
Diversity and Assembly of Bacteria Community in Lime Soil under Different Karst Land-Use Types

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
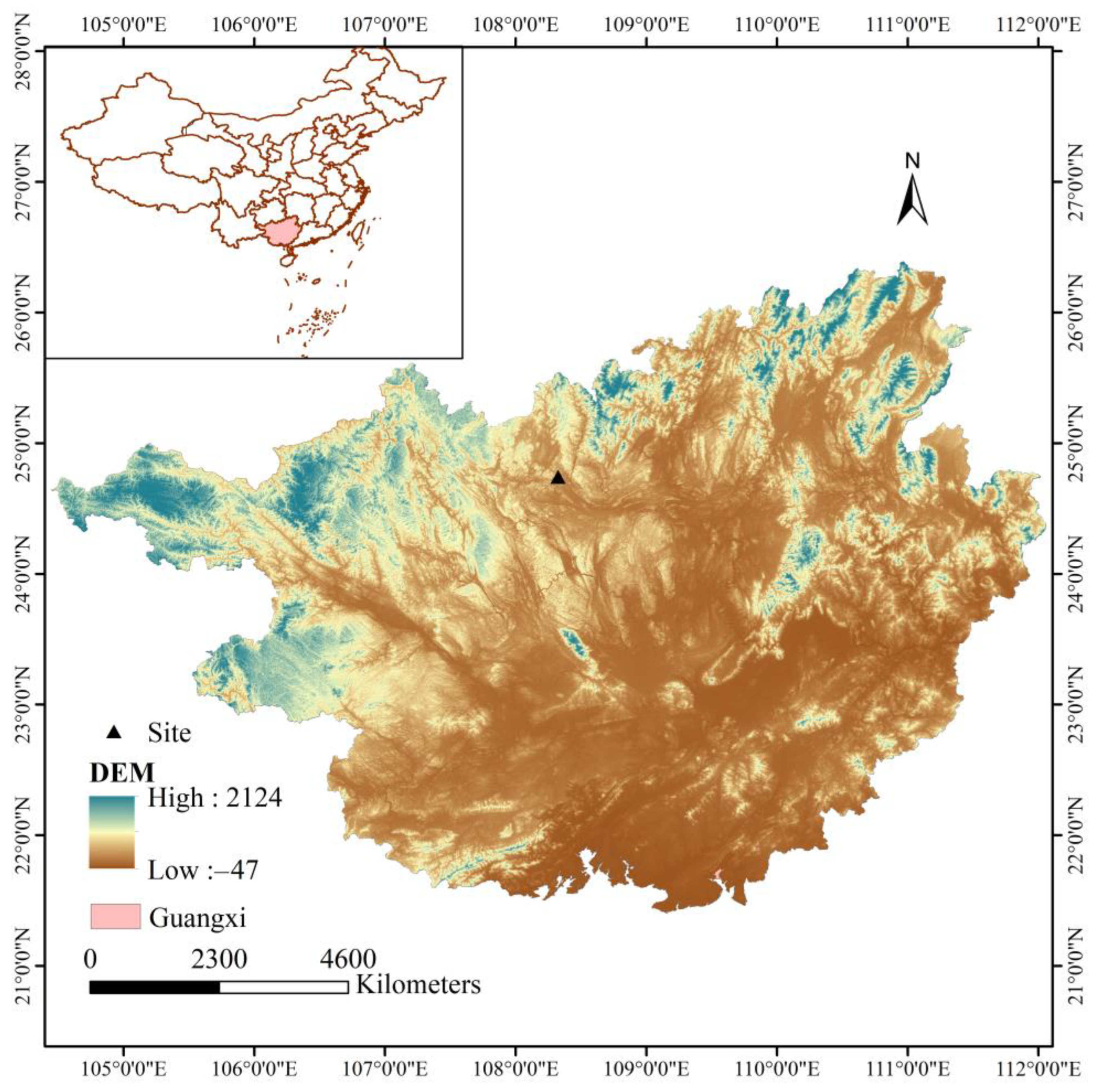
2.1. Research Site
2.2. Determination of Soil Chemical Properties
2.3. High-Throughput DNA Sequencing and Bioinformatics Analysis
2.4. Statistical Analysis
3. Results
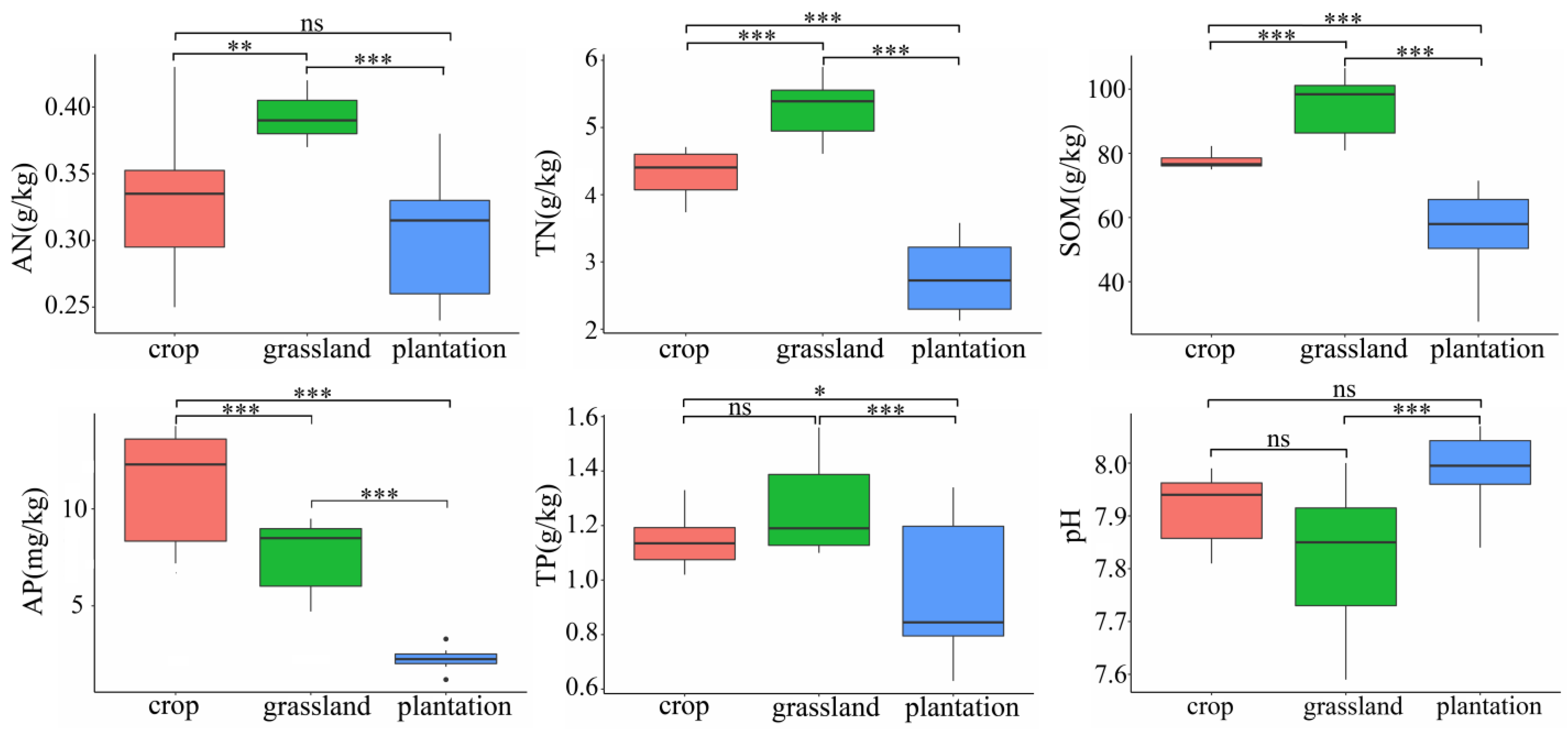
3.1. Soil Chemical Properties in Different Karst Land-Use Types
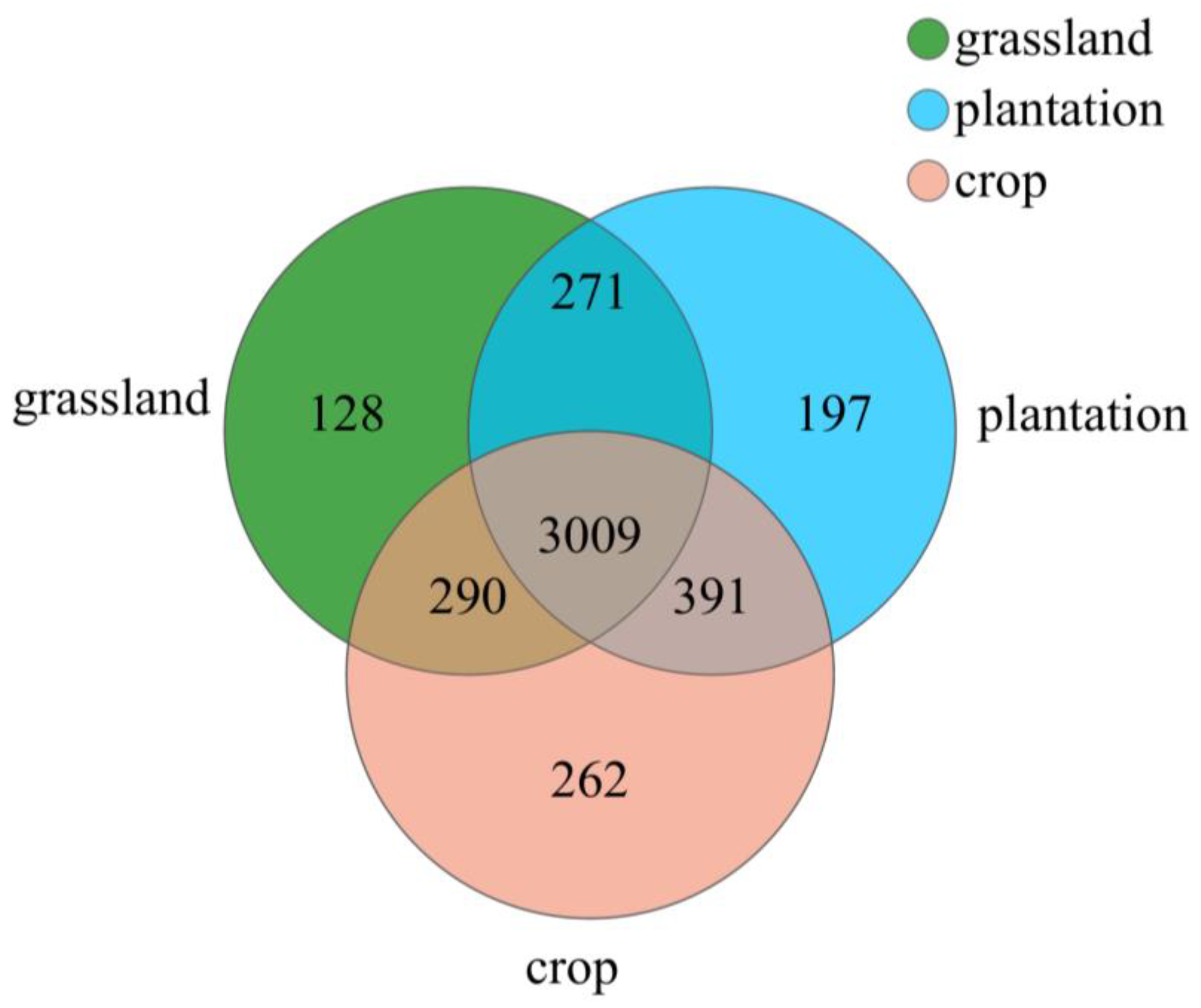
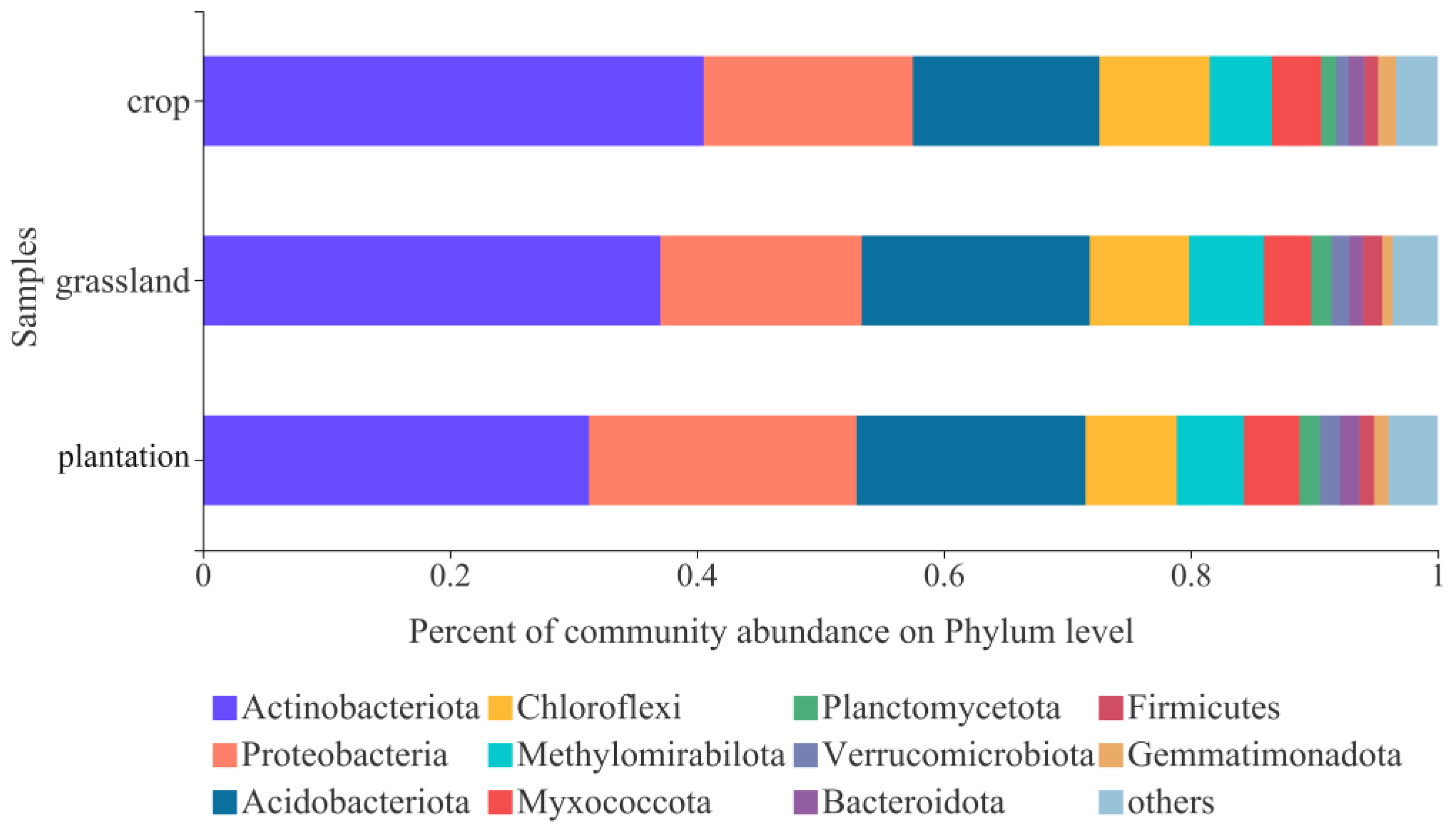
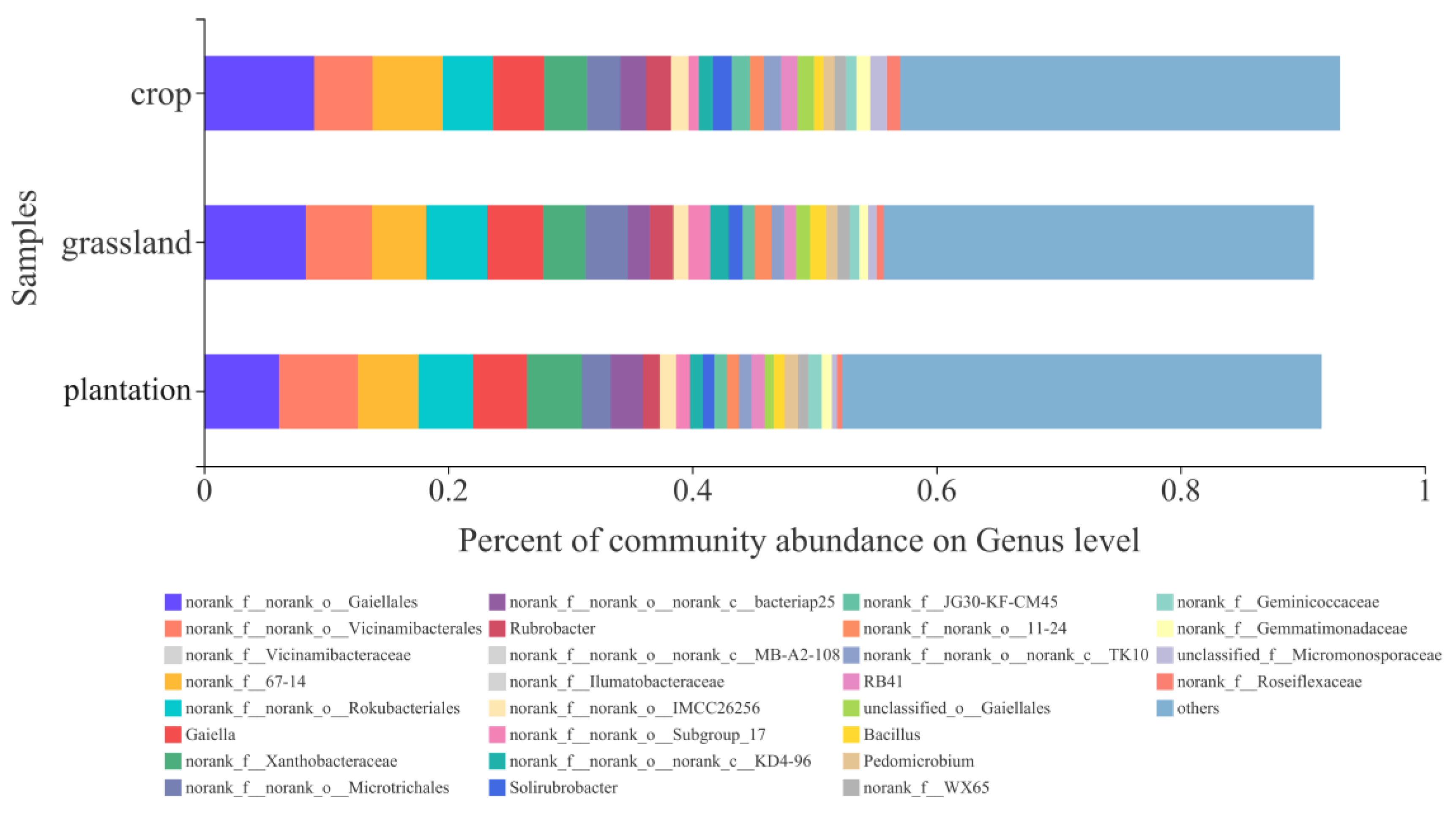
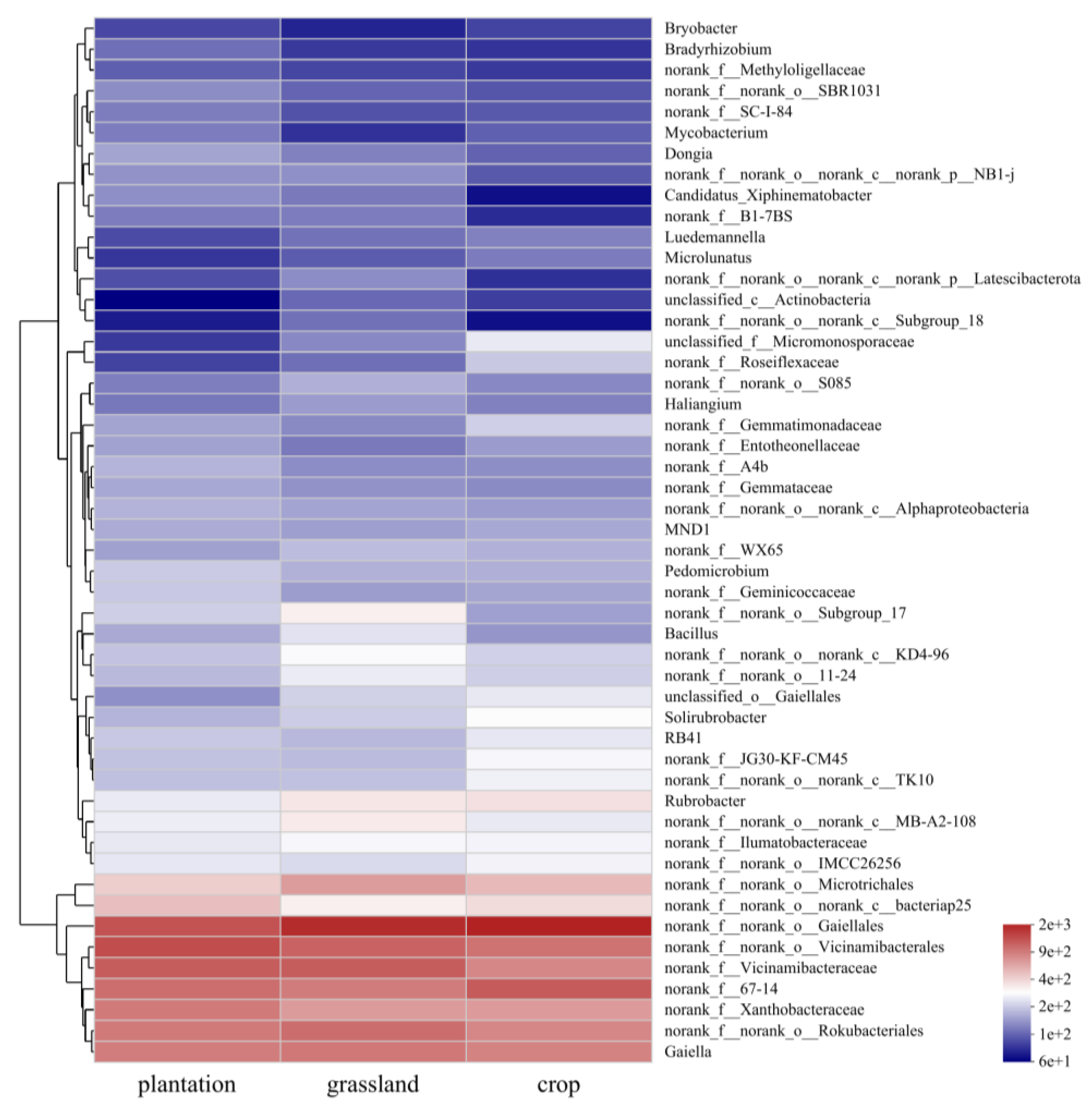
3.2. Soil Bacterial Community Composition in Different Karst Land-Use Types
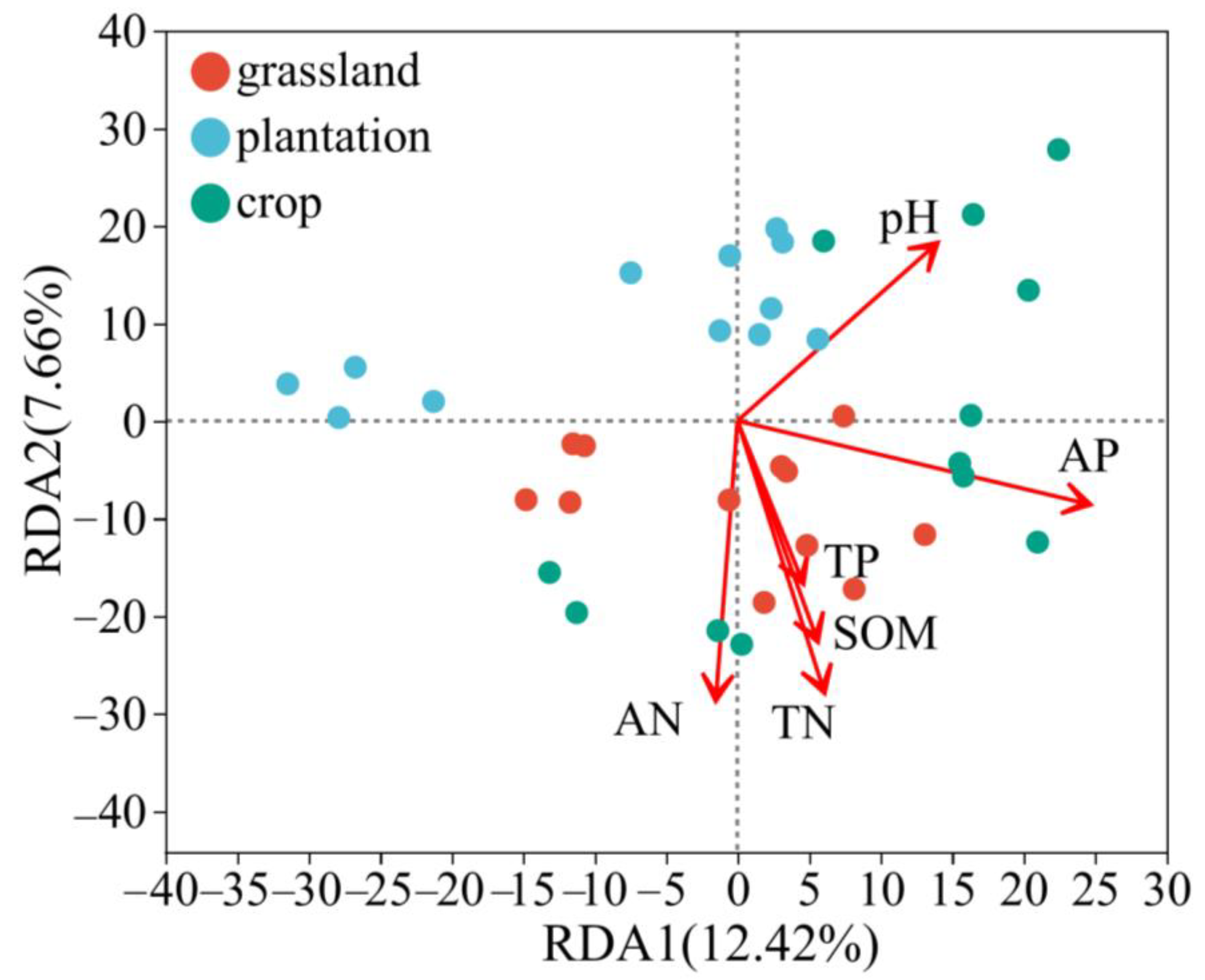
3.3. Association between Soil Chemical Properties and Soil Bacteria in Different Karst Land-Use Types
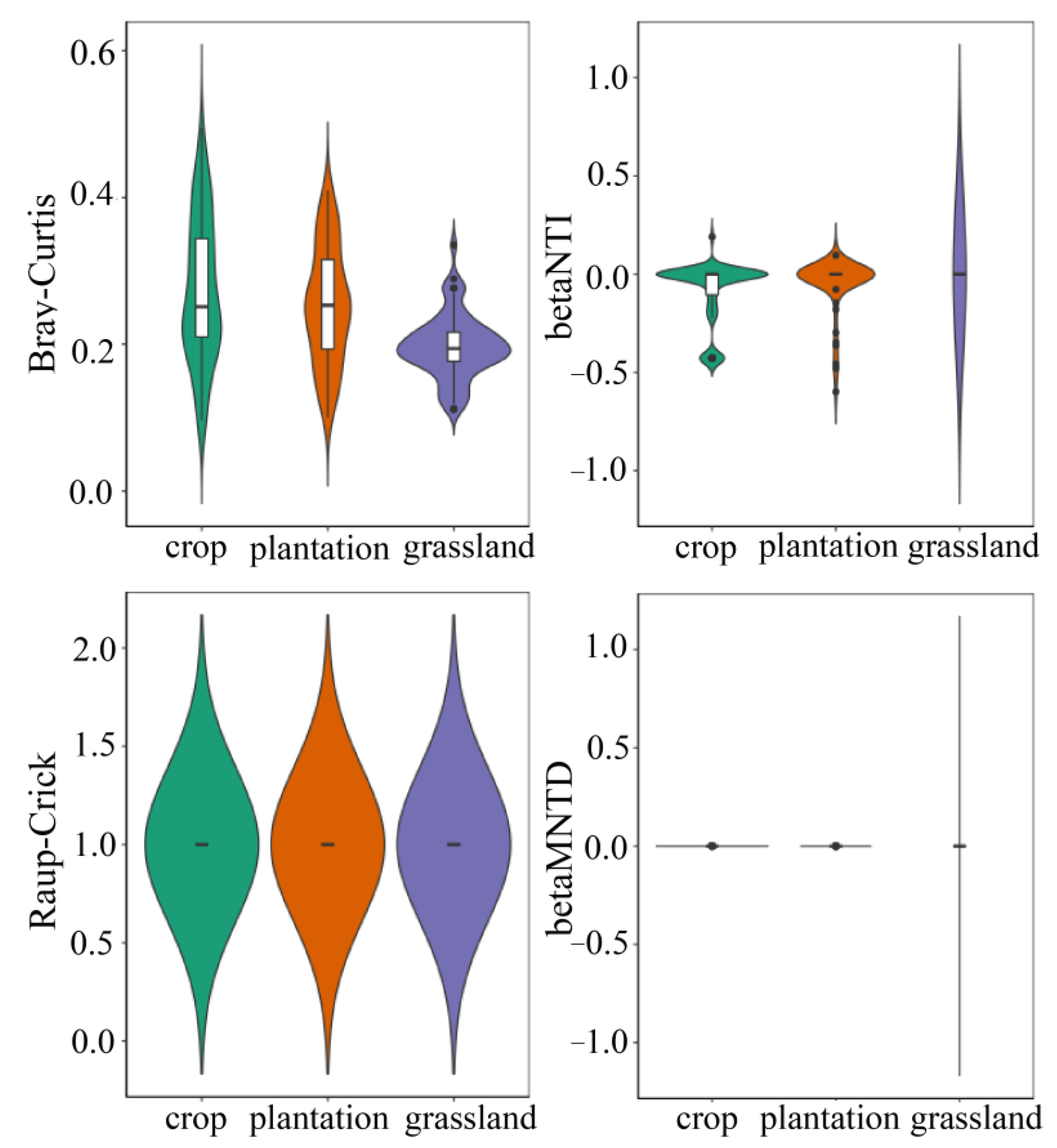
3.4. Assembly Mechanism of Soil Bacterial Community in Different Karst Land-Use Types
4. Discussion
4.1. Changes in Soil Chemical Properties in Different Karst Land-Use Types
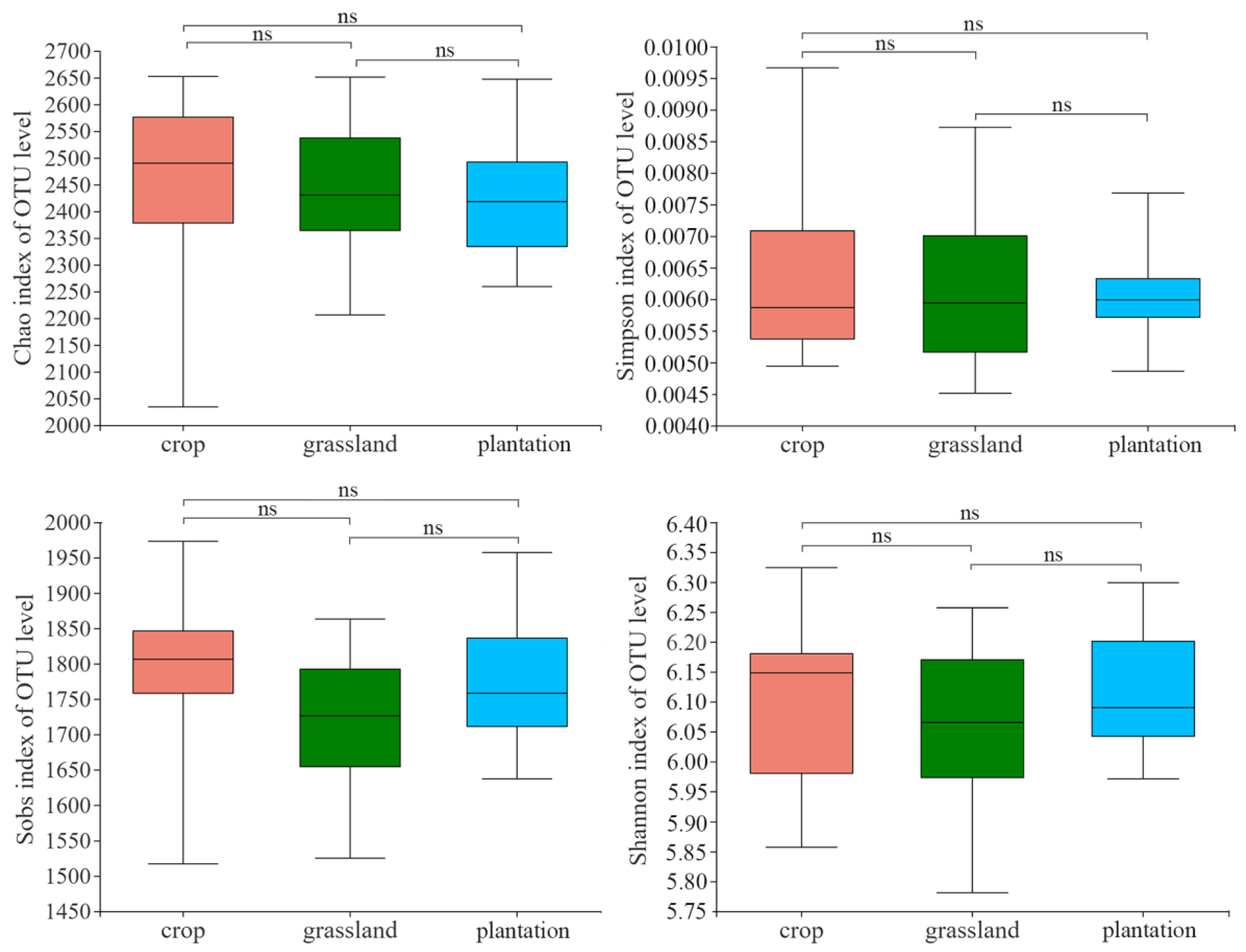
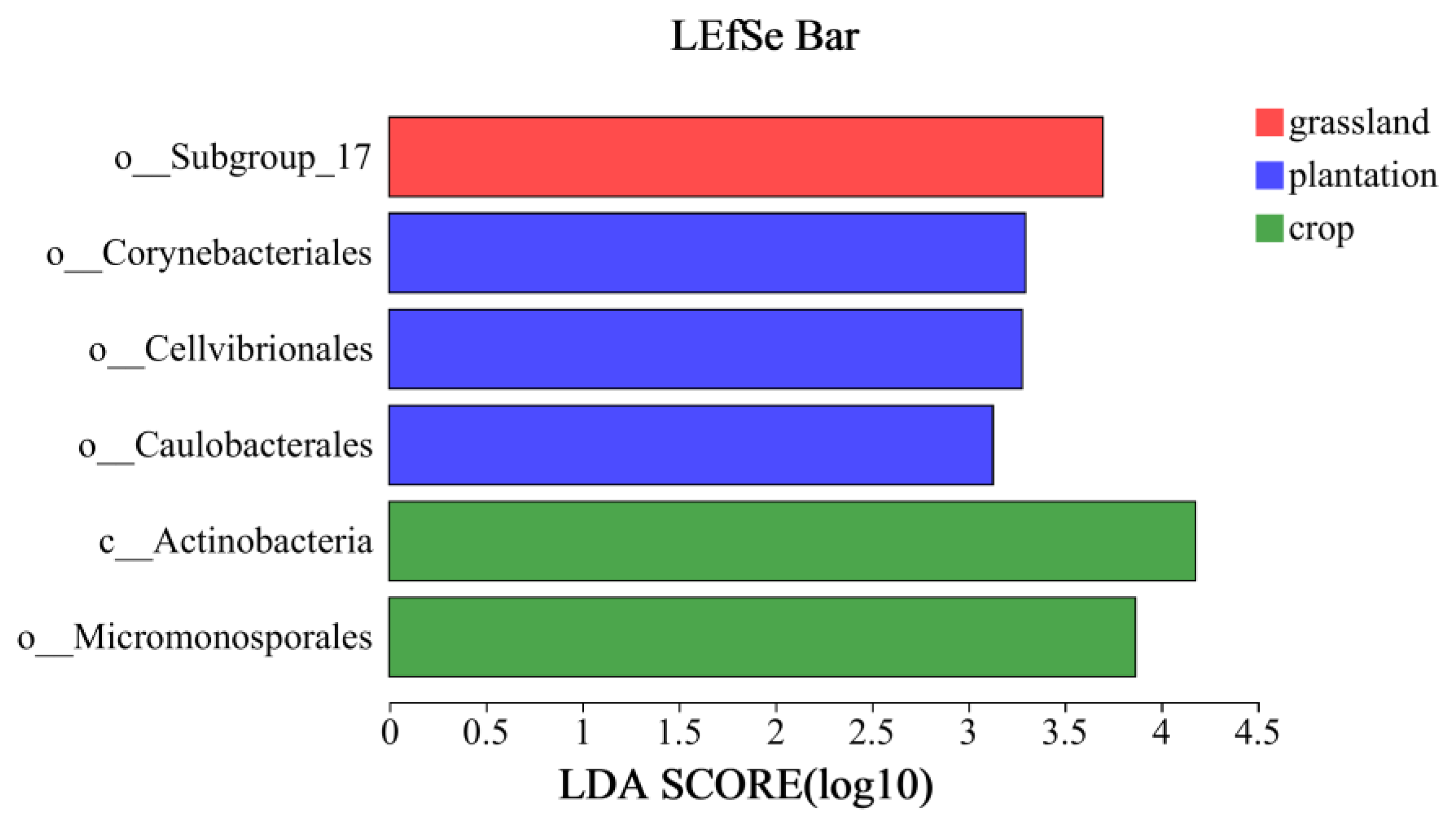
4.2. Changes in Soil Bacterial Community Diversity and Composition in Different Karst Land-Use Types
4.3. Effects of Soil Properties on Soil Microorganisms in Karst Land-Use Types
4.4. Community Assembly of Soil Bacteria in Three Karst Land-Use Types
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wardle, D.A. The influence of biotic interactions on soil biodiversity. Ecol. Lett. 2006, 9, 870–886. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Hu, J.; Jousset, A. More Than the Sum of Its Parts: Microbiome Biodiversity as a Driver of Plant Growth and Soil Health. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 145–168. [Google Scholar] [CrossRef]
