


Response of Soil Microbial Communities and Functions to Long-Term Tea (Camellia sinensis L.) Planting in a Subtropical Region
Abstract
:1. Introduction
2. Materials and Methods
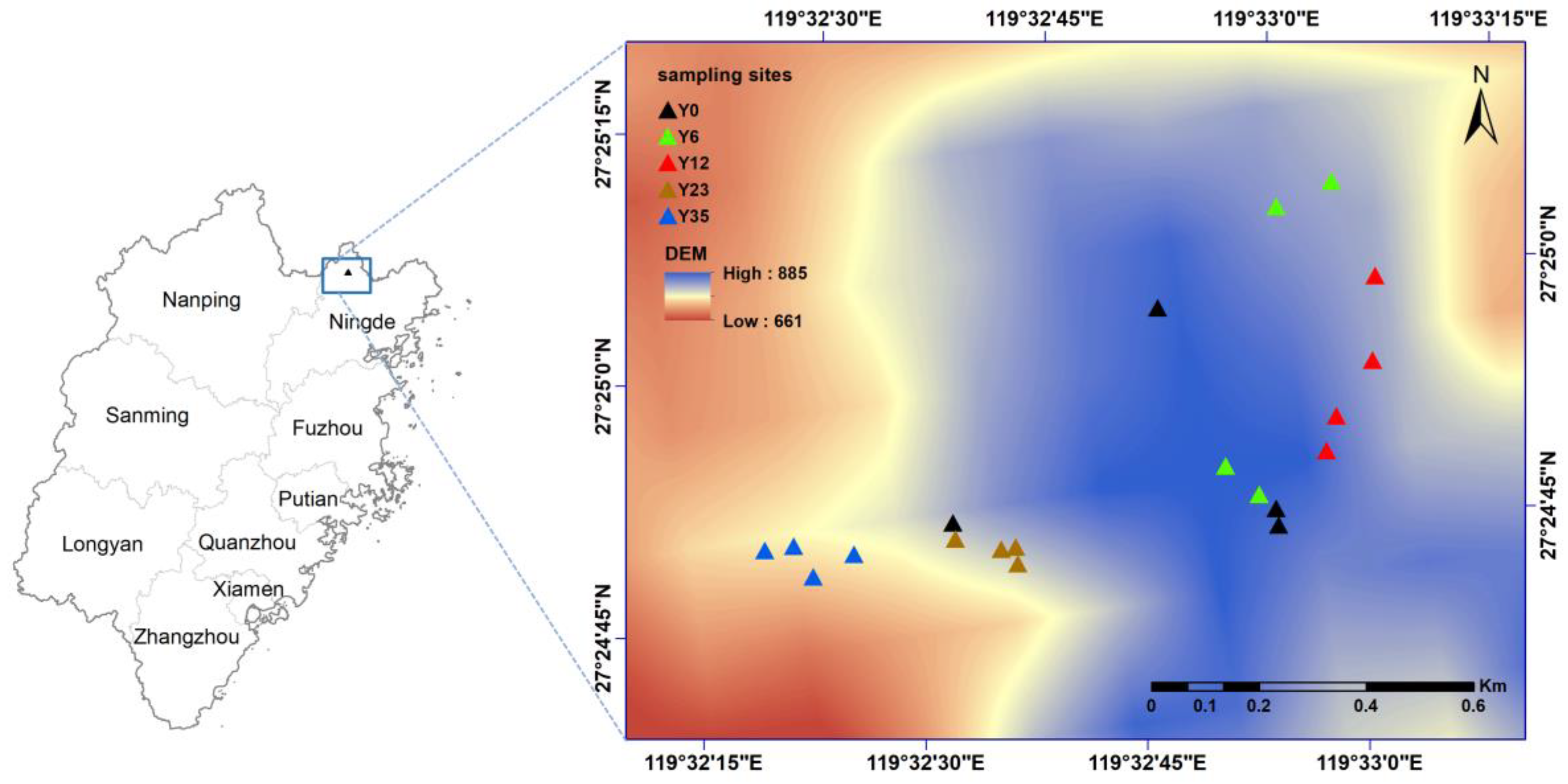
2.1. Study Site
2.2. Soil Physiochemical Analyses
2.3. DNA Extraction, Amplification and Sequencing
2.4. Bioinformatic Processing and Statistical Analysis
3. Results
3.1. Changes in Soil Physicochemical Properties
3.2. Alpha Diversity
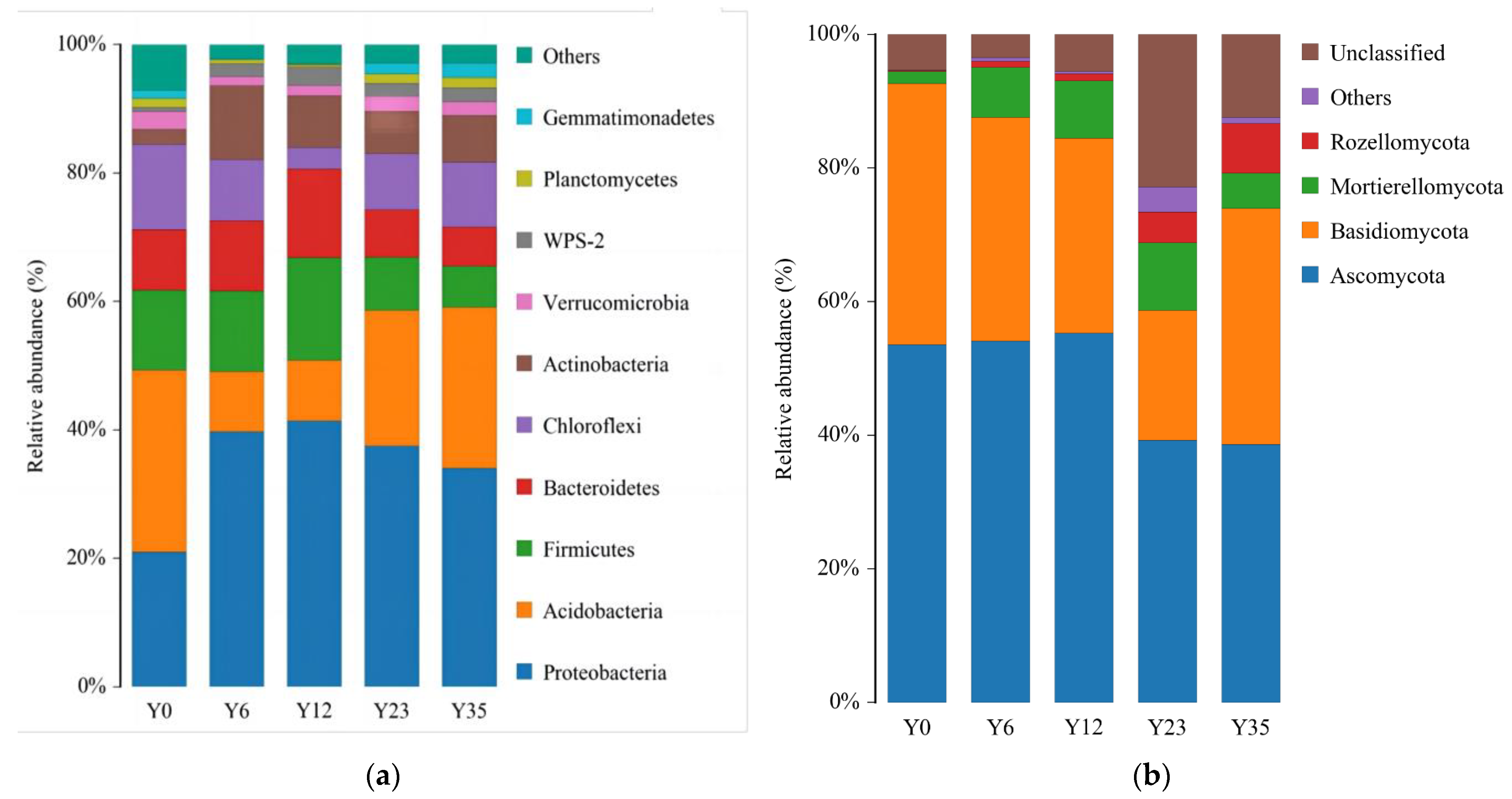
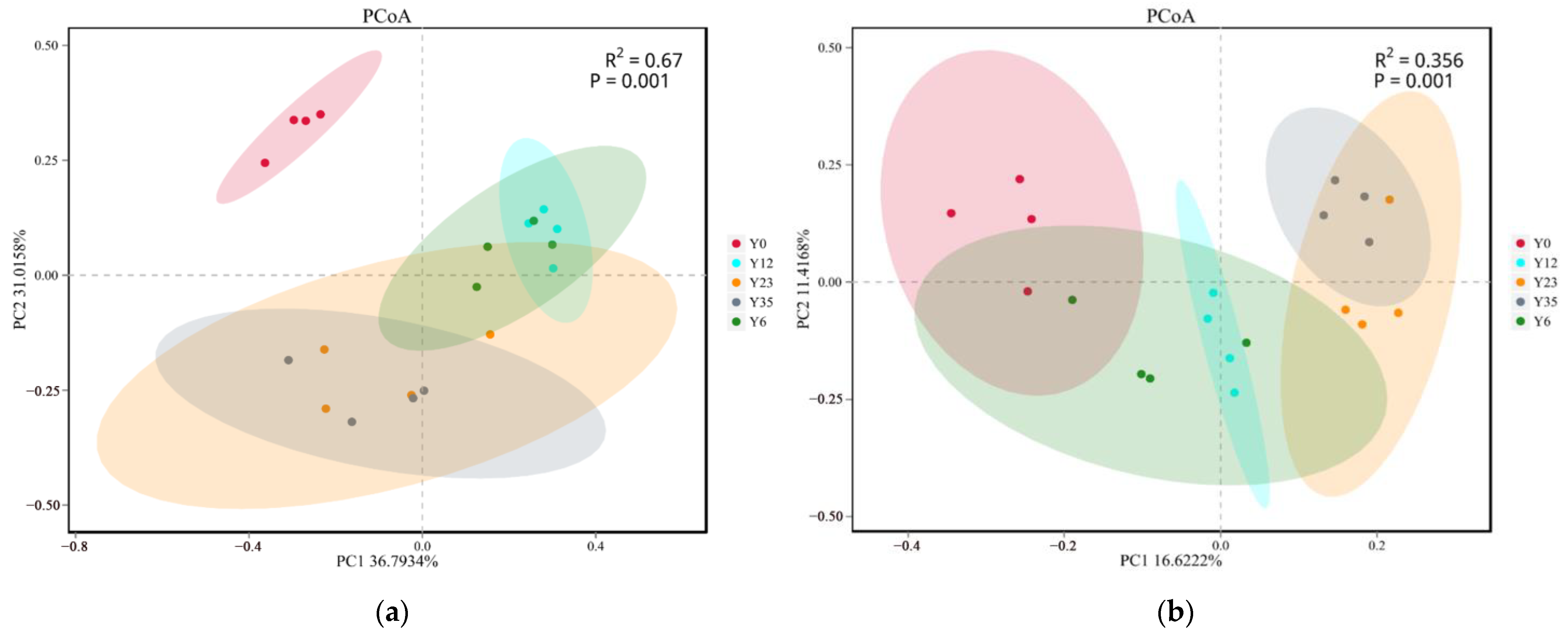
3.3. Structures of Soil Microbial Communities at Different Tea Planting Ages
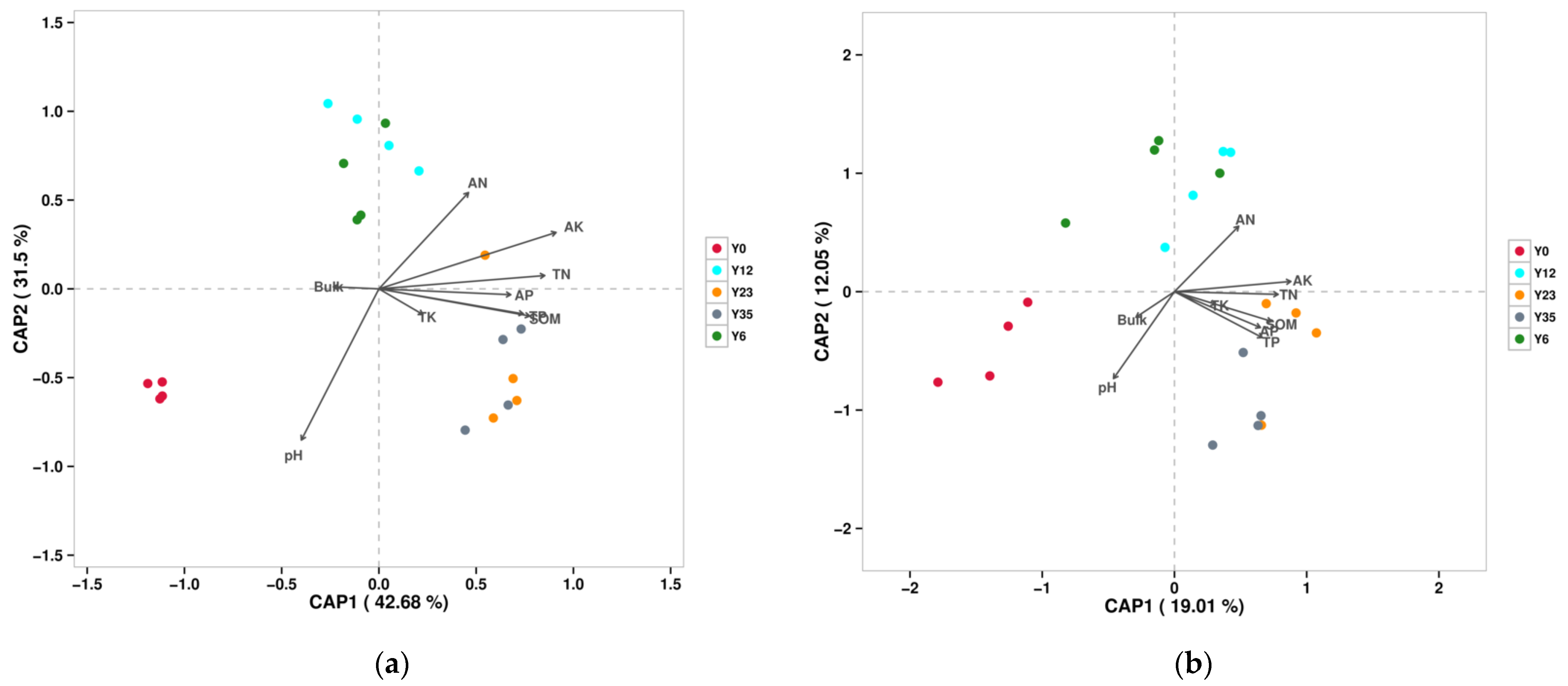
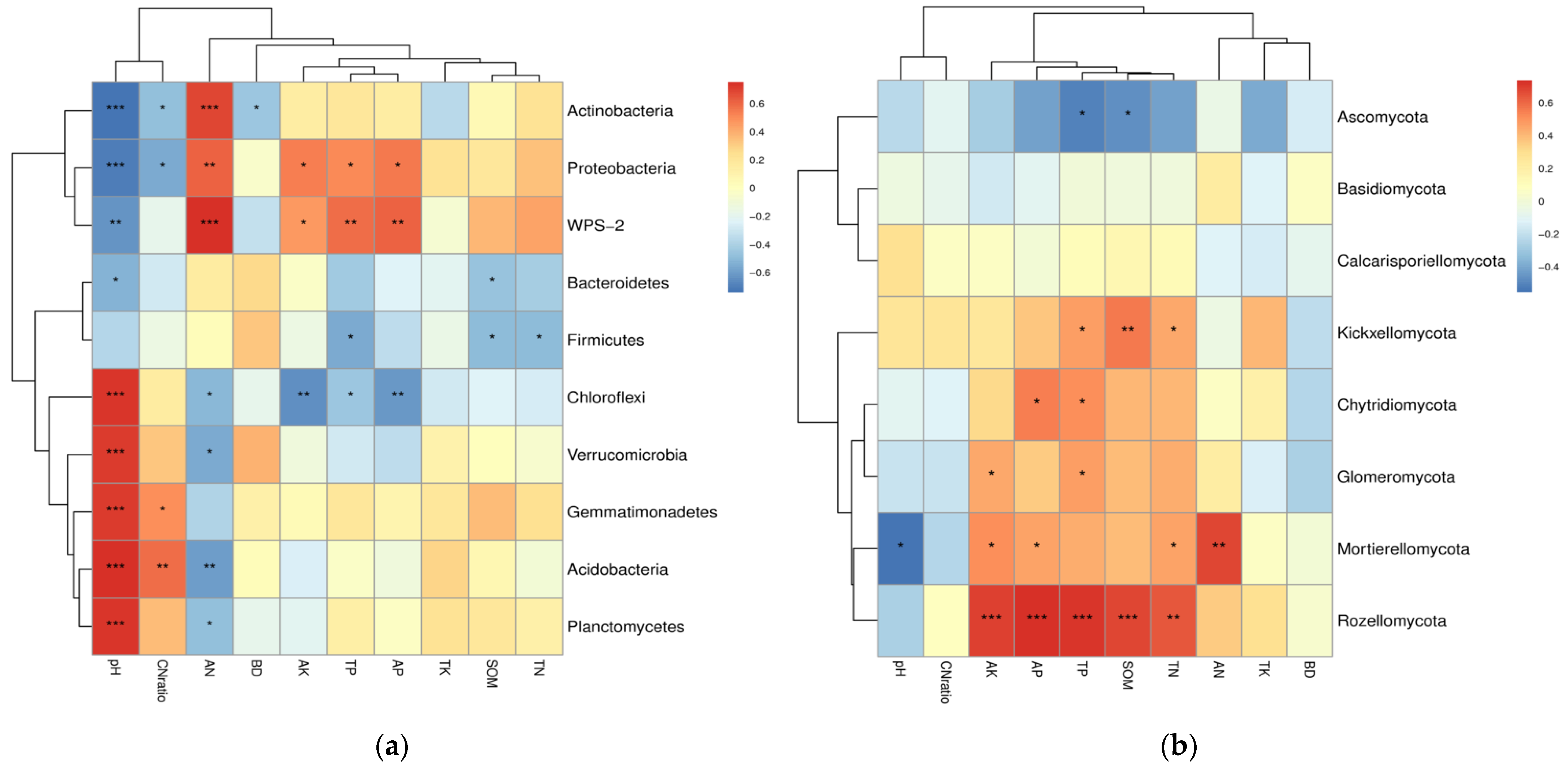
3.4. Correlations between Soil Microbial Community and Environmental Factors
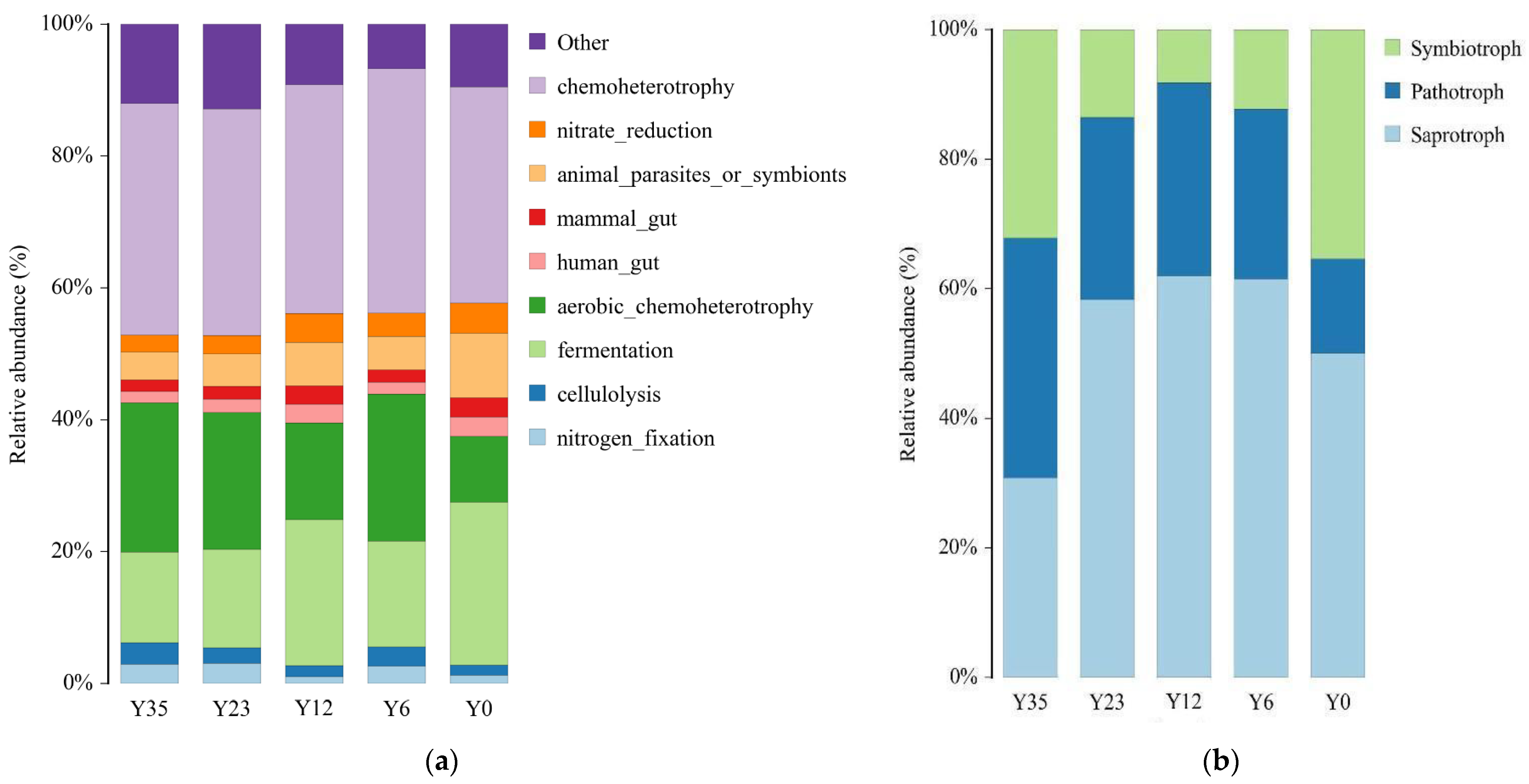
3.5. Microbial Function Potential
4. Discussion
4.1. Long-Term Tea Plantation Increased Soil Bacterial Richness and Diversity but Decreased Fungal Richness
4.2. Different Tea Planting Ages Shift the Soil Microbial Community Structure
4.3. Potential Microbial Functional Variations Influenced by Tea Planting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arafat, Y.; Ud Din, I.; Tayyab, M.; Jiang, Y.; Chen, T.; Cai, Z.; Zhao, H.; Lin, X.; Lin, W.; Lin, S. Soil Sickness in Aged Tea Plantation Is Associated With a Shift in Microbial Communities as a Result of Plant Polyphenol Accumulation in the Tea Gardens. Front. Plant Sci. 2020, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yao, X.; Ye, S. Soil Aggregate-Related Organic Carbon and Relevant Enzyme Activities as Affected by Tea (Camellia Sinensis L.) Planting Age in Hilly Region of Southern Guangxi, China. Appl. Soil Ecol. 2020, 150, 103444. [Google Scholar] [CrossRef]
