1. Introduction
Urban trees are an essential component of urban green spaces, playing a crucial role in enhancing the well-being of city inhabitants. Urban trees offer myriad benefits, including reducing the urban heat island effect (UHI), mitigating the effects of climate change by removing atmospheric CO
2 [
1,
2], moderating microclimates [
3], and providing shade by reducing the temperatures on surfaces under tree canopies, particularly in the summer months in arid cities [
4]. Additionally, these urban green spaces covered by trees also offer a variety of social and cultural benefits, including recreational opportunities, aesthetic value, and potential inspiration for the arts and other creative endeavors [
5]. Furthermore, urban trees ameliorate the thermal environment of surroundings, and provide cooling effects through evapotranspiration and shading, thereby regulating local and regional climates [
6,
7,
8,
9].
Urban streets, particularly in semi-arid regions, can experience a significant increase in temperature, ranging from 3 to 6 °C compared to the surrounding rural environment [
10]. Semi-arid regions account for 42% of the total global land area and support approximately 38% of the global population, and are often located in developing countries [
11]. The provision of tree benefits largely depends on tree growth, which can vary with a range of microenvironmental and other site-specific factors [
12], for instance, anthropogenic disturbances such as mechanical injury [
13,
14], low soil quality [
15], sealed surfaces reducing water availability for tree roots [
16], and limited rooting space [
17], soil compaction [
18], and reduced nutrient resources and soil aeration [
19,
20].
These disturbances are often location-dependent, and the risks they pose to tree vitality can vary substantially over small areas—depending, for instance, on planting locations in parking lots, gardens, squares, or streets. Rötzer et al. [
21] have found that streets, paved squares, rooftops, and car parks limit the growth of trees, while larger gardens and public green spaces, such as parks and cemeteries, can provide ideal habitats for trees. Sanders et al. [
22] demonstrate that planting space has a significant impact on tree growth, with trees planted in reduced space exhibiting reduced maximum size.
In semi-arid regions specifically, irregular rainfall, poor tree management practices, and drought stress can also negatively impact urban tree growth [
23,
24,
25], and could influence the benefits trees are able to provide. Because the effective management of urban trees depends on a detailed understanding of the effects of growing environment, a substantial and growing literature seeks to evaluate the effects of climate change on urban tree growth rates in various climate zones [
23,
24,
26]. Several factors can reduce tree growth in arid and warm areas where water resources are limited [
27,
28]. In contrast, a few studies observed that some factors may increase the urban tree growth rate compared to rural trees, e.g., [
29,
30,
31,
32], including, for instance lower ozone concentration, larger annual atmospheric N deposition, and higher CO
2 concentration [
31,
33].
Considering the various factors influencing urban tree growth and their ecosystem services, recent research on tree growth and structure in urban green spaces has focused on monitoring and understanding these changes. By studying the relationships between structural variables such as leaf area index, crown dimension, tree height, and stem diameter, it is possible to model growth patterns and predict ecosystem services provisioning. This information can aid in the improvement of planning and management practices for urban landscapes [
34]. However, urban tree growth in arid cities is poorly understood, which impedes modelling and limits the available evidence base for planners and managers.
City planners, for instance, must take into account the ability of urban trees to acclimate to their surroundings and the structural variables that affect their future growth in order to optimize their benefits and ensure their long-term survival in an urban environment [
3]. As such, the structural development of urban trees, including size and shape, is closely linked to the benefits they provide [
35]. For instance, the area and density of shading from solar radiation is largely a function of the shape and volume of tree crowns [
36], while carbon sequestration and storage are driven by biomass and growth increment [
37,
38].
Moser et al. [
39] developed a regression equation to predict future structural dimensions through direct field measurements based on tree diameter and age. Issa et al. [
40] used crown dimensions to create an allometric equation to calculate total biomass, serving as a basis for remote sensing prediction and biomass assessment.
Understanding the relationship between structural variables of trees such as tree height, diameter at breast height, crown dimensions, and crown volume is essential to predict growth and ecosystem services [
41,
42]. Typically, diameter at breast height (dbh) is used to estimate tree growth based on the pipe model theory and functional carbon balance theory [
43,
44,
45]. These theories allow for the derivation of tree structure and biomass from basic tree measurements.
