
Enhancing Soil Quality of Short Rotation Forest Operations Using Biochar and Manure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Soil Amendments and Wood Stakes
2.3. Experimental Design, Soil Treatments, and Wood Stake Installation
2.4. Soil and Wood Stake Sampling
2.5. Laboratory Assessments
2.6. Calculations and Statistical Analyses
3. Results
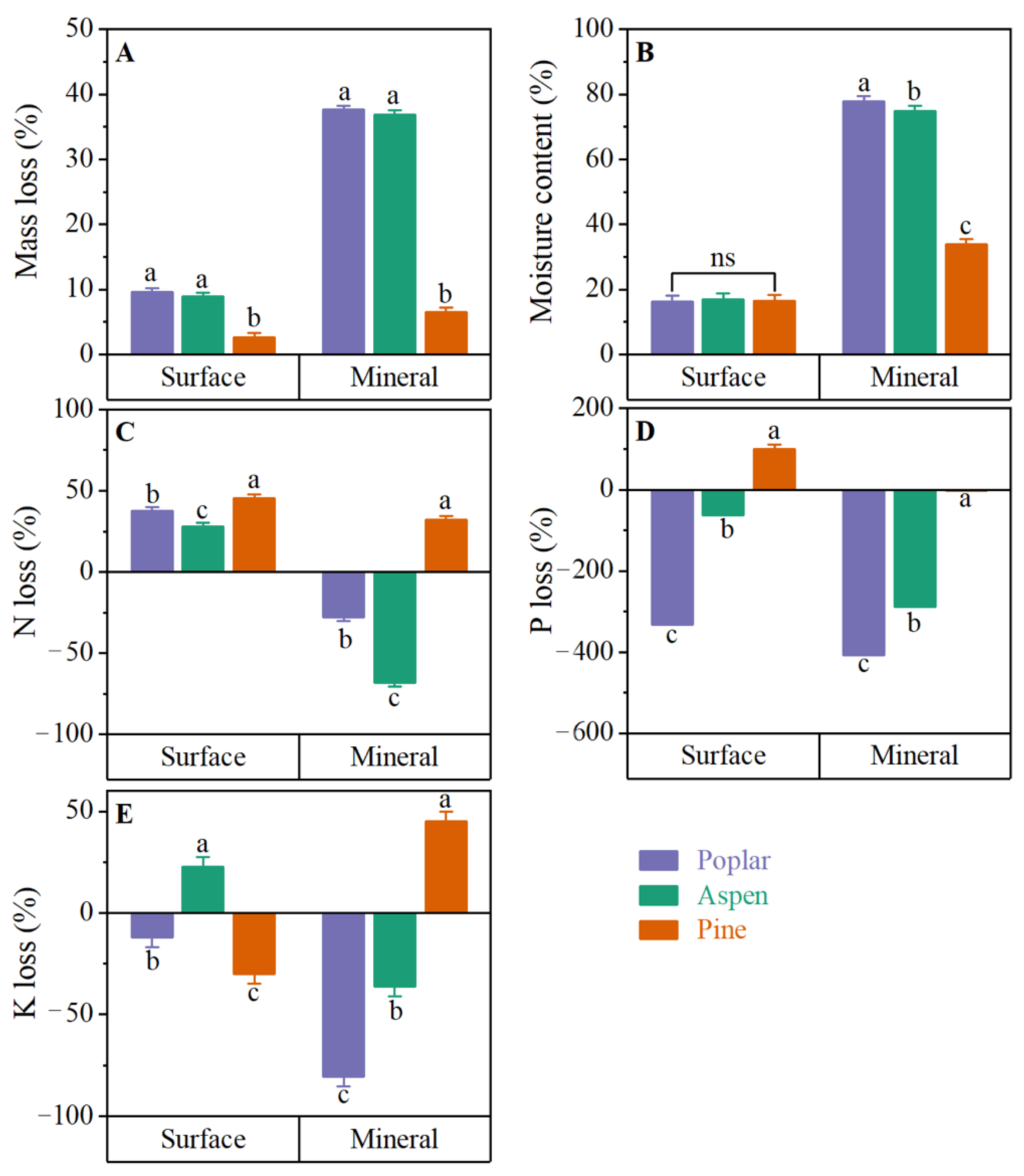
3.1. Wood Stake Decomposition
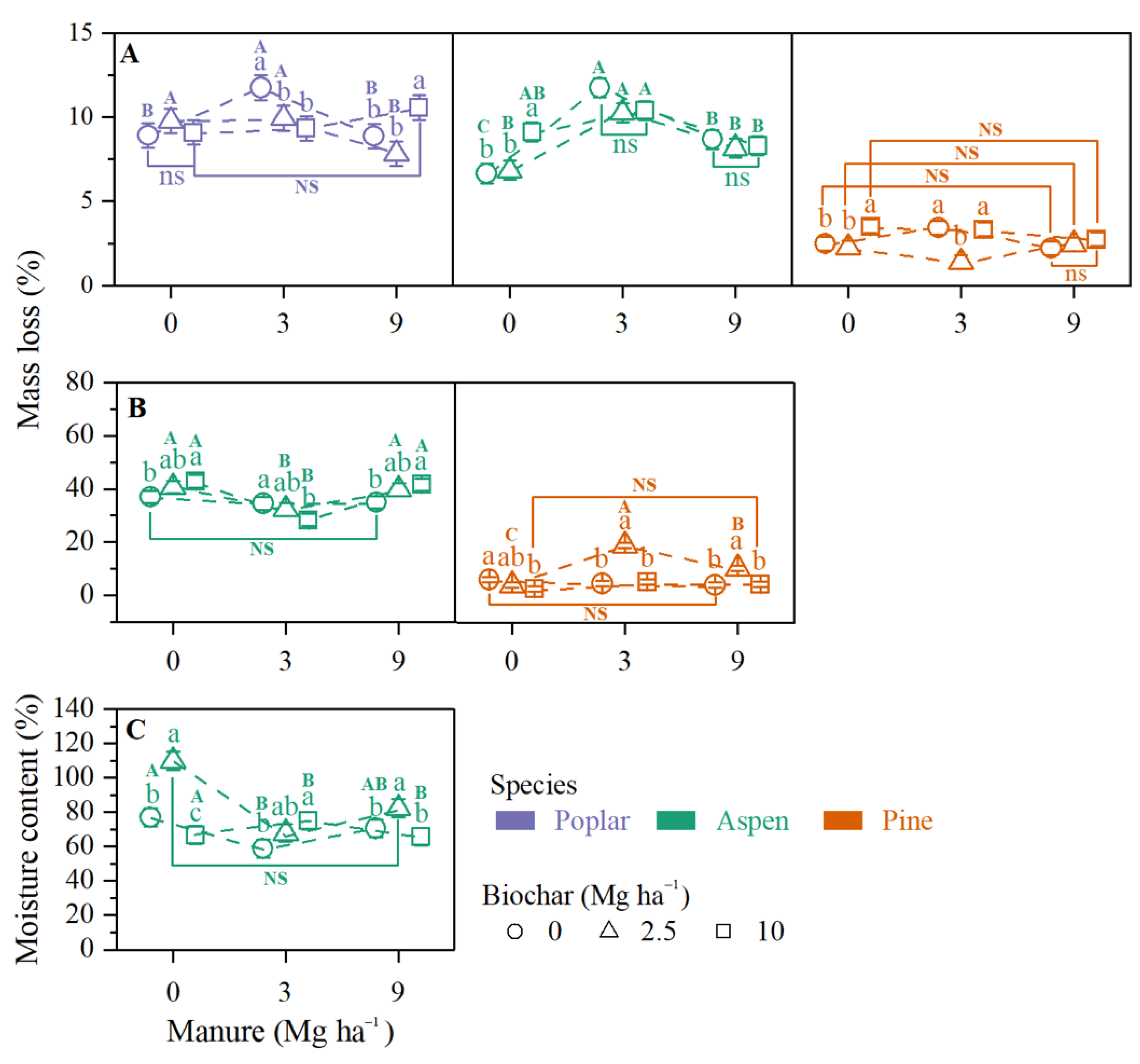
3.1.1. Wood Stake Mass Loss
3.1.2. Wood Stake Moisture Content
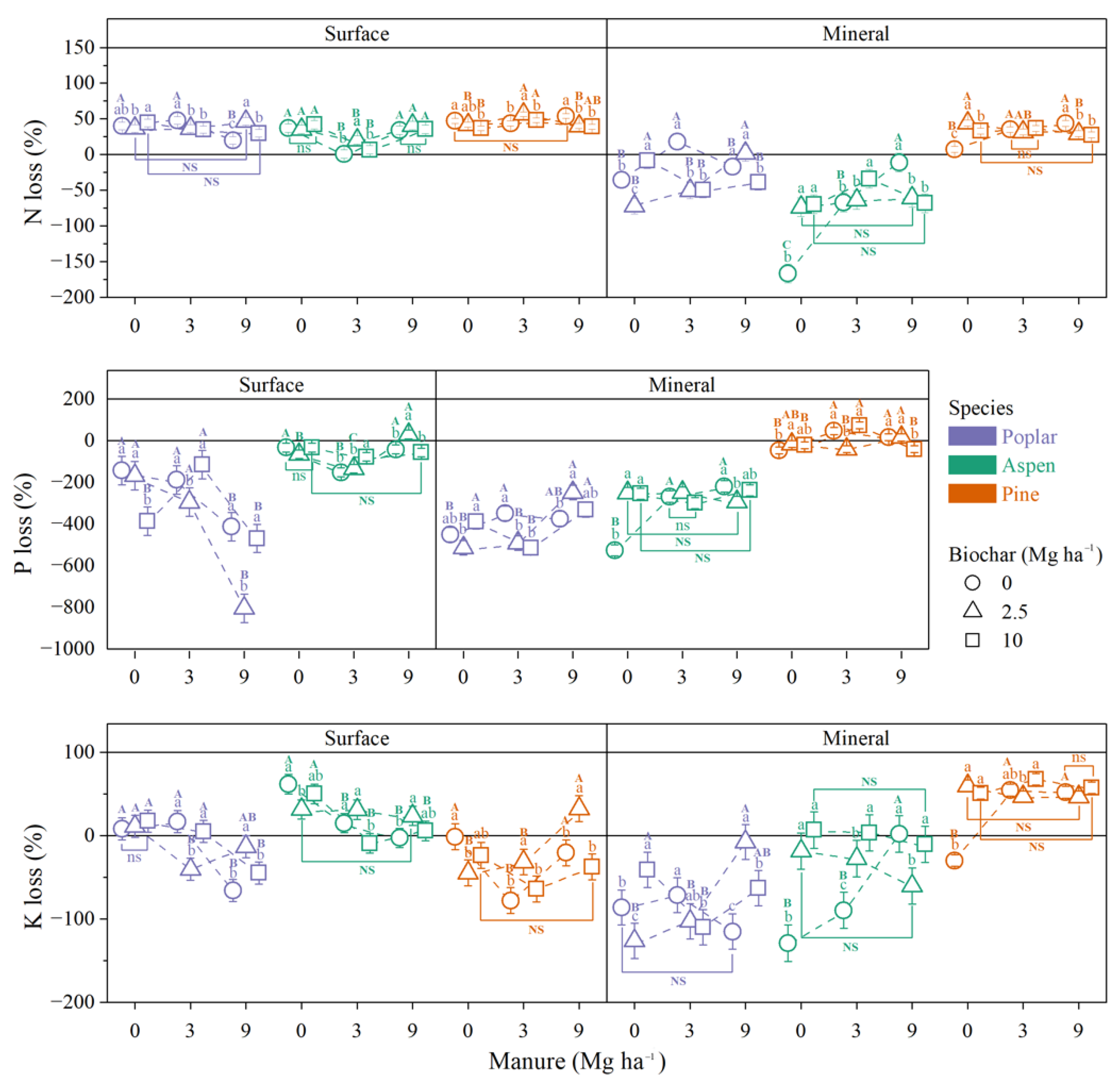
3.1.3. Wood Stake Nutrient Flux
Wood Stake N Loss
Wood Stake P Loss
Wood Stake K Loss
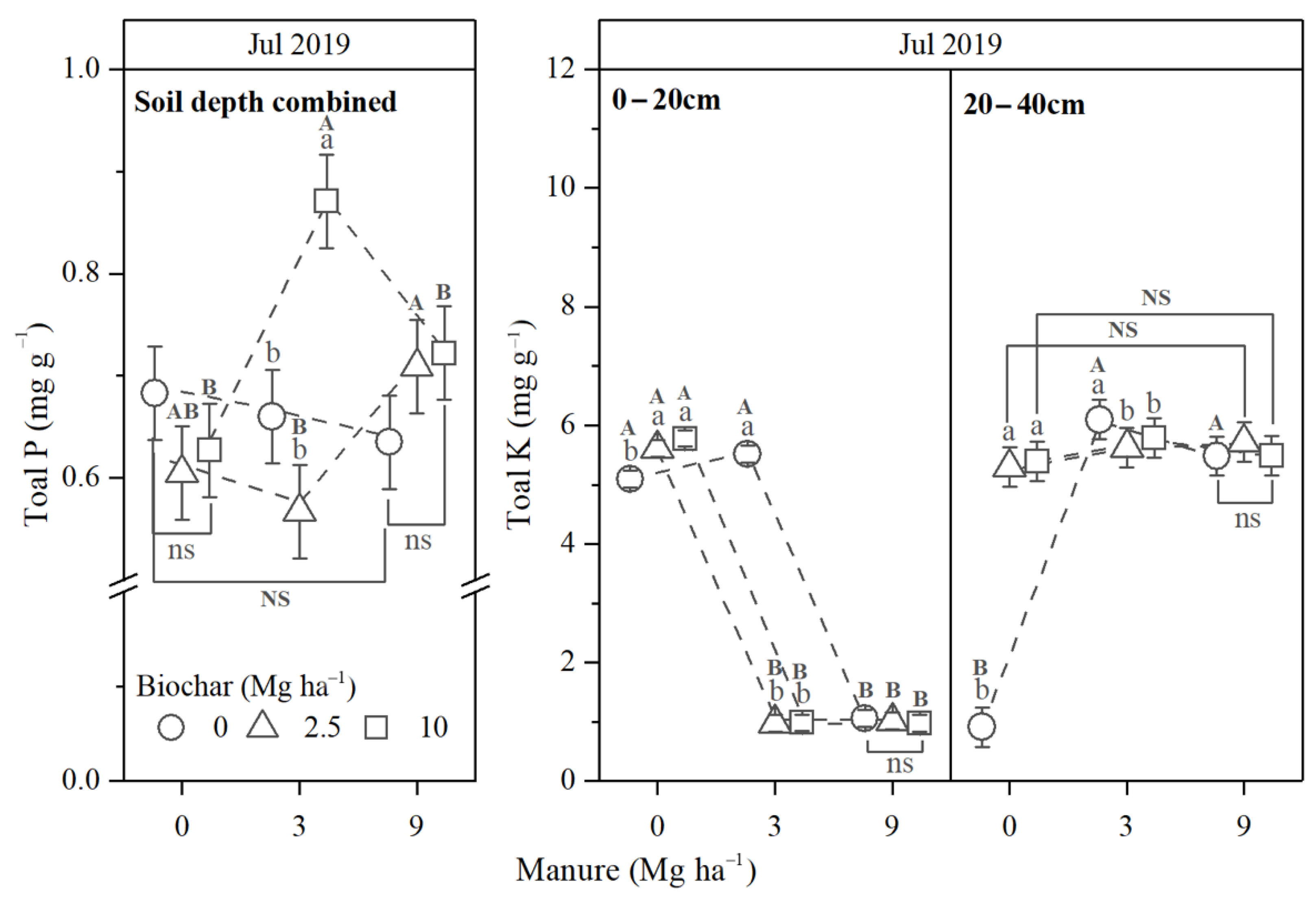
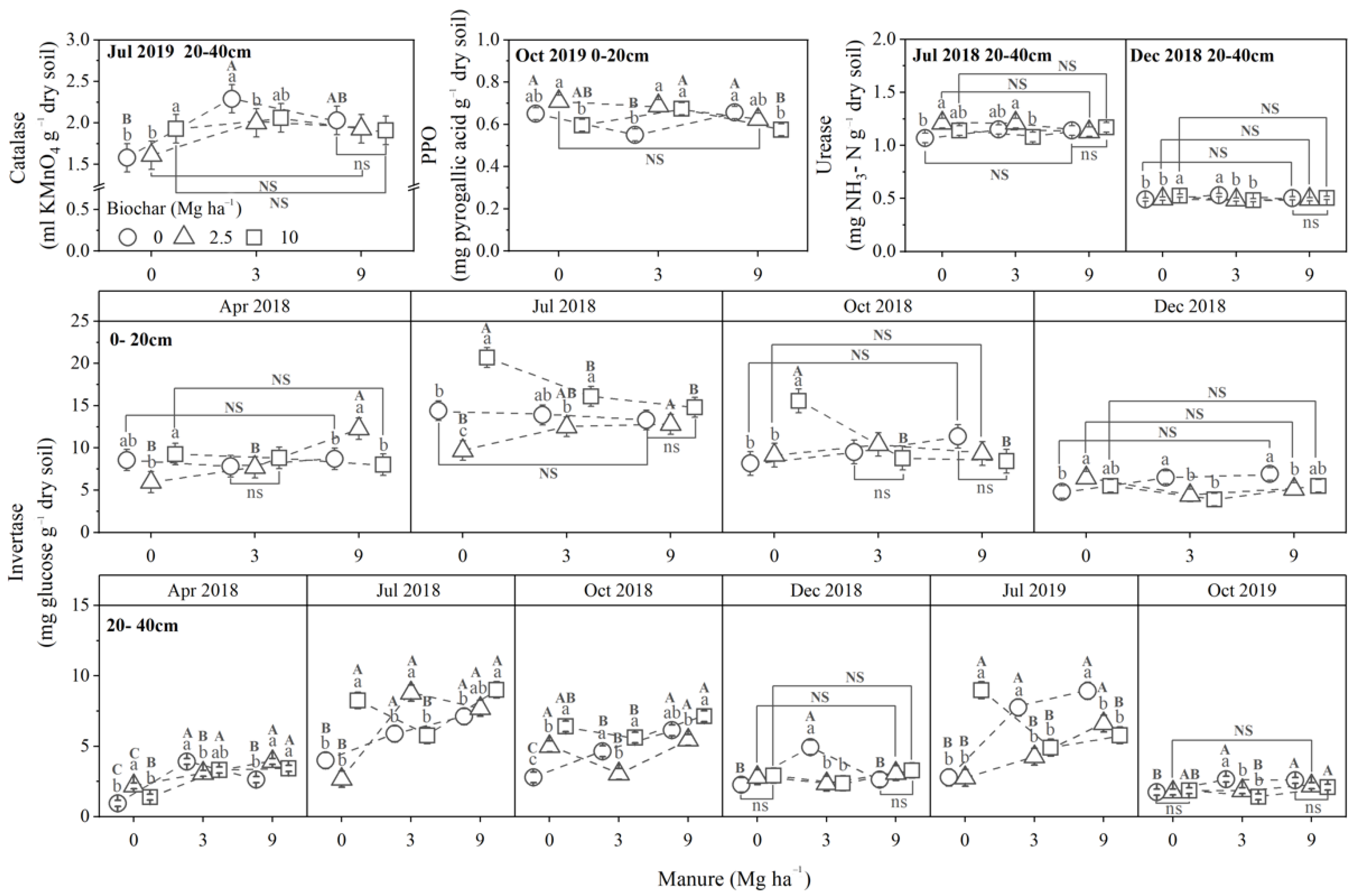
3.2. Soil Properties
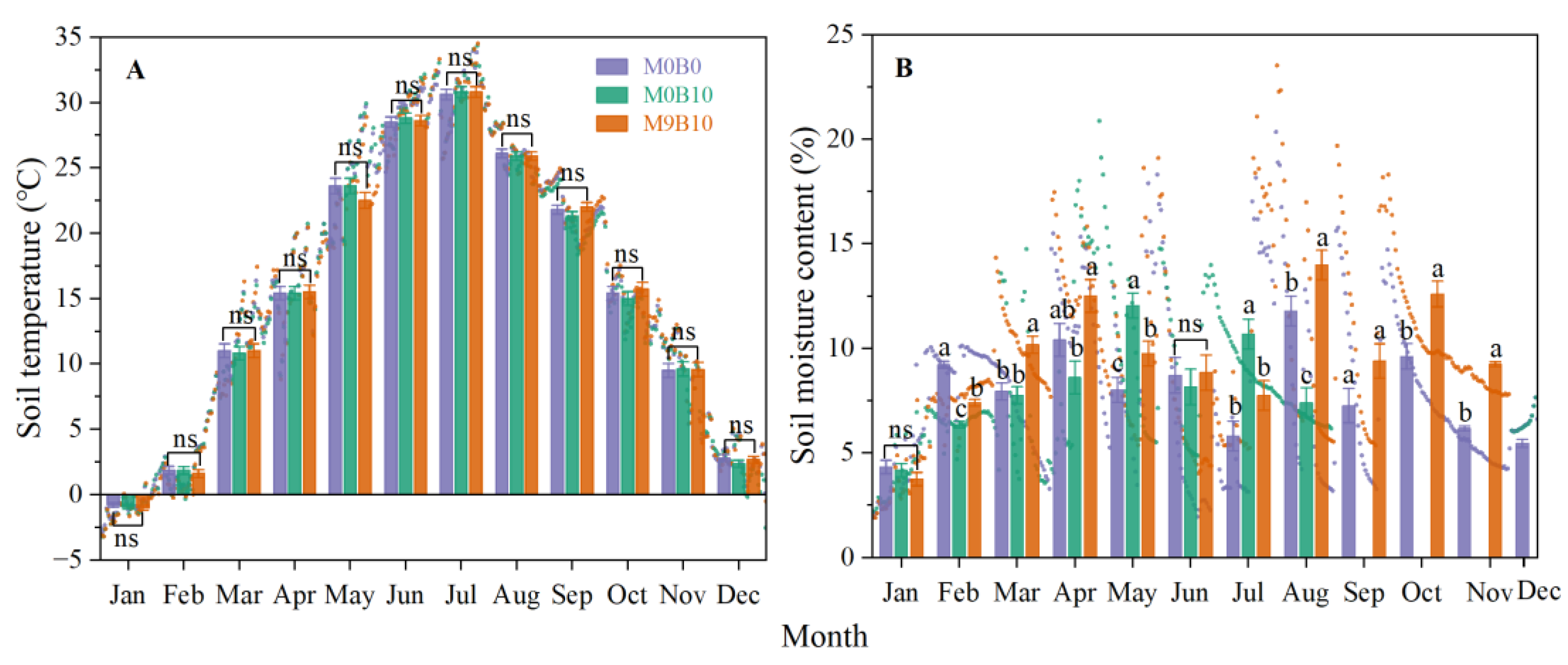
3.2.1. Soil Physical Properties
3.2.2. Soil Nutrients
3.2.3. Soil Enzyme Activities
4. Discussion
4.1. Wood Stake Mass Loss and Moisture Content
4.2. Wood Stake Nutrient Flux
4.3. Soil Physical, Chemical, and Biological Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Q.; Liu, B.J.; Zhang, Y.H.; Lin, Z.B.; Zhu, T.B.; Sun, R.B.; Wang, X.J.; Ma, J.; Bei, Q.C.; Liu, G.; et al. Can biochar alleviate soil compaction stress on wheat growth and mitigate soil N2O emissions? Soil Biol. Biochem. 2017, 104, 8–17. [Google Scholar] [CrossRef]
- Li, Y.P.; Wang, J.; Shao, M.A. Assessment of earthworms as an indicator of soil degradation: A case-study on loess soils. Land Degrad. Dev. 2021, 32, 2606–2617. [Google Scholar] [CrossRef]
- Krupek, F.S.; Redfearn, D.; Eskridge, K.M.; Basche, A. Ecological intensification with soil health practices demonstrates positive impacts on multiple soil properties: A large-scale farmer-led experiment. Geoderma 2022, 409, 115594. [Google Scholar] [CrossRef]
- Ansari, M.A.; Choudhury, B.U.; Mandal, S.; Jat, S.L.; Meitei, C.B. Converting primary forests to cultivated lands: Long-term effects on the vertical distribution of soil carbon and biological activity in the foothills of Eastern Himalaya. J. Environ. Manag. 2022, 301, 113886. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.G.; Zhu, H.G.; Wen, Y.G.; Goodale, U.M.; Zhu, Y.L.; Yu, S.F.; Li, C.T.; Li, X.Q. Intensive management and declines in soil nutrients lead to serious exotic plant invasion in Eucalyptus plantations under successive short-rotation regimes. Land Degrad. Dev. 2020, 31, 297–310. [Google Scholar] [CrossRef]
- Martin, A.R.; Domke, G.M.; Doraisami, M.; Thomas, S.C. Carbon fractions in the world’s dead wood. Nat. Commun. 2021, 12, 889. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Busse, M.D.; Jurgensen, M.F.; Jokela, E.J. Sustaining forest soil quality and productivity. In Soils and Landscape Restoration; Stanturf, J.A., Callaham, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 63–93. [Google Scholar] [CrossRef]
- He, Y.L.; Xi, B.Y.; Bloomberg, M.; Jia, L.M.; Zhao, D.H. Effects of drip irrigation and nitrogen fertigation on stand growth and biomass allocation in young triploid Populus tomentosa plantations. Forest Ecol. Manag. 2020, 461, 117937. [Google Scholar] [CrossRef]
- Palviainen, M.; Aaltonen, H.; Laurén, A.; Köster, K.; Berninger, F.; Ojala, A.; Pumpanen, J. Biochar amendment increases tree growth in nutrient-poor, young Scots pine stands in Finland. For. Ecol. Manag. 2020, 474, 118362. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Sanchez, F.G.; Udawatta, R.P.; Perry, C.H.; González, G. Soil Health Assessment of Forest Soils. In Soil Health Series; Karlen, D., Stott, D.E., Mikha, M.M., Eds.; John Wiley & Sons, Incorporation: Hoboken, NJ, USA, 2021; pp. 100–138. [Google Scholar] [CrossRef]