- Sidle, R.C.; Ziegler, A.D.; Negishi, J.N.; Nik, A.R.; Siew, R.; Turkelboom, F. Erosion Processes in Steep Terrain—Truths, Myths, and Uncertainties Related to Forest Management in Southeast Asia. For. Ecol. Manage. 2006, 224, 199–225. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Zhu, T.; Müller, C.; Cai, Z. ScienceDirect Effect of Orchard Age on Soil Nitrogen Transformation in Subtropical China and Implications. J. Environ. Sci. 2015, 34, 10–19. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Chapman, S.J.; Yao, H.; Zheng, N.; Müller, C. Tea Plantation Affects Soil Nitrogen Transformations in Subtropical China. J. Soils Sediments 2021, 21, 441–451. [Google Scholar] [CrossRef]
- Kamaa, M.; Mburu, H.; Blanchart, E.; Chibole, L.; Chotte, J.-L.; Kibunja, C.; Lesueur, D. Effects of Organic and Inorganic Fertilization on Soil Bacterial and Fungal Microbial Diversity in the Kabete Long-Term Trial, Kenya. Biol. Fertil. Soils 2011, 47, 315–321. [Google Scholar] [CrossRef]
- Chen, Y.; Deng, Y.; Ding, J.; Hu, H.; Xu, T.; Li, F.; Yang, G.; Yang, Y. Distinct Microbial Communities in the Active and Permafrost Layers on the Tibetan Plateau. Mol. Ecol. 2017, 26, 6608–6620. [Google Scholar] [CrossRef]
- Wu, X.; Xu, H.; Tuo, D.; Wang, C.; Fu, B.; Lv, Y.; Liu, G. Land Use Change and Stand Age Regulate Soil Respiration by Influencing Soil Substrate Supply and Microbial Community. Geoderma 2020, 359, 113991. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an Ecological Classification of Soil Bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Sun, L.; Xun, W.; Huang, T.; Zhang, G.; Gao, J.; Ran, W.; Li, D.; Shen, Q.; Zhang, R. Alteration of the Soil Bacterial Community during Parent Material Maturation Driven by Different Fertilization Treatments. Soil Biol. Biochem. 2016, 96, 207–215. [Google Scholar] [CrossRef]