Pretzsch et al. [
46], and Watt et al. [
47] use dbh as an explanatory variable to predict crown dimensions. Although allometric equations for urban tree species have been developed for tropical and temperate regions [
48,
49], studies about the structural dimensions and ecosystem services of urban trees in arid cities are scarce. Despite limited research on the tree growth patterns of urban trees in arid cities [
50], there is a growing need to understand the factors that influence their growth and survival in these challenging environments. This research can provide a basic understanding of the structural dimensions and ecosystem services of urban trees in arid cities. We therefore analyzed the structural variables of urban trees in an arid city and estimated their carbon storage and shading potential (shaded area and shade density) as ecosystem services. We also aimed to examine the influence of site conditions, such as tree planting urban location and total unsealed area (tree pit surface area), on the tree structural variables, to understand the relationship between commonly planted urban tree structural variables and their effect on selected ecosystem services. The following hypotheses were tested:
Hypotheses 1 (H1). For each of the tree species, Delonix regia, Ficus nitida and Phoenix dactylifera, significant different relationships exist in terms of
- (a)
Tree height and crown dimensions with diameter at breast height (dbh, independent parameter).
- (b)
dbh, tree height, and crown dimensions with tree age (independent parameter)
- (c)
dbh, tree height, and crown dimensions with leaf area index (LAI, independent parameter)
- (d)
dbh, tree height, and crown dimensions with tree pit surface area (independent parameter).
Hypotheses 2 (H2). Tree planting urban location has a significant influence on tree structural variables (tree height, dbh, crown dimension), and ecosystem services.
Hypotheses 3 (H3). The ecosystem services of carbon storage and shading potential of the three tree species differ significantly from each other.
4. Discussion