- Shi, S.W.; Zhang, Q.Z.; Lou, Y.L.; Du, Z.L.; Wang, Q.; Hu, N.; Wang, Y.D.; Gunina, A.; Song, J.Q. Soil organic and inorganic carbon sequestration by consecutive biochar application: Results from a decade field experiment. Soil Use Manag. 2021, 37, 95–103. [Google Scholar] [CrossRef]
- Canatoy, R.C.; Cho, S.R.; Ok, Y.S.; Jeong, S.T.; Kim, P.J. Critical evaluation of biochar utilization effect on mitigating global warming in whole rice cropping boundary. Sci. Total Environ. 2022, 827, 154344. [Google Scholar] [CrossRef]
- Ghosh, D.; Maiti, S.K. Effect of invasive weed biochar amendment on soil enzymatic activity and respiration of coal mine spoil: A laboratory experiment study. Biochar 2021, 3, 519–533. [Google Scholar] [CrossRef]
- Li, J.; Wen, Y.C.; Li, X.H.; Li, Y.T.; Yang, X.D.; Lin, Z.A.; Song, Z.Z.; Cooper, J.L.M.; Zhao, B.Q. Soil labile organic carbon fractions and soil organic carbon stocks as affected by long-term organic and mineral fertilization regimes in the North China Plain. Soil Tillage Res. 2018, 175, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Ai, C.; Zhang, S.Q.; Zhang, X.; Guo, D.D.; Zhou, W.; Huang, S.M. Distinct responses of soil bacterial and fungal communities to changes in fertilization regime and crop rotation. Geoderma 2018, 319, 156–166. [Google Scholar] [CrossRef]
- Seyedsadr, S.; Sipek, V.; Jacka, L.; Snehota, M.; Beesley, L.; Pohorely, M.; Kovar, M.; Trakal, L. Biochar considerably increases the easily available water and nutrient content in low-organic soils amended with compost and manure. Chemosphere 2022, 293, 133586. [Google Scholar] [CrossRef]
- Wang, F.L.; Wang, X.X.; Song, N.N. Biochar and vermicompost improve the soil properties and the yield and quality of cucumber (Cucumis sativus L.) grown in plastic shed soil continuously cropped for different years. Agric. Ecosyst. Environ. 2021, 315, 107425. [Google Scholar] [CrossRef]
- Ren, J.H.; Liu, X.L.; Yang, W.P.; Yang, X.X.; Li, W.G.; Xia, Q.; Li, J.H.; Gao, Z.Q.; Yang, Z.P. Rhizosphere soil properties, microbial community, and enzyme activities: Short-term responses to partial substitution of chemical fertilizer with organic manure. J. Environ. Manag. 2021, 299, 113650. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.M.; Li, X.Y.; Peng, Y.T.; Xiang, P.; Zhou, Y.Z.; Yao, B.; Zhou, Y.Y.; Sun, C.R. Co-application of biochar and organic fertilizer promotes the yield and quality of red pitaya (Hylocereus polyrhizus) by improving soil properties. Chemosphere 2022, 294, 133619. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.G.; Thuy, B.T.P.; Lin, C.; Vo, D.-V.N.; Tran, H.T.; Bahari, M.B.; Le, V.G.; Vu, C.T. The nitrogen cycle and mitigation strategies for nitrogen loss during organic waste composting: A review. Chemosphere 2022, 300, 134514. [Google Scholar] [CrossRef]
- Wang, J.; Hao, X.X.; Liu, Z.L.; Guo, Z.L.; Zhu, L.; Xiong, B.J.; Jiang, D.M.; Shen, L.Y.; Li, M.Z.; Kang, B.; et al. Biochar improves heavy metal passivation during wet anaerobic digestion of pig manure. Environ. Sci. Pollut. Res. 2021, 28, 635–644. [Google Scholar] [CrossRef]
- Nguyen, M.K.; Lin, C.; Hoang, H.G.; Sanderson, P.; Dang, B.T.; Bui, X.T.; Nguyen, N.S.H.; Vo, D.-V.N.; Tran, H.T. Evaluate the role of biochar during the organic waste composting process: A critical review. Chemosphere 2022, 299, 134488. [Google Scholar] [CrossRef]