- Herzog, S.; Wemheuer, F.; Wemheuer, B.; Daniel, R. Effects of Fertilization and Sampling Time on Composition and Diversity of Entire and Active Bacterial Communities in German Grassland Soils. PLoS ONE 2015, 10, e0145575. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.Z.; Deng, Y.; Shen, L.N.; Wen, C.Q.; Yan, Q.Y.; Ning, D.L.; Qin, Y.J.; Xue, K.; Wu, L.Y.; He, Z.L.; et al. Temperature mediates continental-scale diversity of microbes in forest soils. Nat. Commun. 2016, 7, 12083. [Google Scholar] [CrossRef] [Green Version]
- Xiao, D.; He, X.Y.; Zhang, W.; Hu, P.L.; Sun, M.M.; Wang, K.L. Comparison of bacterial and fungal diversity and network connectivity in karst and non-karst forests in southwest China. Sci. Total Environ. 2022, 822, 153179. [Google Scholar] [CrossRef]
- Hogberg, M.N.; Hogberg, P.; Myrold, D.D. Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three? Oecologia 2007, 150, 590–601. [Google Scholar] [CrossRef]
- Vellend, M. Conceptual synthesis in community ecology. Q. Rev. Biol. 2010, 85, 183–206. [Google Scholar] [CrossRef] [Green Version]
- Stegen, J.C.; Lin, X.J.; Konopka, A.E.; Fredrickson, J.K. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012, 6, 1653–1664. [Google Scholar] [CrossRef] [Green Version]
- Vilmi, A.; Zhao, W.Q.; Picazo, F.; Li, M.J.; Heino, J.; Soininen, J.; Wang, J.J. Ecological processes underlying community assembly of aquatic bacteria and macroinvertebrates under contrasting climates on the Tibetan Plateau. Sci. Total Environ. 2020, 702, 134974. [Google Scholar] [CrossRef]
- Tang, J.; Tang, X.X.; Qin, Y.M.; He, Q.S.; Yi, Y.; Ji, Z.L. Karst rocky desertification progress: Soil calcium as a possible driving force. Sci. Total Environ. 2019, 649, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.H.; Qi, X.K.; Wang, K.L.; Zhang, M.Y.; Yue, Y.M. The application of geospatial techniques in monitoring karst vegetation recovery in southwest China: A review. Prog. Phys. Geogr. Earth Environ. 2017, 41, 450–477. [Google Scholar] [CrossRef]