A quantitative understanding of the structure and dimensions of urban trees is critical to better predict tree ecosystem services. However, the relationships between tree structure and ecosystem services in arid regions are poorly understood. Therefore, we applied several possible numerical approaches to calculate the structure and ecosystem services of trees. We analyzed the dependency of tree structure on dbh and age and the dependency of LAI and tree structure on three common urban trees in the arid city of Jericho. We also studied the effect of the different urban planting locations and tree pits on urban trees’ dimensions and on their ecosystem services.
The study outcomes provide a basic understanding for further research on the relationship between urban trees structure and ecosystem services in arid regions. It offers valuable insights into the growth patterns of arid urban trees, (e.g., dbh, crown dimension, and age) and their ability to acclimate (by showing growth efficiency that is not native to this region, for example, F. nitida and D. regia). Additionally, an allometric model was built to visualize the impacts of the tree structural variables on the ecosystem services, such as above-ground biomass carbon storage and the shade potential of urban trees based on the relationship between tree structure and ecosystem services. The study highlights the important role of urban trees in providing ecosystem services in arid regions and offers valuable insights for city planners and urban managers in their efforts to improve urban tree selection and create sustainable and resilient urban ecosystems in arid cities.
4.1. Relationship between Structural Tree Parameters (dbh, Age, Tree Pit Surface Area, and Tree Urban Location)
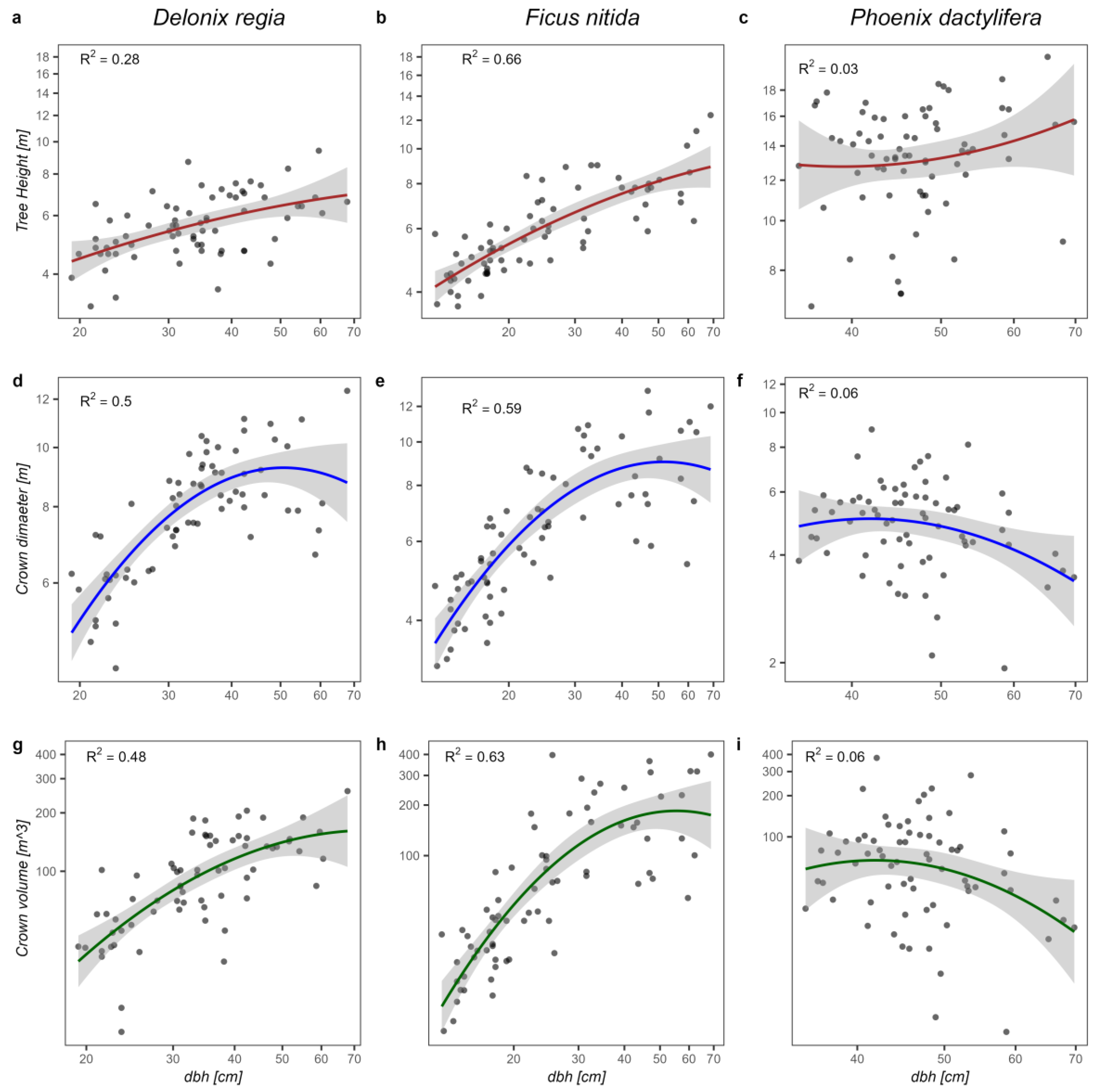
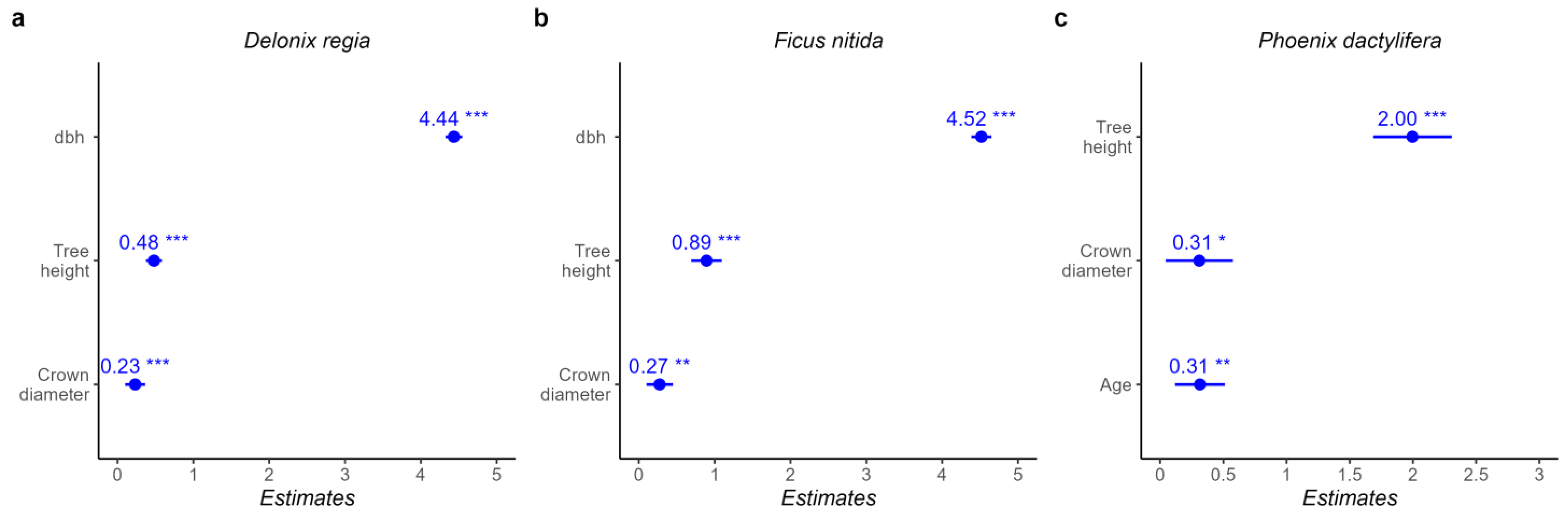
The results indicated a moderate to strong relationship between age and tree structure for
F. nitida (
R2 = 0.3–0.61), which is slightly weaker than the relationships obtained by Moser et al. [
39] for three different urban tree species in central Europe. Our results for
P. dactylifera and
D. regia show a weak and nonsignificant proportion of variance between age and tree structural variables in both species (R
2 ≤ 0.06).
