
Soil Macrofauna Disperse and Reconstruct Soil Nematode Communities: Takeaways from a Microcosm Study
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experimental Materials
2.2. Experiment Design
2.3. Sampling and Analysis
2.4. Statistical Analysis
3. Results
3.1. Soil Properties
3.2. Nematode Abundances and Species Richness
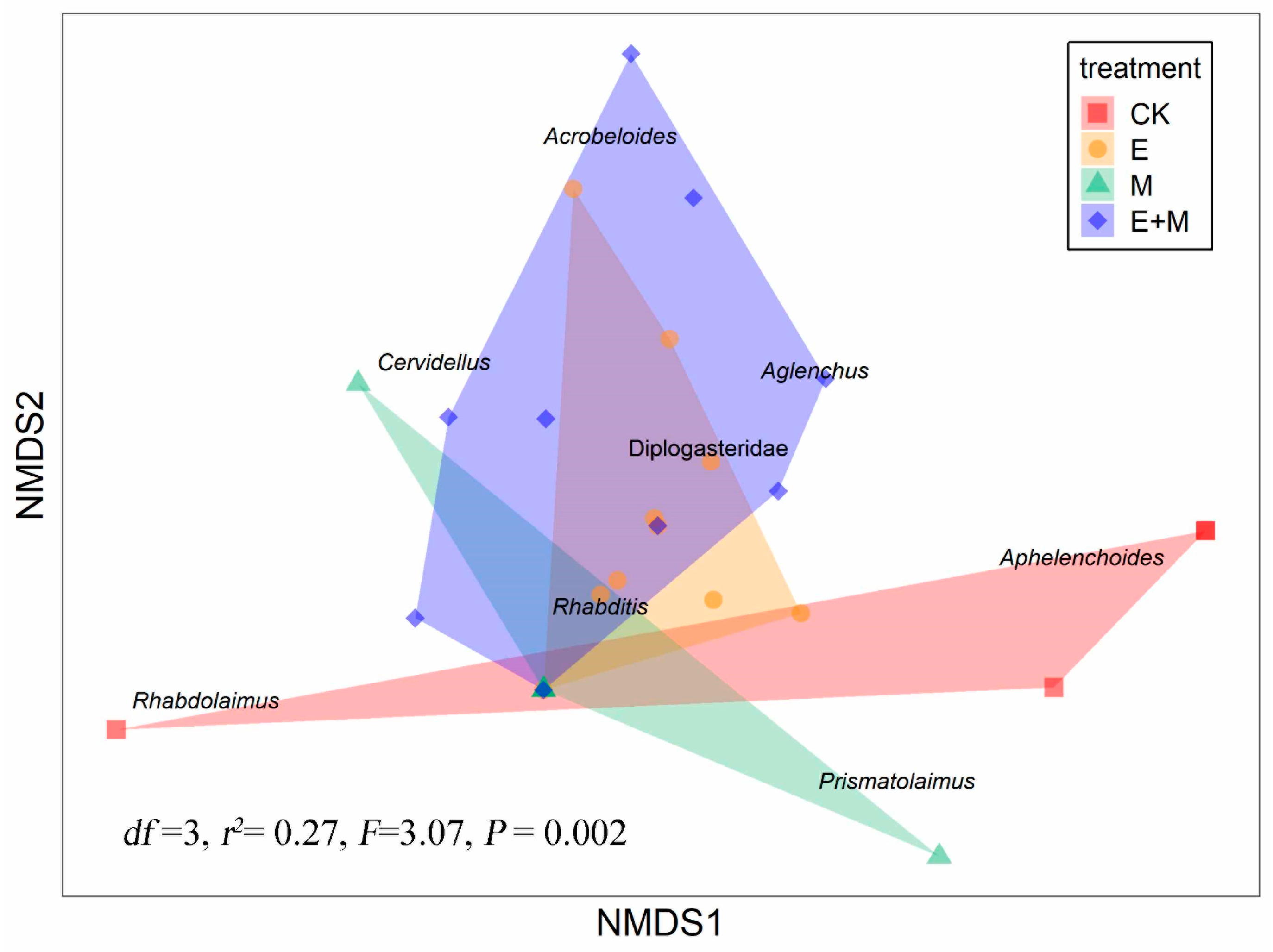
3.3. Treatment Effects on Soil Nematode Taxonomic Community Structure
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Luo, Y.H.; Cadotte, M.W.; Liu, J.; Burgess, K.S.; Tan, S.L.; Ye, L.J.; Zou, J.Y.; Chen, Z.Z.; Jiang, X.L.; Li, J.; et al. Multitrophic diversity and biotic associations influence subalpine forest ecosystem multifunctionality. Ecology 2022, 103, e3745. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Lavelle, P.; Bignell, D.; Lepage, M.; Wolters, V.; Roger, P.; Ineson, P.; Heal, O.W.; Dhillion, S. Soil function in a changing world: The role of invertebrate ecosystem engineers. Eur. J. Soil Biol. 1997, 33, 159–193. [Google Scholar]
- Lavelle, P. Functional domains in soils. Ecol. Res. 2002, 17, 441–450. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Crossley, D.A., Jr. Effects of microfauna on the seasonal dynamics of nutrients in forest litter. Soil Biol. Biochem. 1980, 12, 337–342. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Crossley, D.A., Jr. Nutrients in forest litter treated with naphthalene and simulated throughfall: A field microcosm study. Soil Biol. Biochem. 1983, 15, 159–165. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Chan, K.F. Experimental evidence that soil fauna enhance nutrient mineralization and plant nutrient uptake in montane grassland ecosystems. Soil Biol. Biochem. 1999, 31, 1007–1014. [Google Scholar] [CrossRef]
- Hasegawa, M.; Takeda, H. Carbon and nutrient dynamics in decomposing pine needle litter in relation to fungal and fauna abundances. Pedobiologia 1996, 40, 171–184. [Google Scholar]
- Topp, W.; Kappes, H.; Kulfan, J.; Zach, P. Distribution pattern of woodlice (Isopoda) and millipedes (Diplopoda) in four primeval forests of the Western Carpathians (Central Slovakia). Soil Biol. Biochem. 2006, 38, 43–50. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Partsch, S.; Parkinson, D.; Scheu, S. Invasion of a deciduous forest by earthworms: Changes in soil chemistry, microflora, microarthropods and vegetation. Soil Biol. Biochem. 2007, 39, 1099–1110. [Google Scholar] [CrossRef]
- Scheu, S.; Wolters, V. Influence of fragmentation and bioturbation on the decomposition of 14C-labelled beech leaf litter. Soil Biol. Biochem. 1991, 23, 1029–1034. [Google Scholar] [CrossRef]