- Luo, S.W.; Ren, L.; Wu, W.J.; Chen, Y.J.; Li, G.Y.; Zhang, W.J.; Wei, T.; Liang, Y.Q.; Zhang, D.Y.; Wang, X.Z.; et al. Impacts of earthworm casts on atrazine catabolism and bacterial community structure in laterite soil. J. Hazard. Mater. 2022, 425, 127778. [Google Scholar] [CrossRef] [PubMed]
- El-Naggara, A.; Lee, S.S.; Rinklebed, J.; Farooq, M.; Song, H.; Sarmah, A.K.; Zimmerman, A.R.; Ahmad, M.; Shaheen, S.M.; Ok, Y.S. Biochar application to low fertility soils: A review of current status, and future prospects. Geoderma 2019, 337, 536–554. [Google Scholar] [CrossRef]
- Yu, M.J.; Liang, S.J.; Dai, Z.M.; Li, Y.; Luo, Y.; Tang, C.X.; Xu, J.M. Plant material and its biochar differ in their effects on nitrogen mineralization and nitrification in a subtropical forest soil. Sci. Total Environ. 2021, 763, 143048. [Google Scholar] [CrossRef]
- Guo, K.Y.; Zhao, Y.Z.; Liu, Y.; Chen, J.H.; Wu, Q.F.; Ruan, Y.F.; Li, S.H.; Shi, J.; Zhao, L.; Sun, X.; et al. Pyrolysis temperature of biochar affects ecoenzymatic stoichiometry and microbial nutrient-use efficiency in a bamboo forest soil. Geoderma 2020, 363, 114162. [Google Scholar] [CrossRef]
- Sherman, L.; Coleman, M.D. Forest soil respiration and exoenzyme activity in western North America following thinning, residue removal for biofuel production, and compensatory soil amendments. GCB Bioenergy 2020, 12, 223–236. [Google Scholar] [CrossRef] [Green Version]
- Ventura, M.; Alberti, G.; Panzacchi, P.; Vedove, G.D.; Miglietta, F.; Tonon, G. Biochar mineralization and priming effect in a poplar short rotation coppice from a 3-year field experiment. Biol. Fertil. Soils 2019, 55, 67–78. [Google Scholar] [CrossRef]
- Janzen, H.; Janzen, D.; Gregorich, E. The ‘soil health’ metaphor: Illuminating or illusory? Soil Biol. Biochem. 2021, 159, 108167. [Google Scholar] [CrossRef]
- Martins, N.P.; Fuchslueger, L.; Fleischer, K.; Andersen, K.M.; Assis, R.L.; Baccaro, F.B.; Camargo, P.B.; Cordeiro, A.L.; Grandis, A.; Hartley, I.P.; et al. Fine roots stimulate nutrient release during early stages of leaf litter decomposition in a Central Amazon rainforest. Plant Soil 2021, 469, 287–303. [Google Scholar] [CrossRef]
- Kong, J.Q.; He, Z.B.; Chen, L.F.; Zhang, S.K.; Yang, R.; Du, J. Elevational variability in and controls on the temperature sensitivity of soil organic matter decomposition in alpine forests. Ecosphere 2022, 13, e4010. [Google Scholar] [CrossRef]
- Gross, D.C.; Bork, W.E.; Carlyle, N.C.; Chang, X.S. Biochar and its manure-based feedstock have divergent effects on soil organic carbon and greenhouse gas emissions in croplands. Sci. Total Environ. 2022, 806, 151337. [Google Scholar] [CrossRef]
- Miao, Y.C.; Li, J.J.; Li, Y.; Niu, Y.H.; He, T.H.; Liu, D.Y.; Ding, W.X. Long-term compost amendment sapurs cellulose decomposition by driving shifts in fungal community composition and promoting fungal diversity and phylogenetic relatedness. mBio 2022, 13, e0032322. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.L.; Yu, B.; Wang, B.R.; George, T.S.; Yin, H.Q.; Xu, D.Y.; Li, D.C.; Song, A.L. Microbial mechanisms of the contrast residue decomposition and priming effect in soils with different organic and chemical fertilization histories. Soil Biol. Biochem. 2019, 135, 213–221. [Google Scholar] [CrossRef]
- Bei, S.K.; Li, X.; Kuyper, T.W.; Chadwick, D.R.; Zhang, J.L. Nitrogen availability mediates the priming effect of soil organic matter by preferentially altering the straw carbon-assimilating microbial community. Sci. Total Environ. 2022, 815, 152882. [Google Scholar] [CrossRef]