The availability of resources limited annual precipitation, competition for above- and below-ground space, and poor soil quality, influence the relationship between age and dbh [
41,
81]. The stem diameter at breast height with tree height and crown dimension shows strong to moderate relationships for
F. nitida, but the relationship was slightly weaker in
D. regia, as a light-demanding and shade-intolerant tree, whereas
F. nitida is light-demanding but partially shade-tolerant [
61]. Light-demanding tree species have weaker stem diameter and crown volume relationships [
3]. The growth allocation of trees can greatly change in response to light availability [
82], which also supports our results. The results indicate that street trees, which are often planted in close proximity to one another, experience increased competition for sunlight, particularly when their crowns come into contact with each other. Light availability is a critical factor that can influence the growth and development of trees. Light-demanding tree species, such as those that typically grow in open habitats, require high levels of sunlight to thrive. Specifically, these trees may allocate more resources to the production of leaves and branches, which can increase their ability to capture sunlight and produce energy. This may result in weaker stem diameter and crown volume relationships [
3].
The tree structural relationships of
D. regia illustrate a moderate trend that is slightly weaker than those of the studies conducted by Arzai et al. [
50], who investigated the connections among canopy width, tree height, and dbh of various urban tree species, finding a strong correlation between tree height and crown diameter with dbh, as an adaptive tree species. This difference is possibly based on the natural climate of the study area, which is tropical [
83].
Many other factors, such as annual pruning to shape the tree, especially at an early stage [
84], and the removal of damaged, dead, dried, and crossing branches [
85], can also affect crown dimension–dbh relationships. Pruning mature trees may be for reasons of shape, tree health, aesthetics, safety, or clearance from infrastructure [
86]. The correlation between stem diameter and the crown dimension of
P. dactylifera was nonsignificant. As a monocotyledonous plant,
P. dactylifera lacks the ability to form a vascular cambium, a meristem tissue that allows for secondary growth in dicotyledonous plants. The vascular cambium is responsible for the formation of new layers of xylem and phloem, which contribute to the increase in diameter of the plant’s stem or trunk over time. Without the formation of a vascular cambium, the date palm does not undergo regular secondary growth and does not exhibit the characteristic increase in diameter [
64]. This is in line with the results of Issa et al., [
40], whose regression coefficient shows weak but significant relationships between dbh and crown area for