- Edwards, C.A.; Bohlen, P.J. Biology and Ecology of Earthworms, 3rd ed.; Chapman and Hall: London, UK, 1996. [Google Scholar] [CrossRef]
- Edwards, C.A.; Fletcher, K.E. Interactions between earthworms and micro-organisms in organic matter breakdown. Agric. Ecosyst. Environ. 1988, 24, 235–247. [Google Scholar] [CrossRef]
- van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Neher, D.A. Role of nematodes in soil health and their use as indicators. J. Nematol. 2001, 33, 161–168. [Google Scholar] [PubMed]
- Liu, T.; Mao, P.; Shi, L.; Eisenhauer, N.; Liu, S.; Wang, X.; He, X.; Wang, Z.; Zhang, W.; Liu, Z.; et al. Forest canopy maintains the soil community composition under elevated nitrogen deposition. Soil Biol. Biochem. 2020, 143, 107733. [Google Scholar] [CrossRef]
- Kudrin, A.A.; Zuev, A.G.; Taskaeva, A.A.; Konakova, T.N.; Kolesnikova, A.A.; Gruzdev, I.V.; Gabov, D.N.; Yakovleva, E.V.; Tiunov, A.V. Spruce girdling decreases abundance of fungivorous soil nematodes in a boreal forest. Soil Biol. Biochem. 2021, 155, 108184. [Google Scholar] [CrossRef]
- Liu, T.; Mao, P.; Shi, L.; Wang, Z.; Wang, X.; He, X.; Tao, L.; Liu, Z.; Zhou, L.; Shao, Y.; et al. Contrasting effects of nitrogen deposition and increased precipitation on soil nematode communities in a temperate forest. Soil Biol. Biochem. 2020, 148, 107869. [Google Scholar] [CrossRef]
- Liu, T.; Wang, X.; Zhang, W.; Eisenhauer, N.; Shao, Y.; Zhao, J.; Li, J.; He, X.; Tao, L.; Fu, S.; et al. Earthworms regulate the nematode community by directly enhancing the bacterial-based energy channel rather than through the effect of casts. Pedobiologia 2022, 95, 150843. [Google Scholar] [CrossRef]
- Yeates, G.W. Soil nematode populations depressed in the presence of earthworms. Pedobiologia 1981, 22, 191–195. [Google Scholar]
- Hyvönen, R.; Andersson, S.; Clarholm, M.; Persson, T. Effects of lumbricids and enchytraeids on nematodes in limed and unlimed coniferous mor humus. Biol. Fertil. Soils 1994, 17, 201–205. [Google Scholar] [CrossRef]
- Domínguez, J.; Parmelee, R.W.; Edwards, C.A. Interaction between Eisenia andrei (Oligochaeta) and nematode populations during vermicomposting. Pedobiologia 2003, 47, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Li, C.K.; Liu, T.; Li, T.; Hu, F.; Li, H.X.; Jiao, J.G. Earthworm mucus interfere in the behavior and physiology of bacterial-feeding nematodes. Appl. Soil Ecol. 2019, 143, 107–115. [Google Scholar] [CrossRef]
- Senapati, B.K. Biotic interactions between soil nematodes and earthworms. Soil Biol. Biochem. 1992, 24, 1441–1444. [Google Scholar] [CrossRef]
- Boyer, J.; Reversat, G.; Lavelle, P.; Chabanne, A. Interactions between earthworms and plant-parasitic nematodes. Eur. J. Soil Biol. 2013, 59, 43–47. [Google Scholar] [CrossRef]