- Wang, K.L.; Zhang, C.H.; Chen, H.S.; Yue, Y.M.; Zhang, W.; Zhang, M.Y.; Qi, X.K.; Fu, Z.Y. Karst landscapes of China: Patterns, ecosystem processes and services. Landsc. Ecol. 2019, 34, 2743–2763. [Google Scholar] [CrossRef] [Green Version]
- Hu, P.L.; Xiao, J.; Zhang, W.; Xiao, L.M.; Yang, R.; Xiao, D.; Zhao, J.; Wang, K.L. Response of soil microbial communities to natural and managed vegetation restoration in a subtropical karst region. Catena 2020, 195, 104849. [Google Scholar] [CrossRef]
- Li, X.; Su, Z.; Liu, S.; Ou, Z.; Xiang, W.; Ou, Z.; Lu, S. The Spatial pattern of Natural Vegetation in the Karst Regions of Guangxi and the Ecological Signality for Ecosystem Rehabilitation and Reconstruction. J. Mt. Sci. 2003, 21, 129–139. [Google Scholar]
- Zhang, M.W.; Wang, Y.; Li, B.; Feng, Z.Y.; Zhao, Y.L.; Xu, Z.G. Synergistic succession of the small mammal community and herbaceous vegetation after reconverting farmland to seasonally flooded wetlands in the Dongting Lake Region, China. Mammal Study 2018, 43, 229–243. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.F.; Du, H.; Zeng, F.P.; Song, T.Q.; Peng, W.X. Diminished rhizosphere and bulk soil microbial abundance and diversity across succession stages in Karst area, southwest China. Appl. Soil Ecol. 2021, 158, 103799. [Google Scholar] [CrossRef]
- Chen, H.J.; Peng, W.X.; Du, H.; Song, T.Q.; Zeng, F.P.; Wang, F. Effect of Different Grain for Green Approaches on Soil Bacterial Community in a Karst Region. Front. Microbiol. 2020, 11, 577242. [Google Scholar] [CrossRef]
- Mi, Y.D.; Fang, H.D.; Tao, P.; Zhou, M.; Li, X.R.; Wang, F.F.; Chen, H.Y.; Su, H.L.; Zhu, Y.R.; Wei, Y.; et al. Comparation study of soil genetic diversity of bacteria and fungi in different vegetation successions in a karst of Guizhou province, China. Acta Carsol. 2021, 50, 329–342. [Google Scholar] [CrossRef]