P. dactylifera.
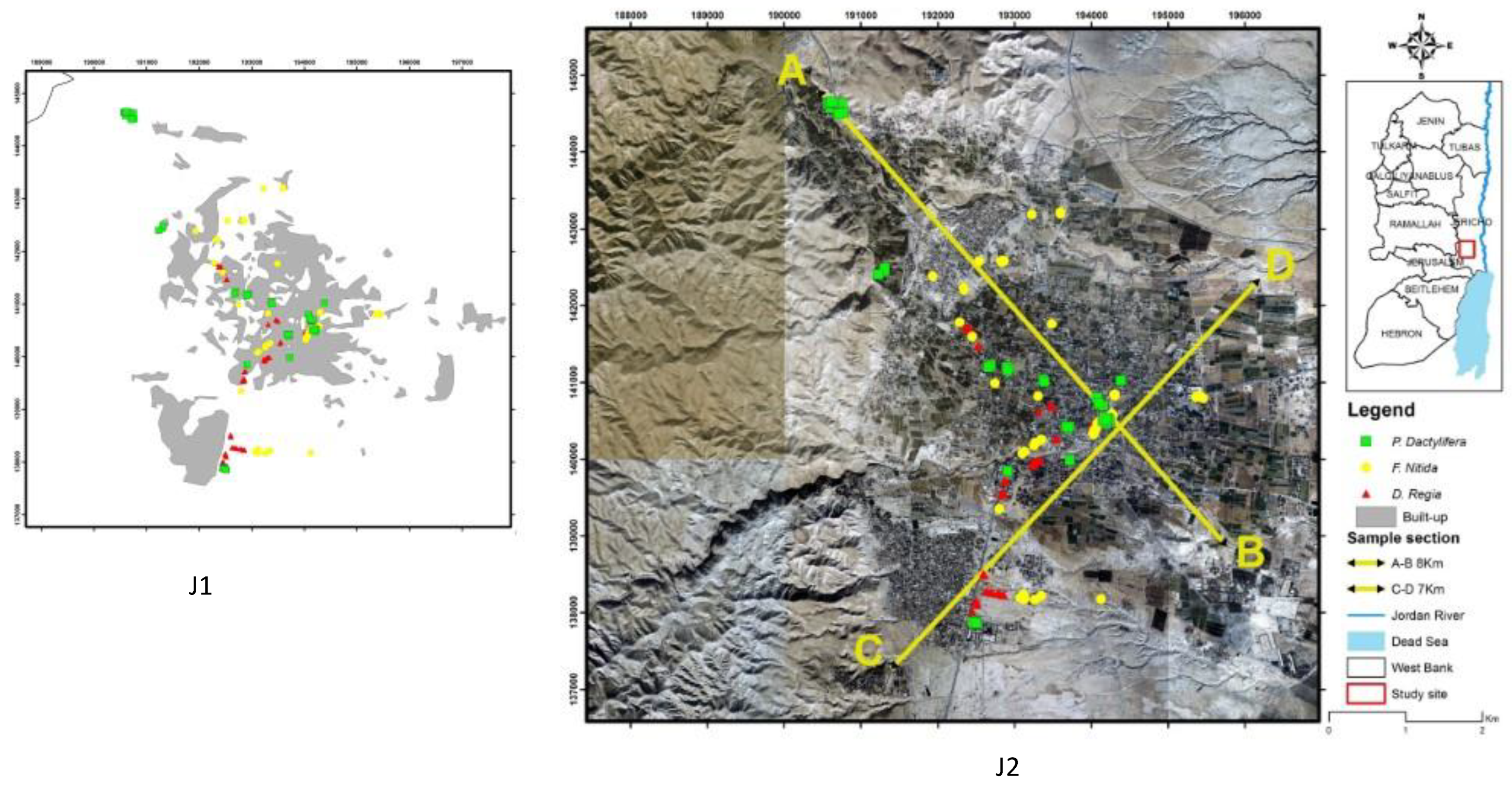
Generally, tree samples were selected from different urban locations, that typically suffer from a scarcity of water due to the lack of a regular irrigation system. Our results show a significant difference in dbh in the tree planting site for F. nitida, and a significant difference in dbh and age in the tree planting site for D. regia.
In Jericho City, many irrigation patterns exist (water transportation tanks, manual plastic tubes, normal irrigation systems, and normal water buckets). Additionally, some street trees are situated close to agricultural farms that provide them with resources (water and nutrition).
However, the research of Coombes et al. [
87] found that the site factors had very little effect on the allometric relationship between dbh and crown diameter. However, the results presented showed that the difference in irrigation patterns and the distribution of nutrient resources for trees in Jericho may lead to different growth patterns in urban areas; therefore, this may be the reason for the different ratio of tree structural relationships. In addition, the results showed differences in
F. nitida, in canopy diameter, and volume between parking lot and street trees due to tree size variations. For
D. regia trees in public places, the trees vary in size as well. The trees in the public place (e.g., garden) are older than the trees in the street, but there were no significant differences in
P. dactylifera at all, and the reasons behind the fact that the overall mean of
P. dactylifera tree ages in different urban locations of the city are not significantly different. Furthermore, the findings revealed that the relation between the tree pit surface area and tree structure for
F. nitida and
P. dactylifera are weak but statistically significant, but is not significant in
D. regia. The possible reason behind that uneven distribution of tree samples selected, e.g., 62 of
D. regia, is that most of the street trees had a very small tree pit surface area. Even if they were irrigated by the above irrigation patterns, the amount of water to reach the plant would be very small, especially in summertime with high evaporation rates.