- Du, C.; Xu, C.; Jian, J.; He, W.; Hou, L.; Geng, Z. Seasonal Dynamics of Bacterial Communities in a Betula Albosinensis Forest. Eur. J. Soil Sci. 2018, 69, 666–674. [Google Scholar] [CrossRef]
- Xu, Y.; Du, A.; Wang, Z.; Zhu, W.; Li, C.; Wu, L. Effects of Different Rotation Periods of Eucalyptus Plantations on Soil Physiochemical Properties, Enzyme Activities, Microbial Biomass and Microbial Community Structure and Diversity. For. Ecol. Manage. 2020, 456, 117683. [Google Scholar] [CrossRef]
- Liu, D.; Huang, Y.; Sun, H.; An, S. The Restoration Age of Robinia Pseudoacacia Plantation Impacts Soil Microbial Biomass and Microbial Community Structure in the Loess Plateau. Catena 2018, 165, 192–200. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, B.; Ma, Y.; Sun, H. Differences in Bacterial Community Structure and Potential Functions among Eucalyptus Plantations with Different Ages and Species of Trees. Appl. Soil Ecol. 2020, 149, 103515. [Google Scholar] [CrossRef]
- Dang, P.; Yu, X.; Le, H.; Liu, J.; Shen, Z.; Zhao, Z. Effects of Stand Age and Soil Properties on Soil Bacterial and Fungal Community Composition in Chinese Pine Plantations on the Loess Plateau. PLoS ONE 2017, 12, e0186501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Li, C.; Wang, S.; Zhou, B.; Mao, Y.; Rensing, C.; Xing, S. Influence of Biochar and Biochar-Based Fertilizer on Yield, Quality of Tea and Microbial Community in an Acid Tea Orchard Soil. Appl. Soil Ecol. 2021, 166, 104005. [Google Scholar] [CrossRef]
- Ni, K.; Liao, W.; Yi, X.; Niu, S.; Ma, L.; Shi, Y.; Zhang, Q.; Liu, M.; Ruan, J. Fertilization Status and Reduction Potential in Tea Gardens of China. J. Plant Nutr. Fer. 2019, 25, 421–432. [Google Scholar]
- Sae-Lee, N.; Kerdchoechuen, O.; Laohakunjit, N. Chemical Qualities and Phenolic Compounds of Assam Tea after Soil Drench Application of Selenium and Aluminium. Plant Soil 2012, 356, 381–393. [Google Scholar] [CrossRef]