- Singh, G.; Mavi, M.S.; Choudhary, O.P.; Kaur, M.; Singh, B. Interaction of pyrolysed and un-pyrolysed organic materials enhances carbon accumulation in soil irrigated with water of variable electrical conductivity. Soil Tillage Res. 2022, 215, 105193. [Google Scholar] [CrossRef]
- Wang, W.W.; Page-Dumroese, D.S.; Jurgensen, M.; Miller, C.; Walitalo, J.; Chen, X.; Liu, Y. Restoration thinning impacts surface and belowground wood decomposition. For. Ecol. Manag. 2019, 449, 117451. [Google Scholar] [CrossRef]
- Ren, H.; Warnock, D.D.; Tiemann, L.K.; Quigley, K.; Miesel, J.R. Evaluating foliar characteristics as early indicators of plant response to biochar amendments. For. Ecol. Manag. 2021, 489, 119047. [Google Scholar] [CrossRef]
- Adams, M.B.; Jurgensen, M.; Palik, B.; Miller, C.; Page-Dumroese, D.S. Wood stake decomposition twenty years after organic matter removal at the Lake States LTSP sites. For. Ecol. Manag. 2021, 496, 119456. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Jurgensen, M.F.; Miller, C.A.; Busse, M.D.; Curran, M.P.; Terry, T.A.; Tirocke, J.M.; Archuleta, J.G.; Murray, M. Decomposition of wood stakes in the Pacific Northwest after soil compaction and organic matter removal. For. Ecol. Manag. 2021, 494, 119362. [Google Scholar] [CrossRef]
- Wang, W.W.; Lindner, D.L.; Jusino, M.A.; Page-Dumroese, D.S.; Palmer, J.M.; Banik, M.T.; Jurgensen, M.; Draeger, K.; Liu, Y. Wood-colonizing fungal community response to forest restoration thinnings in a Pinus tabuliformis plantation in northern China. For. Ecol. Manag. 2020, 476, 118459. [Google Scholar] [CrossRef]
- Ji, M.Y.; Wang, X.X.; Usman, M.; Liu, F.H.; Dan, Y.T.; Zhou, L.; Campanaro, S.; Luo, G.; Sang, W.J. Effects of different feedstocks-based biochar on soil remediation: A review. Environ. Pollut. 2022, 294, 118655. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Jurgensen, M.F.; Miller, C.A.; Pickens, J.B.; Tirocke, J.M. Wildfire alters belowground and surface wood decomposition on two national forests in Montana, USA. Int. J. Wildland Fire 2019, 28, 456. [Google Scholar] [CrossRef]
- Jurgensen, M.F.; Miller, C.A.; Page-Dumroese, D.S. Wood decomposition after an aerial application of hydromulch following wildfire in a southern California chaparral shrubland. Front. For. Glob. Chang. 2020, 3, 93. [Google Scholar] [CrossRef]
- Risch, A.C.; Page-Dumroese, D.S.; Schweiger, A.K.; Beattie, J.R.; Curran, M.P.; Finér, L.; Hyslop, M.D.; Liu, Y.; Schütz, M.; Terry, T.A.; et al. Controls of initial wood decomposition on and in forest soils using standard material. Front. For. Glob. Chang. 2022, 5, 829810. [Google Scholar] [CrossRef]
- Wang, W.W.; Page-Dumroese, D.S.; Jurgensen, M.; Tirocke, J.; Liu, Y. Effect of forest thinning and wood quality on the short-term wood decomposition rate in a Pinus tabuliformis plantation. J. Plant Res. 2018, 131, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Wilke, B.-M. Determination of chemical and physical soil properties. In Manual for Soil Analysis–Monitoring and Cssessing Soil Bioremediation; Varma, A., Ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2005; pp. 47–65. [Google Scholar] [CrossRef]
- Walkley, A.J.; Black, I.A. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agro-Chemistrical Analysis; Agriculture Press: Beijing, China, 2001. (In Chinese) [Google Scholar]
- Thomas, R.L.; Sheard, R.W.; Moyer, J.R. Comparison of conventional and automated procedures for nitrogen, phosphorus, and potassium analysis of plant material using a single digestion. Agron. J. 1967, 59, 240–243. [Google Scholar] [CrossRef]