4.2. Leaf Area Index of the Three Urban Tree Species
The results show a nonsignificant and weak proportion of variance between LAI and structural parameters. The R
2 values were close to zero for all variables. Özbayram et al. [
88], in their research, studied the correlation between LAI values and tree variables in Turkey, and a negative correlation in black pine stands was found (i.e., stand age, mean diameter) and a positive correlation in red pine (i.e., stand age, mean diameter, top height, green tree height, and basal area). Özbayram et al. concluded that the leaf area index (LAI) varies according to species. The LAI results were 5.4 for
F. nitida, 5.8 for
D. regia, and 2.9 for
P. dactylifera. These results can be placed in comparison with those of Liu et al., [
89], who found a mean LAI of value 4.73 ± 0.40 for
D. regia and 5.00 ± 0.47 for
F. nitida, whereas Lin et al. [
90] found an LAI of 6.11 for
Ficus macrocarpa and 5.05 for
Ficus elastica, and Awal et al. found an LAI of 1.7 for
P. dactylifera [
91]. A higher leaf area index means higher photosynthesis and efficient use of light, which indicates higher carbon capturing ability and stocks [
92].
4.3. Ecosystem Services of Trees in Arid Cities
Urban trees provide ecosystem services [
93,
94], which can significantly improve the climate in cities [
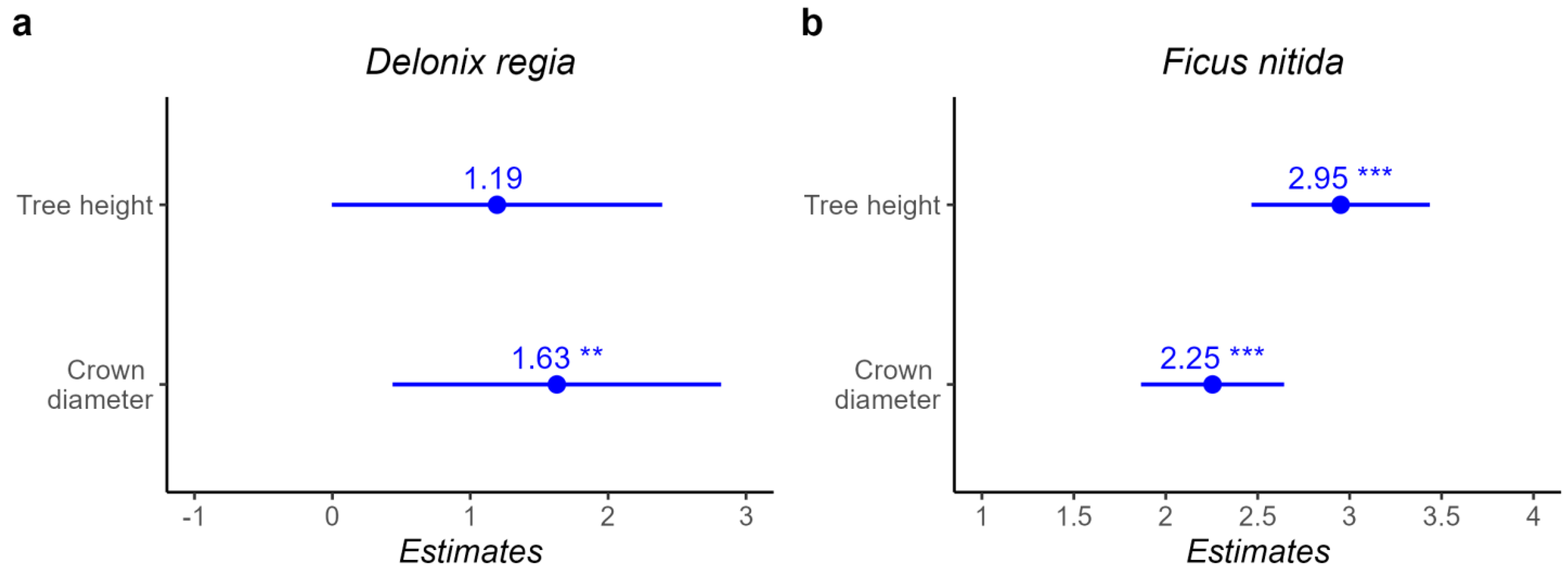
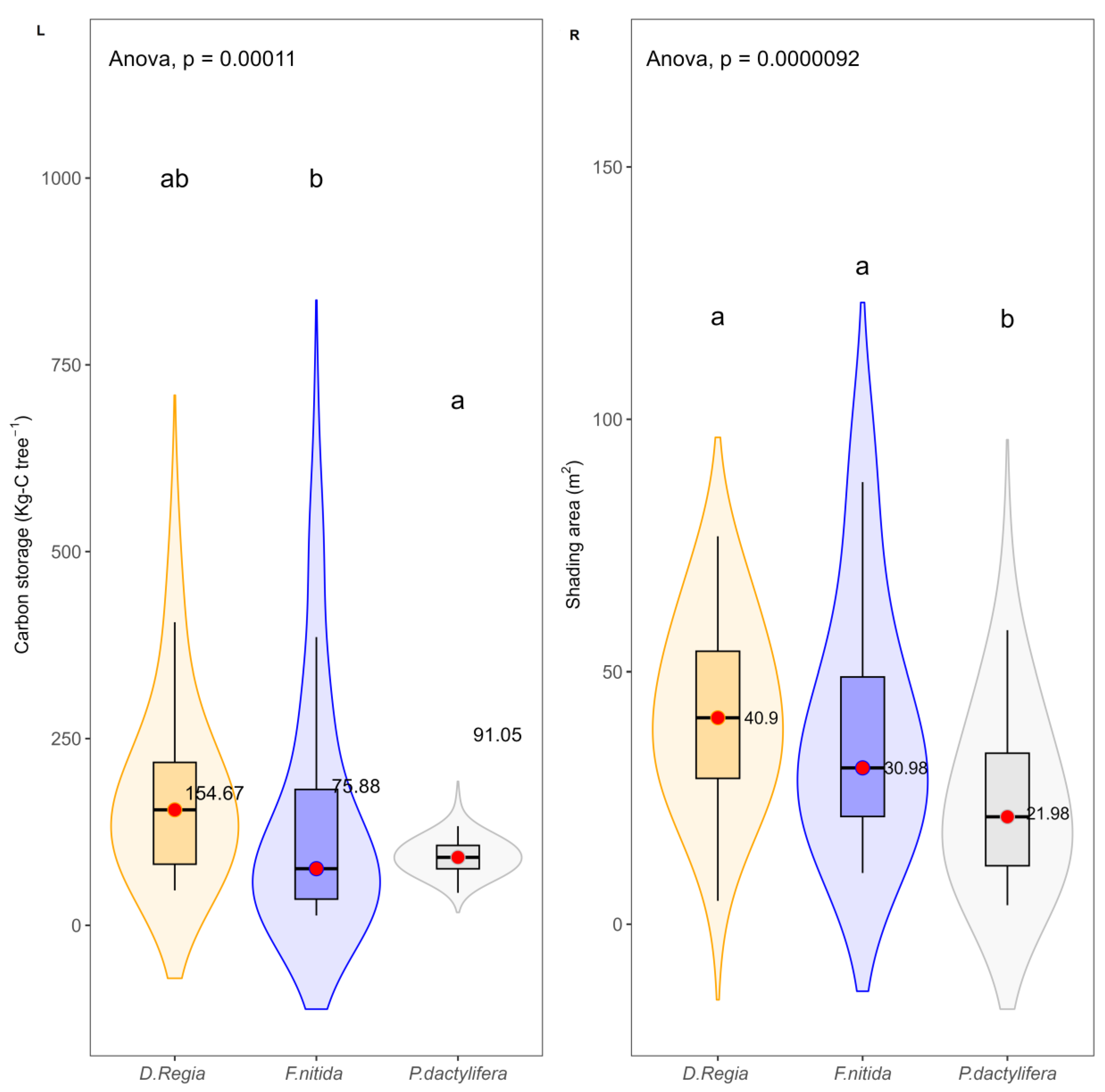
95]. The study estimated above-ground biomass carbon storage and shading potential. Results showed that tree height, dbh, and crown diameter have a strong relationship with above-ground biomass carbon storage in
D. regia and
F. nitida. Similarly, tree height, crown diameter, and age have a significant relationship with above-ground biomass carbon storage in
P. dactylifera, consistent with prior research, (e.g., Yoon et al. [
37]). Issa et al. [
96] found that the amount of CO
2 absorbed is proportional to the tree component, above-ground biomass can be highly estimated by the green plant component (e.g., canopy area) and tree stems as variables measured in the field. Further, Betemariyam et al. [
97], found that
P. dactylifera trees older than 20 years had a mean above-ground biomass carbon stock of 159.50 kg/plant, in date palm on a farm in north-eastern Ethiopia. Issa et al. [
40] found that trunk height and crown diameter are strongly correlated with the age of date palm trees and reported an average carbon storage of 225 kg C of the palm trees in Abu Dhabi, United Arab Emirates, for trees older than 20 years. The results show the average carbon storage of