- Wang, S.; Li, T.; Zheng, Z. Distribution of Microbial Biomass and Activity within Soil Aggregates as Affected by Tea Plantation Age. Catena 2017, 153, 1–8. [Google Scholar] [CrossRef]
- Bai, Y.; Zha, X.; Chen, S. Effects of the Vegetation Restoration Years on Soil Microbial Community Composition and Biomass in Degraded Lands in Changting County, China. J. For. Res. 2019, 31, 1295–1308. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; Wiley Online Library: Hoboken, NJ, USA, 1983; Volume 9, pp. 539–579. [Google Scholar]
- Lu, R.K. Soil Agricultural Chemical Analysis Methods; China Agricultural Science and Technology: Beijing, China, 2000; pp. 154–196. [Google Scholar]
- Huse, S.M.; Dethlefsen, L.; Huber, J.A.; Welch, D.M.; Relman, D.A.; Sogin, M.L. Exploring Microbial Diversity and Taxonomy Using SSU RRNA Hypervariable Tag Sequencing. PLoS Genet. 2008, 4, e1000255. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS Primers with Enhanced Specificity for Basidiomycetes-application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling Function and Taxonomy in the Global Ocean Microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An Open Annotation Tool for Parsing Fungal Community Datasets by Ecological Guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Wang, S.; Li, T.; Zheng, Z.; Chen, H.Y.H. Soil Aggregate-Associated Bacterial Metabolic Activity and Community Structure in Different Aged Tea Plantations. Sci. Total Environ. 2019, 654, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil Bacterial and Fungal Communities across a PH Gradient in an Arable Soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yan, M.; Lin, C.; Guo, B.; Ding, H.; Yu, J.; Peng, S.; Sveen, T.R.; Zhang, Y. Vegetation Restoration Types Affect Soil Bacterial Community Composition and Diversity in Degraded Lands in Subtropical of China. Restor. Ecol. 2022, 30, e13494. [Google Scholar] [CrossRef]