- Shapiro, D.I.; Berry, E.C.; Lewis, L.C. Interactions between nematodes and earthworms: Enhanced dispersal of Steinernema carpocapsae. J. Nematol. 1993, 25, 189–192. [Google Scholar] [PubMed]
- Villenave, C.; Rabary, B.; Kichenin, E.; Djigal, D.; Blanchart, E. Earthworms and plant residues modify nematodes in tropical cropping soils (Madagascar): A mesocosm experiment. Appl. Environ. Soil Sci. 2010, 323640. [Google Scholar] [CrossRef] [Green Version]
- Phillips, G.; Yates, D.I.; Shelley, R.M.; Ortstadt, P.R.; Bernard, E.C. Investigating commensal relationships of nematodes in millipedes: Life in unexpected Places. Am. Biol. Teach. 2019, 81, 278–283. [Google Scholar] [CrossRef]
- Phillips, D.; Bernard, E.C.; Shelley, R.M.; Sun, X. Diversity, abundance and prevalence of nematodes inside the intestine of North American millipedes. J. Nematol. 2016, 48, 362. [Google Scholar]
- Senapati, B.K.; Dash, M.C. Effect of grazing on the elements of production in vegetation and Oligochaete components of a tropical pasture. Rev. Decol. Biol. Sol. 1981, 18, 487–505. [Google Scholar]
- Aira, M.; Sampedro, L.; Monroy, F.; Domínguez, J. Detritivorous earthworms directly modify the structure, thus altering the functioning of a microdecomposer food web. Soil Biol. Biochem. 2008, 40, 2511–2516. [Google Scholar] [CrossRef]
- Barker, K.R. Nematode extraction and bioassays. In An Advanced Treatise on Meloidogyne; Barker, K.R., Carter, C.C., Sasser, J.N., Eds.; Methodology; North Carolina State University: Raleigh, NC, USA, 1985; Volume 2, pp. 19–35. [Google Scholar]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.L.C.; Guan, P.; Cui, S.; de Tomasel, C.M.; Gherardi, L.A.; Sala, O.E.; Wall, D.H. Precipitation effects on nematode diversity and carbon footprint across grasslands. Glob. Chang. Biol. 2022, 28, 2124–2132. [Google Scholar] [CrossRef]
- Sutherland, W.J.; Freckleton, R.P.; Godfray, H.; Beissinger, S.R.; Benton, T.; Cameron, D.D.; Carmel, Y.; Coomes, D.A.; Coulson, T.; Emmerson, M.C.; et al. Identification of 100 fundamental ecological questions. J. Ecol. 2013, 101, 58–67. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, X.; Zhang, W.; Lu, X.; Liu, T. Effects of exotic and native earthworms on soil micro-decomposers in a subtropical forest: A field mesocosm experiment. Forests 2022, 13, 1924. [Google Scholar] [CrossRef]