- Chen, H.S.; Liu, J.W.; Wang, K.L.; Zhang, W. Spatial distribution of rock fragments on steep hillslopes in karst region of northwest Guangxi, China. Catena 2011, 84, 21–28. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000; pp. 263–270. [Google Scholar]
- Xu, M.P.; Gao, D.X.; Fu, S.Y.; Lu, X.Q.; Wu, S.J.; Han, X.H.; Yang, G.H.; Feng, Y.Z. Long-term effects of vegetation and soil on the microbial communities following afforestation of farmland with Robinia pseudoacacia plantations. Geoderma 2020, 367, 114263. [Google Scholar] [CrossRef]
- Ren, C.J.; Liu, W.C.; Zhao, F.Z.; Zhong, Z.K.; Deng, J.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Soil bacterial and fungal diversity and compositions respond differently to forest development. Catena 2019, 181, 104071. [Google Scholar] [CrossRef]
- Lan, J.C.; Wang, S.S.; Wang, J.X.; Qi, X.; Long, Q.X.; Huang, M.Z. The Shift of Soil Bacterial Community After Afforestation Influence Soil Organic Carbon and Aggregate Stability in Karst Region. Front. Microbiol. 2022, 13, 901126. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Yang, X.M.; Song, Y.H.; Liu, H.F.; Wang, G.L.; Xue, S.; Liu, G.B.; Ritsema, C.J.; Geissen, V. Revealing the nutrient limitation and cycling for microbes under forest management practices in the Loess Plateau—Ecological stoichiometry. Geoderma 2020, 361, 114108. [Google Scholar] [CrossRef]
- Lauber, C.L.; Ramirez, K.S.; Aanderud, Z.; Lennon, J.; Fierer, N. Temporal variability in soil microbial communities across land-use types. ISME J. 2013, 7, 1641–1650. [Google Scholar] [CrossRef]