- De Gannes, V.; Bekele, I.; Dipchansingh, D.; Wuddivira, M.N.; De Cairies, S.; Boman, M.; Hickey, W.J. Microbial Community Structure and Function of Soil Following Ecosystem Conversion from Native Forests to Teak Plantation Forests. Front. Microbiol. 2016, 7, 1976. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Shen, W.; Yin, Y.; Zhang, J.; Cai, Z.; Zhong, W. Response of Soil Microbial Diversity to Land-Use Conversion of Natural Forests to Plantations in a Subtropical Mountainous Area of Southern China. Soil Sci. Plant Nutr. 2012, 58, 450–461. [Google Scholar] [CrossRef]
- Nie, S.; Lei, X.; Zhao, L.; Brookes, P.C.; Wang, F.; Chen, C.; Yang, W.; Xing, S. Fungal Communities and Functions Response to Long-Term Fertilization in Paddy Soils. Appl. Soil Ecol. 2018, 130, 251–258. [Google Scholar] [CrossRef]
- Husson, O. Redox Potential (Eh) and PH as Drivers of Soil/Plant/Microorganism Systems: A Transdisciplinary Overview Pointing to Integrative Opportunities for Agronomy. Plant Soil 2013, 362, 389–417. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.J.; Whiteley, A.S. The Bacterial Biogeography of British Soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Yu, Z.; Shi, Y.U.; Chu, H.; Jin, J.; Liu, X.; Wang, G. High Throughput Sequencing Analysis of Biogeographical Distribution of Bacterial Communities in the Black Soils of Northeast China. Soil Biol. Biochem. 2014, 70, 113–122. [Google Scholar] [CrossRef]
- Ji, L.; Ni, K.; Wu, Z.; Zhang, J.; Yi, X.; Yang, X.; Ling, N.; You, Z.; Guo, S.; Ruan, J. Effect of Organic Substitution Rates on Soil Quality and Fungal Community Composition in a Tea Plantation with Long-Term Fertilization. Biol. Fertil. Soils 2020, 56, 633–646. [Google Scholar] [CrossRef]