P. dactylifera is higher than the averages of
D. regia and
F. nitida; these findings support that
P. dactylifera trees in this study contribute to emission reduction and carbon sink enhancement.
Higher above-ground biomass carbon storage averages for D. regia were found in public places and parking lots compared with street trees, whereas P. dactylifera trees provided similar rates at all sites. The second ecosystem service is shade potential. The results showed a statistically significant and strong relationship between tree height, diameter at breast height and crown diameter with the shaded area of D. regia and F. nitida, and a nonsignificant relationship with the shaded area of the P. dactylifera tree. This could be due to its monocotyledon nature. The results of the P. dactylifera shade area show a smaller value for older trees, where the most likely reason could be leaf senescence due to age. With age, trees may lose some leaf area due to leaf senescence. Another reason could be leaf pruning each year.
Different shaded areas and shading densities exist among urban tree species.
F. nitida and
D. regia have the highest shading potential compared to
P. dactylifera. Shade density is particularly important for lowering surface temperatures and improving thermal comfort [
90,
98,
99].
F. nitida and
D. regia have higher shading potential compared to
P. dactylifera, as produced by their crown canopies. A possible reason is that the sampled trees are mixed between taller trees with narrow canopies and shorter trees with wider canopies. This result is in line with Armson et al., [
6], with the outcome about tree morphology and shade for five different street tree species in Manchester (UK) exhibiting a significant difference between the species’ canopy sizes but nonsignificant differences between tree canopies’ shaded areas. Rahman et al. [
100] demonstrated that urban trees can mitigate temperatures underneath canopy surfaces during the day through shading.
The potential cooling effect of tropical trees is higher than that of other species, e.g.,
Ficus retusa trees can reduce the temperature values during the summer by 4 °C, while the cooling effect for date palm trees (
P. dactylifera) is only 1.5 °C, which is characterized by a small canopy [
101]. Reflecting the weakness of
P. dactylifera as a tree for shading benefits and the higher shading potentials of
F. nitida and
D. regia, which are characterized as tropical trees in arid cities, shading measures have special importance, where the sun has intense solar radiation, leading to higher air temperatures that can negatively affect most human daily activities [
102]. Based on previous studies, the importance of cooling by shading in an arid city is particularly important where solar radiation is intense, leading to higher temperatures that can negatively impact human activities. Overall, this study provides valuable insights into the ecosystem services provided by urban trees, specifically carbon storage and shading potential, and their correlation with structural variables.
5. Conclusions
In conclusion, this study analyzed the growth, ecosystem services, and tree structural characteristics of three common urban trees (i.e., D. regia, F. nitida, and P. dactylifera) in the arid city of Jericho using a numerical approach of a City Tree model. The results showed that tree structural variables (i.e., tree height, crown volume, and crown diameter) have a strong to moderately significant relationship with dbh for D. regia and F. nitida. The results also show no relationships between leaf area index and tree structure for all tree species’ structural variables, while showing a statistically moderate relationship for tree structure with age for F. nitida, and no relationship for all other tree species. The tree pit surface area also showed weak significant relationship with tree structure for F. nitida and P. dactylifera, but not for D. regia. Different urban plant growth location also induced various influences among the three species; the results show a significant influence on tree structure for D. regia and F. nitida, while the influence was not significant on P. dactylifera.
D. regia has higher shading potential and above ground biomass carbon storage, compared to F. nitida and P. dactylifera, respectively, as common urban trees in the city. The results may vary based on species and site conditions. Our results are similar to research from other climates; for example, Moser et al. (2015) carried out similar research in Germany (temperate region) and found strong to moderate relationships between crown dimensions and stem diameter, which is identical to our results except for P. dactylifera. Although results can vary based on species and site conditions, overall patterns are comparable, which indicates that similar results are also applicable to other climate regions. However, species functionality should be considered.
Based on these findings for the selected ecosystem services (above-ground biomass carbon storage and shade potential), it is recommended that D. regia, F. nitida and P. dactylifera be considered for future urban greening in arid cities, with D. regia outperforming the others. However, further research in other non-arid regions and climate-sensitive growth models are needed to better understand the growth and adaptation capacity of these trees in changing climates. We recommend conducting further research on the relationship between tree species’ dimensions and the ecosystem services they provide, with a specific focus on urban areas in Mediterranean and/or arid climates.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}