- Taniike, T.; Chammingkwan, P.; Thang, V.Q.; Funako, T.; Terano, M. Validation of BET specific surface area for heterogeneous Ziegler-Natta catalysts based on alpha(s)-plot. Appl. Catal. A Gen. 2012, 437, 24–27. [Google Scholar] [CrossRef]
- Pilcer, G.; Vanderbist, F.; Amighi, K. Correlations between cascade impactor analysis and laser diffraction techniques for the determination of the particle size of aerosolised powder formulations. Int. J. Pharm. 2008, 358, 75–81. [Google Scholar] [CrossRef]
- Mulvaney, R.L. Nitrogen-Inorganic Forms. In Methods of Soil Analysis; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; American Society of Agronomy, Incorporation: Madison, WI, USA, 1996; pp. 1123–1184. [Google Scholar] [CrossRef] [Green Version]
- Guan, S.Y. Soil Enzyme and Its Research Approaches; China Agriculture Press: Beijing, China, 1986. (In Chinese) [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing. In R Core Team. 2021. Available online: http://www.R-project.org/ (accessed on 10 August 2021).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V.; Buerkner, P.; Herve, M.; Love, J.; Miguez, F.; Riebl, H.; Singmann, H. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. In R Package Version 1.7.5. 2022. Available online: https://github.com/rvlenth/emmeans (accessed on 1 September 2022).
- Hollander, M.; Wolfe, D.A. Nonparametric Statistical Methods; Wiley: New York, NY, USA, 1973. [Google Scholar]
- Press, W.H.; Flannery, B.P.; Teukolsky, S.A.; Vetterling, W.T. Numerical Recipes in C; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Piaszczyk, W.; Lasota, J.; Błońska, E.; Foremnik, K. How habitat moisture condition affects the decomposition of fine woody debris from different species. Catena 2022, 208, 105765. [Google Scholar] [CrossRef]
- Hu, Z.H.; Michaletz, S.T.; Johnson, D.J.; McDowell, N.G.; Huang, Z.Q.; Zhou, X.H.; Xu, C.G. Traits drive global wood decomposition rates more than climate. Glob. Chang. Biol. 2018, 24, 5259–5269. [Google Scholar] [CrossRef]
- Somerville, P.D.; Farrell, C.; May, P.B.; Livesley, S.J. Biochar and compost equally improve urban soil physical and biological properties and tree growth, with no added benefit in combination. Sci. Total Environ. 2020, 706, 135736. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Bao, H.Y.; Xing, W.J.; Yang, J.; Liu, R.F.; Wang, X.; Lv, L.H.; Tong, X.G.; Wu, F.Y. Succession of fungal dynamics and their influence on physicochemical parameters during pig manure composting employing with pine leaf biochar. Bioresour. Technol. 2020, 297, 122377. [Google Scholar] [CrossRef]
- Baietto, A.; Hirigoyen, A.; Hernández, J.; del Pino, A. Comparative dynamics of nutrient release through litter decomposition in Eucalyptus grandis Hill ex Maiden and Pinus taeda L. stands. Forests 2021, 12, 1227. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Ascher-Jenull, J.; Bardelli, T.; Fornasier, F.; Fravolini, G.; Arfaioli, P.; Ceccherini, M.T.; Pietramellara, G.; Lamorski, K.; Sławiński, C.; et al. Physico-chemical and microbiological evidence of exposure effects on Picea abies – Coarse woody debris at different stages of decay. For. Ecol. Manag. 2017, 391, 376–389. [Google Scholar] [CrossRef]
- Sterner, R.; Elser, J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar] [CrossRef]
- Romashkin, I.; Shorohova, E.; Kapitsa, E.; Galibina, N.; Nikerova, K. Substrate quality regulates density loss, cellulose degradation and nitrogen dynamics in downed woody debris in a boreal forest. For. Ecol. Manag. 2021, 491, 119143. [Google Scholar] [CrossRef]