- Bai, Z.; Wu, X.; Lin, J.-J.; Xie, H.-T.; Yuan, H.-S.; Liang, C. Litter-, Soil-and C: N-Stoichiometry-Associated Shifts in Fungal Communities along a Subtropical Forest Succession. Catena 2019, 178, 350–358. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Puusepp, R.; Nilsson, R.H.; James, T.Y. Novel Soil-Inhabiting Clades Fill Gaps in the Fungal Tree of Life. Microbiome 2017, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Li, K.; Zhou, W.; Qiu, S.; Huang, S.; He, P. Changes in Soil Microbial Community, Enzyme Activities and Organic Matter Fractions under Long-Term Straw Return in North-Central China. Agric. Ecosyst. Environ. 2016, 216, 82–88. [Google Scholar] [CrossRef]
- Zhang, J.B.; Zhu, T.B.; Cai, Z.C.; Qin, S.W.; Müller, C. Effects of Long-term Repeated Mineral and Organic Fertilizer Applications on Soil Nitrogen Transformations. Eur. J. Soil Sci. 2012, 63, 75–85. [Google Scholar] [CrossRef]
- Wang, F.; Chen, Y.; Yu, X.; Yu, W.; You, Z.; Yang, Z. Response of the Soil Fungal Community and Its Function during the Conversion of Forestland to Tea Plantations: A Case Study in Southeast China. Forests 2023, 14, 209. [Google Scholar] [CrossRef]
- Guo, X.; Yang, Y.; Niu, Z.; Lu, D.P.; Zhu, C.; Feng, J.; Wu, J.; Chen, Y.; Tou, F.; Liu, M.; et al. Characteristics of Microbial Community Indicate Anthropogenic Impact on the Sediments along the Yangtze Estuary and Its Coastal Area, China. Sci. Total Environ. 2019, 648, 306–314. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Y0 2 | Y6 | Y12 | Y23 | Y35 |
|---|---|---|---|---|---|
| pH 3 | 5.0 ± 0.22 4,a | 3.84 ± 0.16 c | 3.63 ± 0.2 c | 4.46 ± 0.16 b | 4.25 ± 0.34 b |