- Liu, J.L.; Yang, Z.L.; Dang, P.; Zhu, H.L.; Gao, Y.; Ha, V.N.; Zhao, Z. Response of soil microbial community dynamics to Robinia pseudoacacia L. afforestation in the loess plateau: A chronosequence approach. Plant Soil 2018, 423, 327–338. [Google Scholar] [CrossRef]
- Buckley, D.H.; Schmidt, T.M. The structure of microbial communities in soil and the lasting impact of cultivation. Microb. Ecol. 2001, 42, 11–21. [Google Scholar] [CrossRef]
- Jangid, K.; Whitman, W.B.; Condron, L.M.; Turner, B.L.; Williams, M.A. Soil bacterial community succession during long-term ecosystem development. Mol. Ecol. 2013, 22, 3415–3424. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.C.; An, S.S.; Liu, Y. Soil bacterial community response to vegetation succession after fencing in the grassland of China. Sci. Total Environ. 2017, 609, 2–10. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, S.K.; Gao, Q.Z.; Liu, S.L.; Zhou, H.K.; Ganjurjav, H.; Wang, X.X. Climate change and human activities altered the diversity and composition of soil microbial community in alpine grasslands of the Qinghai-Tibetan Plateau. Sci. Total Environ. 2016, 562, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, G.B.; Xue, S.; Wang, G.L. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Foulon, J.; Appelini, C.; Durand, A.; Valot, B.; Girardclos, O.; Blaudez, D.; Chalot, M. Environmental metabarcoding reveals contrasting microbial communities at two poplar phytomanagement sites. Sci. Total Environ. 2016, 571, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Fang, Y.; Yang, Q.; Chen, X.; Yuan, F.; Xu, H.; He, L.; Yan, J.; Chen, T.; Yu, C.; et al. Community characteristics of a mid-subtropical evergreen broad-leaved forest plot in the Wuyi Mountains, Fujian Province, southeastern China. Biodivers. Sci. 2015, 23, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Costello, E.K.; Schmidt, S.K. Microbial diversity in alpine tundra wet meadow soil: Novel Chloroflexi from a cold, water-saturated environment. Environ. Microbiol. 2006, 8, 1471–1486. [Google Scholar] [CrossRef]
