations of upland forests in the western Great Lakes area. Am. Nat. 1971, 105, 499–528. [Google Scholar] [CrossRef]
- Pitkänen, S. Correlation between stand structure and ground vegetation: An analytical approach. Plant Ecol. 1997, 131, 109–126. [Google Scholar] [CrossRef]
- Oliver, P.A.T. Patterns of shrub diversity and tree regeneration across topographic and stand-structural gradients in a Mediterranean forest. For. Syst. 2015, 24, 11. [Google Scholar] [CrossRef] [Green Version]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Ma, L.; Sun, G.Q.; Qu, Y.; Li, J.Q. Applying systematic conservation planning to constitute a protection strategy for broad-leaved Korean pine forests in Changbai Mountains, China. J. Mt. Sci. 2016, 13, 493–507. [Google Scholar] [CrossRef]
- Song, H.-J.; Ye, J.; Shi, S.; Zhang, Z.-C.; Kuang, X.; Xing, D.-L.; Yuan, Z.-Q.; Lin, F.; Wang, X.-G.; Hao, Z.-Q. Woody plant species composition and community structure in residual fragments of broad-leaved Korean pine mixed forests in Changbai Mountains area. J. Appl. Ecol. 2014, 25, 1239–1249. [Google Scholar]
- Xue, D.; Tisdell, C. Valuing ecological functions of biodiversity in Changbaishan Mountain Biosphere Reserve in northeast China. Biodivers. Conserv. 2001, 10, 467–481. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; He, C.; Jiang, H.; Sheng, L. Multi-environment factors dominate plant community structure and diversity in an ombrotrophic bog: The water level is the main regulating mechanism. Front. Environ. Sci. 2022, 10, 1032068. [Google Scholar] [CrossRef]
- Wang, X.; Bai, X.; Ma, L.; He, C.; Jiang, H.; Sheng, L.; Luo, W. Snow depths’ impact on soil microbial activities and carbon dioxide fluxes from a temperate wetland in Northeast China. Sci. Rep. 2020, 10, 8709. [Google Scholar] [CrossRef]
- Curtis, J.T.; McIntosh, R.P. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Ma, K.; Huang, J.; Yu, S.; Chen, L. Studies on the Diversity of Plant Communities in Dongling Mountain Area of Beijing: II Abundance, Evenness and Species Diversity. J. Ecol. 1995, 15, 268–277. (In Chinese) [Google Scholar]
- Zhang, L.; Du, H.; Yang, Z.; Song, T.; Zeng, F.; Peng, W.; Huang, G. Topography and Soil Properties Determine Biomass and Productivity Indirectly via Community Structural and Species Diversity in Karst Forest, Southwest China. Sustainability 2022, 14, 7644. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, B.; Zhao, S.; Piao, S.; Fang, J. Comparison of community structure and species diversity of mixed forests of deciduous broad-leaved tree and Korean pine in Northeast China. Biol. Divers. 2004, 12, 174–181. (In Chinese) [Google Scholar]
- Zhang, Y.; Chen, H.Y.; Taylor, A. Multiple drivers of plant diversity in forest ecosystems. Glob. Ecol. Biogeogr. 2014, 23, 885–893. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Pekin, B.K.; Boer, M.M.; Wittkuhn, R.S.; Macfarlane, C.; Grierson, P.F. Plant diversity is linked to nutrient limitation of dominant species in a world biodiversity hotspot. J. Veg. Sci. 2012, 23, 745–754. [Google Scholar] [CrossRef]
- Wright, J.S. Plant diversity in tropical forests: A review of mechanisms of species coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef]
- Özçelik, R.; Diamantopoulou, M.J.; Brooks, J.R.; Wiant, H.V.L., Jr. Estimating tree bole volume using artificial neural network models for four species in Turkey. J. Environ. Manag. 2010, 91, 742–753. [Google Scholar] [CrossRef]
- Santos, A. Tree allometry and crown shape of four tree species in Atlantic rain forest, south-east Brazil. J. Trop. Ecol. 2002, 18, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, M.; Zhang, J.; Lu, H.; Hauer, R.J. Allometric Growth of Common Urban Tree Species in Qingdao City of Eastern China. Forests 2023, 14, 472. [Google Scholar] [CrossRef]
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. National-scale biomass estimators for United States tree species. For. Sci. 2003, 49, 12–35. [Google Scholar] [CrossRef]
- Sawadogo, L.; Savadogo, P.; Tiveau, D.; Dayamba, S.D.; Zida, D.; Nouvellet, Y.; Oden, P.C.; Guinko, S. Allometric prediction of above-ground biomass of eleven woody tree species in the Sudanian savanna-woodland of West Africa. J. For. Res. 2020, 21, 475–481. [Google Scholar] [CrossRef]