| SOM (g kg−1) | 14.5 ± 5.81 c | 21.67 ± 4.85 bc | 26.04 ± 4.82 ab | 28.84 ± 8.14 ab | 34.82 ± 6.26 a |
| TN (g kg−1) | 0.69 ± 0.16 b | 1.38 ± 0.22 a | 1.48 ± 0.39 a | 1.52 ± 0.28 a | 1.8 ± 0.2 a |
| C/N | 11.92 ± 2.17 a | 9.1 ± 1.0 b | 10.43 ± 1.72 ab | 10.86 ± 1.4 ab | 11.25 ± 1.6 ab |
| TP (g kg−1) | 0.16 ± 0.03 c | 0.38 ± 0.02 bc | 0.53 ± 0.14 bc | 0.81 ± 0.32 b | 1.31 ± 0.61 a |
| TK (g kg−1) | 11.28 ± 4.98 b | 7.82 ± 1.89 b | 10.3 ± 3.34 b | 19.16 ± 7.91 a | 9.06 ± 1.05 b |
| AN (mg kg−1) | 53.82 ± 17.54 c | 202.04 ± 33.85 ab | 260.58 ± 128.66 a | 135.01 ± 28.16 bc | 181.27 ± 28.76 ab |
| AP (mg kg−1) | 1.03 ± 0.24 b | 10.94 ± 3.58 b | 70.26 ± 22.99 ab | 102.67 ± 68.66 a | 140.03 ± 95.06 a |
| AK (mg kg−1) | 26.19 ± 5.74 c | 106.81 ± 17.85 b | 134.52 ± 7.26 a | 135.36 ± 22.51 a | 140.4 ± 24.59 a |
| BD | 1.13 ± 0.07 a | 1.04 ± 0.1 a | 1.11 ± 0.12 a | 1.1 ± 0.03 a | 1.05 ± 0.08 a |
| Microbial Community | Diversity Indices | Y0 2 | Y6 | Y12 | Y23 | Y35 |
|---|---|---|---|---|---|---|
| Bacteria | ACE | 1168.22 ± 21.47 b, 3 | 1209.66 ± 43.74 b | 1156.59 ± 65.86 b | 1402.17 ± 86.74 a | 1392.16 ± 30.82 a |
| Chao1 | 1205.3 ± 33.18 b | 1239.94 ± 37.74 b | 1201.53 ± 50.16 b | 1419.15 ± 74.38 a | 1408.89 ± 35.78 a | |
| Simpson | 0.9909 ± 0.0008 ab | 0.9834 ± 0.0074 bc | 0.9776 ± 0.0099 c | 0.9924 ± 0.0042 ab | 0.9932 ± 0.0031 a | |
| Shannon | 8.07 ± 0.14 b | 7.86 ± 0.24 b | 7.63 ± 0.37 b | 8.58 ± 0.45 a | 8.57 ± 0.29 a | |
| Fungi | ACE | 509.44 ± 70.33 a | 454.6 ± 63.29 b | 458.99 ± 52.06 b | 420.81 ± 57.71 bc | 357.49 ± 24.78 c |
| Chao1 | 427.79 ± 69.7 ab | 468.44 ± 64.67 a | 456.59 ± 35.8 a | 430.18 ± 38.97 ab | 368.84 ± 30 b | |
| Simpson | 0.891 ± 0.0343 a | 0.9435 ± 0.0268 a | 0.9303 ± 0.0131 a | 0.9401 ± 0.0445 a | 0.8828 ± 0.0569 a | |
| Shannon | 4.67 ± 0.52 a | 5.62 ± 0.72 a | 5.05 ± 0.3 a | 5.5 ± 0.86 a | 4.65 ± 0.65 a |