- Chen, Y.P.; Rekha, P.D.; Arun, A.B.; Shen, F.T.; Lai, W.A.; Young, C.C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Kraut-Cohen, J.; Zolti, A.; Shaltiel-Harpaz, L.; Argaman, E.; Rabinovich, R.; Green, S.J.; Minz, D. Effects of tillage practices on soil microbiome and agricultural parameters. Sci. Total Environ. 2020, 705, 135791. [Google Scholar] [CrossRef] [PubMed]
- Hermans, S.M.; Buckley, H.L.; Case, B.S.; Curran-Cournane, F.; Taylor, M.; Lear, G. Bacteria as Emerging Indicators of Soil Condition. Appl. Environ. Microbiol. 2017, 83, e02826-16. [Google Scholar] [CrossRef] [Green Version]
- Sperlea, T.; Kreuder, N.; Beisser, D.; Hattab, G.; Boenigk, J.; Heider, D. Quantification of the covariation of lake microbiomes and environmental variables using a machine learning-based framework. Mol. Ecol. 2021, 30, 2131–2144. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-Based Assessment of Soil pH as a Predictor of Soil Bacterial Community Structure at the Continental Scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, K.; Wemheuer, B.; Korolkow, V.; Wemheuer, F.; Nacke, H.; Schoning, I.; Schrumpf, M.; Daniel, R. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Rep. 2016, 6, 33696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.J.; Zheng, Y.M.; Yan, J.X.; Li, H.J.; He, J.Z. Succession of plant and soil microbial communities with restoration of abandoned land in the Loess Plateau, China. J. Soils Sediments 2013, 13, 760–769. [Google Scholar] [CrossRef]
- Xian, W.; Zhang, X.; Li, W. Research status and prospect on bacterial phylum Chloroflexi. Acta Microbiol. Sin. 2020, 60, 1801–1820. [Google Scholar]
- Chase, J.M. Spatial scale resolves the niche versus neutral theory debate. J. Veg. Sci. 2014, 25, 319–322. [Google Scholar] [CrossRef]