- Niu, X.; Zhai, P.; Zhang, W.; Gu, Y. Effects of Earthworms and Agricultural Plant Species on the Soil Nematode Community in a Microcosm Experiments. Sci. Rep. 2019, 9, 11660. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Bignell, D.E. Bacteria in the food, gut contents and faeces of the litter-feeding millipede Glomeris marginata (Villers). Soil Biol. Biochem. 1980, 12, 251–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | E | M | Depth | E × M | E × Depth | M × Depth | E × M × Depth | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F(1,32) | p | F(1,32) | p | F(1,32) | p | F(1,32) | p | F(1,32) | p | F(1,32) | p | F(1,32) | p | |
| Soil properties | ||||||||||||||
| SOC | 30.266 | <0.001 | 9.313 | 0.005 | 0.475 | 0.496 | 0.182 | 0.673 | 2.182 | 0.149 | 3.664 | 0.065 | 0.206 | 0.653 |
| STN | 27.263 | <0.001 | 9.580 | 0.004 | 0.040 | 0.842 | 1.860 | 0.182 | 1.820 | 0.187 | 2.502 | 0.124 | 0.303 | 0.586 |
| pH | 31.744 | <0.001 | 49.874 | <0.001 | 1.794 | 0.190 | 3.554 | 0.069 | 18.259 | <0.001 | 0.788 | 0.381 | 4.028 | 0.053 |
| SWC | 40.931 | <0.001 | 3.573 | 0.068 | 38.377 | <0.001 | 1.645 | 0.209 | 0.769 | 0.387 | 0.016 | 0.901 | 0.229 | 0.636 |
| Soil nematodes | ||||||||||||||
| Total nematodes | 42.515 | <0.001 | 2.083 | 0.159 | 0.035 | 0.854 | 0.310 | 0.581 | 0.886 | 0.354 | 0.234 | 0.632 | 0.025 | 0.876 |
| Bacterivores (Ba) | 47.880 | <0.001 | 3.010 | 0.092 | 0.021 | 0.887 | 0.676 | 0.417 | 0.867 | 0.359 | 0.159 | 0.693 | 0.055 | 0.816 |
| Ba1 | 29.823 | <0.001 | 2.095 | 0.158 | 0.538 | 0.469 | 1.707 | 0.201 | 1.031 | 0.318 | 0.033 | 0.857 | 0.214 | 0.647 |
| Species richness | 30.215 | <0.001 | 0.002 | 0.964 | 0.646 | 0.427 | 1.796 | 0.190 | 1.249 | 0.272 | 1.249 | 0.272 | 0.072 | 0.790 |
| Variable | E | M | E × M | |||
|---|---|---|---|---|---|---|
| F(1,16) | p | F(1,16) | p | F(1,16) | p | |
| Total nematodes | 0.239 | 0.631 | 0.036 | 0.852 | 0.694 | 0.417 |
| Bacterivores (Ba) | 0.027 | 0.871 | 0.477 | 0.500 | 4.193 | 0.057 |
| Ba1 | 1.109 | 0.308 | 0.029 | 0.867 | 1.515 | 0.236 |
| Species richness | 0.018 | 0.896 | 0.018 | 0.896 | 0.258 | 0.618 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.; Wang, Z.; Guan, H.; Zhong, B.; He, X.; Wang, Y.; Qi, Y.; Yan, W.; Lu, X. Soil Macrofauna Disperse and Reconstruct Soil Nematode Communities: Takeaways from a Microcosm Study. Forests 2023, 14, 748. https://doi.org/10.3390/f14040748
Liu T, Wang Z, Guan H, Zhong B, He X, Wang Y, Qi Y, Yan W, Lu X. Soil Macrofauna Disperse and Reconstruct Soil Nematode Communities: Takeaways from a Microcosm Study. Forests. 2023; 14(4):748. https://doi.org/10.3390/f14040748
Chicago/Turabian StyleLiu, Tao, Zixuan Wang, Huiling Guan, Buqing Zhong, Xinxing He, Yihan Wang, Yifei Qi, Wende Yan, and Xiankai Lu. 2023. "Soil Macrofauna Disperse and Reconstruct Soil Nematode Communities: Takeaways from a Microcosm Study" Forests 14, no. 4: 748. https://doi.org/10.3390/f14040748