- Palviainen, M.; Finér, L.; Laiho, R.; Shorohova, E.; Kapitsa, E.; Vanha-Majamaa, I. Carbon and nitrogen release from decomposing Scots pine, Norway spruce and silver birch stumps. For. Ecol. Manag. 2010, 259, 390–398. [Google Scholar] [CrossRef]
- Jurgensen, M.; Reed, D.; Page-Dumroese, D.; Laks, P.; Collins, A.; Mroz, G.; Degórski, M. Wood strength loss as a measure of decomposition in northern forest mineral soil. Eur. J. Soil Biol. 2006, 42, 23–31. [Google Scholar] [CrossRef]
- Berg, B.; Staaf, H. Leaching, accumulation and release of nitrogen in decomposing forest litter. Ecol. Bull. 1981, 33, 163–178. [Google Scholar]
- Li, X.N.; Wang, T.; Chang, S.X.; Jiang, X.; Song, Y. Biochar increases soil microbial biomass but has variable effects on microbial diversity: A meta-analysis. Sci. Total Environ. 2020, 749, 141593. [Google Scholar] [CrossRef]
- Han, Z.Q.; Xu, P.S.; Li, Z.T.; Lin, H.Y.; Zhu, C.; Wang, J.Y.; Zou, J.W. Microbial diversity and the abundance of keystone species drive the response of soil multifunctionality to organic substitution and biochar amendment in a tea plantation. GCB Bioenergy 2022, 14, 481–495. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Schencker, J.R.; Wild, B.; Leitner, S.; Hofhansl, F.; Chl, A.B.; Mmerle, I.H.; Frank, A.H.; Fuchslueger, L.; et al. Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter. Ecology 2012, 93, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.W.D.; Soares, E.M.B.; Oliveira, F.C.C.; Silva, I.R.; Dungait, J.A.J.; Souza, I.F.; Vergütz, L. Nutrient release from decomposing Eucalyptus harvest residues following simulated management practices in multiple sites in Brazil. For. Ecol. Manag. 2016, 370, 1–11. [Google Scholar] [CrossRef]
- Jaroszewicz, B.; Cholewińska, O.; Chećko, E.; Wrzosek, M. Predictors of diversity of deadwood-dwelling macrofungi in a European natural forest. For. Ecol. Manag. 2021, 490, 119123. [Google Scholar] [CrossRef]
- Bormann, H.; Klaassen, K. Seasonal and land use dependent variability of soil hydraulic and soil hydrological properties of two Northern German soils. Geoderma 2008, 145, 295–302. [Google Scholar] [CrossRef]
- Razzaghi, F.; Obour, P.B.; Arthur, E. Does biochar improve soil water retention? A systematic review and meta-analysis. Geoderma 2020, 361, 114055. [Google Scholar] [CrossRef]
- Thomas, S.C.; Gale, N. Biochar and forest restoration: A review and meta-analysis of tree growth responses. New For. 2015, 46, 931–946. [Google Scholar] [CrossRef]
- Tammeorg, P.; Simojoki, A.; Mäkelä, P.; Stoddard, F.L.; Alakukkuc, L.; Heleniusa, J. Short-term effects of biochar on soil properties and wheat yield formation with meat bone meal and inorganic fertiliser on a boreal loamy sand. Agric. Ecosyst. Environ. 2014, 191, 108–116. [Google Scholar] [CrossRef]
- Espinosa, N.J.; Moore, D.J.P.; Rasmussen, C.; Fehmi, J.S.; Gallery, R.E. Woodchip and biochar amendments differentially influence microbial responses, but do not enhance plant recovery in disturbed semiarid soils. Restor. Ecol. 2020, 28, S381–S392. [Google Scholar] [CrossRef]
- Lehmann, J.; Silva Junior, J.; Steiner, C.; Nehls, T.; Zech, W.; Glaser, B. Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: Fertilizer, manure and charcoal amendments. Plant Soil 2003, 249, 343–357. [Google Scholar] [CrossRef]
- Frankenberger, W.; Johanson, J. Factors affecting invertase activity in soils. Plant Soil 1983, 74, 313–323. [Google Scholar] [CrossRef]
- Sarma, B.; Borkotoki, B.; Narzari, R.; Kataki, R.; Gogoi, N. Organic amendments: Effect on carbon mineralization and crop productivity in acidic soil. J. Clean. Prod. 2017, 152, 157–166. [Google Scholar] [CrossRef]