- Peper, P.J.; Alzate, C.P.; McNeil, J.W.; Hashemi, J. Allometric equations for urban ash trees (Fraxinus spp.) in Oakville, Southern Ontario, Canada. Urban For. Urban Green. 2014, 13, 175–183. [Google Scholar] [CrossRef]
- Sun, S.; Gao, X.; Bao, W.; Wang, Z. Effects of Plantation Density of Pinus tabulaeformis on the Growth and Community Structure of Pinus tabulaeformis in the Upper Reaches of Minjiang River. J. Appl. Environ. Biol. 2005, 11, 8–13. (In Chinese) [Google Scholar]
- Li, F.; Pan, P.; Ni, J.; Lai, G.; Ouyang, X.; Xu, H.; Guo, L.; Wu, Z.; Yi, Z. Effects of spatial structure of aerial seeding Pinus massoniana forest on understory vegetation diversity. J. Northeast. For. Univ. 2016, 44, 6. (In Chinese) [Google Scholar]
- Espinosa, C.I.; Cabrera, O.; Luzuriaga, A.L.; Escudero, A. What Factors Affect Diversity and Species Composition of Endangered Tumbesian Dry Forests in Southern Ecuador? Biotropica 2011, 43, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Schoonmaker, P.; McKee, A. Species composition and diversity during secondary succession of coniferous forests in the western Cascade Mountains of Oregon. For. Sci. 1988, 34, 960–979. [Google Scholar] [CrossRef]
- Király, I.; Ódor, P. The effect of stand structure and tree species composition on epiphytic bryophytes in mixed deciduous–coniferous forests of Western Hungary. Biol. Conserv. 2010, 143, 2063–2069. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Chen, L.; Xie, G.; Liu, C.; Pei, S. Effects of topographical and edaphic factors on tree community structure and diversity of subtropical mountain forests in the Lower Lancang River Basin. Forests 2016, 7, 222. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Model | Parameter Estimation | Evaluation Index | Significance Test | ||||
|---|---|---|---|---|---|---|---|---|
| a | b | c | R2 | RSS | F-Value | p-Value | ||
| Acer mandshuricum | CW = aDb | 0.807 | 0.646 | 0.624 | 15.750 | 382.590 | 0.000 | |
| CW = aDbHc | 0.505 | 0.716 | 0.273 | 0.551 | 17.773 | |||
| Ulmus japonica | CW = aDb | 0.685 | 0.682 | 0.579 | 57.574 | |||
| CW = aDbHc | 0.608 | 0.381 | 0.390 | 0.607 | 53.785 | 422.307 | 0.000 | |
| Juglans mandshurica | CW = aDb | 0.728 | 0.678 | 0.716 | 16.549 | |||
| CW = aDbHc | 0.502 | 0.533 | 0.295 | 0.738 | 14.264 | 534.089 | 0.000 | |
| Acer mono | CW = aDb | 0.827 | 0.644 | 0.618 | 34.490 | |||
| CW = aDbHc | 0.795 | 0.540 | 0.157 | 0.636 | 30.290 | 477.96 | 0.000 | |
| Tilia amurensis | CW = aDb | 0.637 | 0.698 | 0.708 | 19.521 | |||
| CW = aDbHc | 0.541 | 0.484 | 0.301 | 0.782 | 17.926 | 460.426 | 0.000 | |
| Ra | Da | Ha | Jsw | Ea | Dy | DBH | TH | CW | BC | CD | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ra | 1 | ||||||||||
| Da | 0.658 ** | 1 | |||||||||
| Ha | 0.853 ** | 0.919 ** | 1 | ||||||||
| Jsw | 0.108 | 0.630 ** | 0.466 ** | 1 | |||||||
| Ea | −0.204 | 0.430 ** | 0.137 | 0.672 ** | 1 | ||||||
| Dy | 0.321 ** | 0.060 | 0.128 | −0.271 * | −0.248 * | 1 | |||||
| DBH | −0.265 * | −0.077 | −0.161 | 0.243 * | 0.232 * | −0.346 ** | 1 | ||||
| TH | −0.215 | −0.136 | −0.194 | 0.061 | 0.192 | −0.096 | 0.700 ** | 1 | |||
| CW | −0.229 * | −0.028 | −0.043 | 0.298 ** | 0.253 * | −0.677 ** | 0.561 ** | 0.392 ** | 1 | ||
| BC | −0.033 | −0.006 | −0.035 | 0.092 | 0.044 | 0.322 ** | 0.546 ** | 0.369 ** | 0.139 | 1 | |
| CD | −0.081 | 0.067 | 0.028 | 0.196 | 0.186 | −0.481 ** | 0.762 ** | 0.480 ** | 0.721 ** | 0.191 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wang, X.; Zhang, C.; Li, J.; Jiang, H.; Sheng, L.; Li, Z. Stand Structure Management and Tree Diversity Conservation Based on Using Stand Factors: A Case Study in the Longwan National Nature Reserve. Forests 2023, 14, 750. https://doi.org/10.3390/f14040750
Li Y, Wang X, Zhang C, Li J, Jiang H, Sheng L, Li Z. Stand Structure Management and Tree Diversity Conservation Based on Using Stand Factors: A Case Study in the Longwan National Nature Reserve. Forests. 2023; 14(4):750. https://doi.org/10.3390/f14040750
Chicago/Turabian StyleLi, Yue, Xue Wang, Chaofan Zhang, Jinpeng Li, Haibo Jiang, Lianxi Sheng, and Zhenxin Li. 2023. "Stand Structure Management and Tree Diversity Conservation Based on Using Stand Factors: A Case Study in the Longwan National Nature Reserve" Forests 14, no. 4: 750. https://doi.org/10.3390/f14040750