| Microbial Community | Factor | pH 2 | SOM | TN | TP | TK | AN | AP | AK | BD | C/N |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacteria | ACE | 0.195 | 0.681 ** | 0.615 ** | 0.543 * | 0.189 | 0.059 | 0.412 | 0.443 | −0.293 | 0.132 |
| Chao1 | 0.179 | 0.703 ** | 0.614 ** | 0.536 * | 0.206 | 0.072 | 0.415 | 0.450 * | −0.270 | 0.190 | |
| Simpson | 0.625 ** | 0.179 | −0.044 | 0.166 | −0.029 | −0.515 * | 0.064 | −0.163 | −0.021 | 0.428 | |
| Shannon | 0.484 * | 0.436 | 0.277 | 0.307 | 0.038 | −0.276 | 0.159 | 0.157 | −0.051 | 0.327 | |
| Fungi | ACE | 0.059 | −0.429 | −0.549 * | −0.508 * | 0.096 | −0.246 | −0.360 | −0.582 ** | −0.065 | 0.197 |
| Chao1 | −0.332 | −0.174 | −0.142 | −0.302 | 0.043 | 0.112 | −0.217 | −0.125 | −0.409 | −0.135 | |
| Simpson | −0.260 | 0.258 | 0.243 | −0.187 | −0.140 | 0.220 | −0.215 | 0.263 | −0.008 | 0.058 | |
| Shannon | −0.236 | 0.320 | 0.282 | −0.100 | −0.167 | 0.149 | −0.148 | 0.226 | −0.242 | 0.086 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Wu, Y.; Xu, A.; Lin, C.; Wang, H.; Yu, J.; Ding, H.; Zhang, Y. Response of Soil Microbial Communities and Functions to Long-Term Tea (Camellia sinensis L.) Planting in a Subtropical Region. Forests 2023, 14, 1288. https://doi.org/10.3390/f14071288
Zheng X, Wu Y, Xu A, Lin C, Wang H, Yu J, Ding H, Zhang Y. Response of Soil Microbial Communities and Functions to Long-Term Tea (Camellia sinensis L.) Planting in a Subtropical Region. Forests. 2023; 14(7):1288. https://doi.org/10.3390/f14071288
Chicago/Turabian StyleZheng, Xiangzhou, Yiqun Wu, Aiai Xu, Cheng Lin, Huangping Wang, Juhua Yu, Hong Ding, and Yushu Zhang. 2023. "Response of Soil Microbial Communities and Functions to Long-Term Tea (Camellia sinensis L.) Planting in a Subtropical Region" Forests 14, no. 7: 1288. https://doi.org/10.3390/f14071288