- Feng, M.M.; Adams, J.M.; Fan, K.K.; Shi, Y.; Sun, R.B.; Wang, D.Z.; Guo, X.S.; Chu, H.Y. Long-term fertilization influences community assembly processes of soil diazotrophs. Soil Biol. Biochem. 2018, 126, 151–158. [Google Scholar] [CrossRef]
- Chen, W.M.; Jiao, S.; Li, Q.P.; Du, N.N. Dispersal limitation relative to environmental filtering governs the vertical small-scale assembly of soil microbiomes during restoration. J. Appl. Ecol. 2020, 57, 402–412. [Google Scholar] [CrossRef]
- Ettema, C.H.; Wardle, D.A. Spatial soil ecology. Trends Ecol. Evol. 2002, 17, 177–183. [Google Scholar] [CrossRef]
- Kong, W.B.; Wei, X.R.; Wu, Y.H.; Shao, M.G.; Zhang, Q.; Sadowsky, M.J.; Ishii, S.; Reich, P.B.; Wei, G.H.; Jiao, S.; et al. Afforestation can lower microbial diversity and functionality in deep soil layers in a semiarid region. Glob. Chang. Biol. 2022, 28, 6086–6101. [Google Scholar] [CrossRef]
- Jiao, S.; Lu, Y.H. Soil pH and temperature regulate assembly processes of abundant and rare bacterial communities in agricultural ecosystems. Environ. Microbiol. 2020, 22, 1052–1065. [Google Scholar] [CrossRef]
- Liao, H.K.; Zheng, C.L.; Li, J.; Long, J. Revegetation of a degraded karst ecosystem triggers shifts in diazotroph communities (composition, diversity, function and assembly processes). Appl. Soil Ecol. 2022, 176, 104479. [Google Scholar] [CrossRef]
- Jurburg, S.D.; Nunes, I.; Stegen, J.C.; Le Roux, X.; Prieme, A.; Sorensen, S.J.; Salles, J.F. Autogenic succession and deterministic recovery following disturbance in soil bacterial communities. Sci. Rep. 2017, 7, 45691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.Q.; Qiao, Y.C.; Wu, B.; Chen, H.; Li, W.; Jiang, N.; Zhang, X.L.; Liu, X.Z. Ecological Succession Pattern of Fungal Community in Soil along a Retreating Glacier. Front. Microbiol. 2017, 8, 1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B.H. Beyond biogeographic patterns: Processes shaping the microbial landscape. Nat. Rev. Microbiol. 2012, 10, 497–506. [Google Scholar] [CrossRef]