- Sekaran, U.; Sandhu, S.S.; Qiu, Y.Y.; Kumar, S.; Hernandez, J.L.G. Biochar and manure addition influenced soil microbial community structure and enzymatic activities at eroded and depositional landscape positions. Land Degrad. Dev. 2019, 31, 894–908. [Google Scholar] [CrossRef]
- Singh, R.; Nye, P. The effect of soil pH and high urea concentrations on urease activity in soil. J. Soil Sci. 2006, 35, 519–527. [Google Scholar] [CrossRef]
- Li, X.N.; Song, Y.; Wang, F.; Bian, Y.R.; Jiang, X. Combined effects of maize straw biochar and oxalic acid on the dissipation of polycyclic aromatic hydrocarbons and microbial community structures in soil: A mechanistic study. J. Hazard. Mater. 2019, 364, 325–331. [Google Scholar] [CrossRef]
- Kunito, T.; Akagi, Y.; Park, H.-D.; Toda, H. Influences of nitrogen and phosphorus addition on polyphenol oxidase activity in a forested Andisol. Eur. J. For. Res. 2009, 128, 361–366. [Google Scholar] [CrossRef]
- Amonette, J.; Blanco-Canqui, H.; Hassebrook, C.; Laird, D.; Lal, R.; Lehmann, J.; Page-Dumroese, D. Integrated biochar research: A roadmap. J. Soil Water Conserv. 2021, 76, 24A–29A. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Mineral Soil Depth | Maize Straw Biochar | Pig Manure | |
|---|---|---|---|---|
| 0–30 cm | 30–60 cm | |||
| Total N (mg g−1) | 2.2 | 1.6 | 10.7 | 14.6 |
| Total P (mg g−1) | 1.4 | 1.8 | 1.6 | 19.5 |
| Total K (mg g−1) | 8.3 | 7.5 | 2.6 | 26.8 |
| pH | 8.2 | 7.5 | 8.8 | 7.9 |
| Organic matter content (g kg−1) | 7 | 6 | - | 450 |
| C (%) | 1.1 | 1.0 | 48 | - |
| C:N | 5.0 | 6.1 | 44 | - |
| Hydrogen content (g kg−1) | - | - | 28 | - |
| Specific surface area (m2 g−1) | - | - | 5.0 | - |
| Pore volume (cm3 g−1) | - | - | 6.9 × 10−3 | - |
| Pore size (nm) | - | - | 5.6 | - |
| Total ash content (%) | - | - | 44 | - |
| Species | C (%) | N (mg g−1) | C:N | P (mg g−1) | K (mg g−1) | C:P | N:P | Lignin (%) | Carbohydrate (%) | Lignin:N |
|---|---|---|---|---|---|---|---|---|---|---|
| Poplar | 46.75 ± 0.54 b | 1.4 ± 0.4 a | 354.64 ± 97.84 c | 0.05 ± 0.02 ab | 0.59 ± 0.11 a | 12182.77 ± 2504.18 a | 28.81 ± 4.44 a | - | - | - |
| Aspen | 46.40 ± 0.51 b | 1.10 ± 0.10 b | 421.81 ± 37.10 b | 0.06 ± 0.02 a | 0.62 ± 0.13 a | 7158.14 ± 1782.18 b | 18.95 ± 1.24 b | 20.64 ± 1.08 b | 68.66 ± 3.38 a | 189.50 ± 19.59 b |
| Pine | 48.62 ± 1.05 a | 0.8 ± 0.2 c | 631.65 ± 110.29 a | 0.03 ± 0.01 b | 0.29 ± 0.07 b | 14646.21 ± 2756.08 a | 27.86 ± 1.76 ab | 31.09 ± 2.09 a | 59.60 ± 2.64 b | 403.72 ± 61.58 a |
| p values | 0.0000 *** | 0.0000 *** | 0.0000 *** | 0.0043 ** | 0.0042 ** | 0.0016 ** | 0.0146 * | 0.0000 *** | 0.0000 *** | 0.0000 *** |
| Location and Species | Wood Stake | Soil Properties | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Moisture Content | N Loss | P Loss | K Loss | Total N | Total P | Total K | Soil Organic Carbon | Available N | C:N | Catalase | PPO | Urease | Invertase | ||
| Pearson Correlation Coefficients (r) | |||||||||||||||
| Surface | Mass loss | 0.32 *** | 0.02 | 0.28 *** | 0.09 | 0.04 | −0.15 | 0.02 | 0.15 | 0.14 | 0.12 | 0.08 | 0.09 | 0.03 | 0.18 * |
| Poplar | N loss | 0.25 ** | −0.01 | 0.09 | 0.20 * | 0.04 | −0.06 | 0.18 * | −0.07 | 0.05 | −0.07 | ||||
| P loss | −0.03 | −0.06 | 0.05 | 0.00 | 0.01 | 0.05 | 0.02 | 0.09 | 0.08 | 0.06 | |||||
| K loss | 0.17 * | −0.13 | 0.15 | 0.16 | 0.07 | 0.00 | 0.17 * | 0.01 | 0.10 | −0.01 | |||||
| Aspen | Mass loss | 0.28 *** | −0.40 *** | −0.41 *** | −0.20 * | −0.08 | 0.04 | 0.06 | 0.08 | 0.14 | 0.18 * | ||||
| N loss | −0.14 | 0.10 | −0.18 * | −0.16 | 0.02 | −0.03 | 0.07 | 0.11 | −0.06 | 0.03 | |||||