- Ferrenberg, S.; O’Neill, S.P.; Knelman, J.E.; Todd, B.; Duggan, S.; Bradley, D.; Robinson, T.; Schmidt, S.K.; Townsend, A.R.; Williams, M.W.; et al. Changes in assembly processes in soil bacterial communities following a wildfire disturbance. Isme J. 2013, 7, 1102–1111. [Google Scholar] [CrossRef] [Green Version]
- Dini-Andreote, F.; Stegen, J.C.; van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.Y.; Bao, J.B.; Wang, X.; Liu, Y.; Li, C.; Chai, B.F. Deterministic processes dominate soil microbial community assembly in subalpine coniferous forests on the Loess Plateau. PeerJ 2019, 7, e6746. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Li, Q.; Yan, J.H.; Liu, C.; Zhong, J.X. Vegetation restoration facilitates belowground microbial network complexity and recalcitrant soil organic carbon storage in southwest China karst region. Sci. Total Environ. 2022, 820, 153137. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, X.; Yao, K.; Zeng, F.; Zhang, C.; Zeng, Z.; Zhang, H. Diversity and Assembly of Bacteria Community in Lime Soil under Different Karst Land-Use Types. Forests 2023, 14, 672. https://doi.org/10.3390/f14040672
Zou X, Yao K, Zeng F, Zhang C, Zeng Z, Zhang H. Diversity and Assembly of Bacteria Community in Lime Soil under Different Karst Land-Use Types. Forests. 2023; 14(4):672. https://doi.org/10.3390/f14040672
Chicago/Turabian StyleZou, Xiaoxiao, Kai Yao, Fuping Zeng, Chen Zhang, Zhaoxia Zeng, and Hao Zhang. 2023. "Diversity and Assembly of Bacteria Community in Lime Soil under Different Karst Land-Use Types" Forests 14, no. 4: 672. https://doi.org/10.3390/f14040672