| P loss | 0.09 | 0.11 | −0.16 | −0.06 | 0.00 | −0.18 * | −0.08 | −0.01 | −0.08 | 0.07 | |||||
| K loss | 0.15 | −0.01 | −0.05 | 0.03 | −0.10 | −0.13 | −0.01 | −0.20 * | −0.06 | −0.09 | |||||
| −0.04 | −0.11 | −0.03 | −0.08 | ||||||||||||
| Pine | Mass loss | 0.12 | −0.27** | −0.01 | −0.09 | 0.02 | 0.02 | 0.07 | −0.02 | 0.04 | −0.05 | ||||
| N loss | 0.00 | 0.02 | −0.07 | −0.01 | 0.03 | −0.02 | 0.12 | 0.08 | 0.06 | −0.09 | |||||
| P loss | −0.04 | 0.07 | −0.02 | 0.03 | −0.25 ** | 0.09 | 0.04 | −0.02 | 0.05 | 0.15 | |||||
| K loss | −0.17 * | 0.05 | −0.10 | −0.14 | −0.03 | 0.03 | −0.07 | −0.07 | 0.05 | −0.06 | |||||
| −0.09 | −0.04 | −0.04 | −0.03 | ||||||||||||
| Mineral | Mass loss | 0.16 | 0.30 *** | 0.18 * | 0.13 | 0.02 | −0.09 | −0.10 | 0.05 | 0.13 | 0.02 | ||||
| Poplar | N loss | 0.08 | −0.11 | 0.02 | 0.17 | 0.15 | 0.07 | 0.06 | 0.05 | −0.05 | 0.06 | ||||
| P loss | 0.12 | 0.03 | 0.05 | 0.06 | −0.0 | −0.08 | 0.18 * | 0.04 | 0.18 * | 0.10 | |||||
| K loss | 0.01 | −0.03 | 0.06 | −0.05 | 0.01 | −0.07 | 0.17 | −0.01 | 0.14 | 0.03 | |||||
| 0.11 | 0.05 | 0.13 | 0.02 | ||||||||||||
| Aspen | Mass loss | 0.31 *** | −0.16 | 0.13 | −0.06 | −0.10 | 0.03 | −0.01 | −0.01 | 0.03 | 0.09 | ||||
| N loss | −0.08 | −0.07 | 0.07 | 0.02 | 0.01 | 0.12 | −0.05 | 0.17 | −0.09 | 0.02 | |||||
| P loss | −0.15 | −0.01 | −0.09 | −0.05 | 0.01 | 0.11 | −0.08 | 0.10 | −0.06 | 0.07 | |||||
| K loss | −0.01 | 0.02 | −0.07 | 0.00 | −0.01 | 0.01 | −0.11 | 0.13 | −0.09 | 0.16 | |||||
| −0.09 | 0.10 | −0.11 | 0.05 | ||||||||||||
| Pine | Mass loss | 0.35 *** | −0.18 * | −0.26 ** | −0.16 | −0.07 | 0.10 | −0.04 | −0.04 | −0.04 | 0.04 | ||||
| N loss | −0.10 | −0.11 | 0.05 | −0.01 | 0.09 | 0.09 | −0.04 | −0.04 | 0.01 | 0.06 | |||||
| P loss | 0.03 | −0.05 | −0.02 | −0.10 | 0.12 | −0.13 | 0.07 | 0.08 | 0.02 | 0.13 | |||||
| K loss | −0.09 | −0.01 | 0.06 | −0.05 | −0.01 | 0.04 | 0.05 | −0.04 | −0.01 | −0.04 | |||||
| Treatments | Soil Hydraulic Properties | ||||
|---|---|---|---|---|---|
| Bulk Density (Mg m−3) | Maximum Water Holding Capacity (g kg−1) | Capillary Capacity (g kg−1) | Field Capacity (g kg−1) | Lower Limit of Optimum Moisture Content (g kg−1) | |
| M0B0 | 1.20 ± 0.03 b | 604 ± 29 a | 585 ± 28 a | 537 ± 28 ab | 376 ± 19 ab |
| M0B2.5 | 1.17 ± 0.03 b | 626 ± 29 a | 607 ± 28 a | 566 ± 28 a | 396 ± 20 a |
| M0B10 | 1.19 ± 0.02 b | 607 ± 22 a | 590 ± 22 a | 547 ± 21 a | 383 ± 15 a |
| M3B0 | 1.27 ± 0.03 a | 512 ± 29 b | 496 ± 28 b | 458 ± 28 b | 321 ± 19 b |
| M3B2.5 | 1.17 ± 0.02 b | 619 ± 22 a | 599 ± 22 a | 556 ± 21 a | 389 ± 15 a |
| M3B10 | 1.17 ± 0.02 b | 621 ± 22 a | 604 ± 22 a | 560 ± 21 a | 392 ± 15 a |
| M9B0 | 1.14 ± 0.03 b | 644 ± 29 a | 626 ± 28 a | 583 ± 28 a | 408 ± 20 a |
| M9B2.5 | 1.18 ± 0.02 b | 588 ± 22 a | 572 ± 22 a | 530 ± 21 ab | 371 ± 15 ab |
| M9B10 | 1.15 ± 0.03 b | 620 ± 29 a | 602 ± 28 a | 557 ± 28 a | 390 ± 19 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, R.; Liu, Y.; Page-Dumroese, D.S.; Dumroese, R.K.; Wang, K. Enhancing Soil Quality of Short Rotation Forest Operations Using Biochar and Manure. Forests 2022, 13, 2090. https://doi.org/10.3390/f13122090
Zhao R, Liu Y, Page-Dumroese DS, Dumroese RK, Wang K. Enhancing Soil Quality of Short Rotation Forest Operations Using Biochar and Manure. Forests. 2022; 13(12):2090. https://doi.org/10.3390/f13122090
Chicago/Turabian StyleZhao, Ruirui, Yong Liu, Deborah S. Page-Dumroese, R. Kasten Dumroese, and Kai Wang. 2022. "Enhancing Soil Quality of Short Rotation Forest Operations Using Biochar and Manure" Forests 13, no. 12: 2090. https://doi.org/10.3390/f13122090