1. Introduction
Due to the high demand for agricultural activities, many lands were subjected to unattended activities. Land burning with aggressive shifting cultivation resulted in degraded soil, leading governments to maximize land usage in forest plantations [
1]. In Sabah, 19,058 ha were used for logging activities and 162,022 m
3 for timber harvesting in 2021. Around 370,130 ha of forest plantation area were acknowledged under various Sustainable Forest Management License Agreements (SFMLA) [
2].
The importance of forest plantations varied in many ways based on evaluation criteria [
3]. Most forest plantations were built on poor soils [
4], resulting from logging activity degrading the soil structure and decreasing plant growth [
5]. Nevertheless, marginal lands can provide ideal soil conditions with proper management [
6]. This is mainly needed to restore ecosystem services and productivity in degraded lands so that all species can perform well [
7].
At present,
Neolamarckia cadamba Roxb. Bosser and
Octomeles sumatrana Miq. have been introduced as a replacement for light hardwood as both species can grow in optimal environments, as was proved in early plantation trials by Sabah Forestry Department (SFD). In addition, both species are preferable choices for locations where timber species do not really regenerate [
8].
N. cadamba, also known as Kelempayan (Peninsular Malaysia and Sarawak), Laran (Sabah), Jabon (Kalimantan), or Kadam (India), is one of the most profit-oriented plantation species in Borneo (Sabah and Sarawak) and in some experimental plantings in Peninsular Malaysia supervised by the Forest Plantation Development Programme [
9].
N. cadamba belongs to the Rubiaceae family [
10]. On the other hand,
O. sumatrana, also known as Binuang, belongs to the Datiscaceae family and has straight boles like
N. cadamba [
11].
O. sumatrana can grow at an altitude of 0–600 m above sea level with mineral soils [
12]. Moreover, in field planting, root interaction can be very challenging [
13] because of methodological restrictions to measuring individual plant roots [
14]. Heterogenous soil conditions will affect competition with neighboring roots [
15]. Therefore, the present study was conducted on a small scale to measure and compare which species can survive in water-stress and water-logged conditions.
Both species are known as fast-growing species [
2]; comparing their root phenotype and growth is essential to assess their performance [
16].
N. cadamba is planted in different types of locations, wet and dry areas, regardless of the risk of failure because of its suitability to the negative effect of water-stress and water-logging on plant growth [
17].
O. sumatrana also has high adaptability to dry and moist areas, including two types of soil—clay and sandy clay—along the river banks [
18]. Although both are local species, scientific research is lacking regarding their early development in terms of root phenotyping, and water-stress and water-logged treatments. However, the information on this aspect is limited because of the challenges faced in field sampling—especially in root growth [
8]. Other root parameters that can be studied to compensate for the absence of data and have a better understanding of the survivability of these species: are root intensity (RI), specific root length (SRL), root length density (RLD) [
19], and root depth (RD).
Throughout this research, two indigenous species—
N. cadamba and
O. sumatrana—were used to determine if water-stress or water-logging treatments were suitable for their survival ability, mainly in terms of growth rate and root phenotyping. The application of water-stress and water-logging treatments influences the growth performance and root phenotyping of the seedlings of both species. Consequently, the adverse effects on plant growth, development, and seedling survival will restrict the species’ potential range. It is essential to understand how they will adapt to the current environment, including water-stress and water-logged conditions [
17]. Thus, the study compares the initial growth performance and below-ground parameters between
N. cadamba (White Laran) and
O. sumatrana (Binuang) under water-logging and water-stress conditions.
4. Discussion
The present study demonstrated that the growth of seedlings could be influenced by the different treatments received, including drought pressure, water-logged stress, and control conditions. Previous studies showed that the height and growth ability of trees could be influenced by water-stress [
24]. In addition, the average increment of
O. sumatrana’s height under water-stress conditions was far higher than the one observed in
N. cadamba, although both species are classified as fast-growing. This may be caused by phenotypic plasticity, defined as individuals’ ability to cope when exposed to different environmental conditions in terms of growth, survival, and even transformation [
25]. For
N. cadamba, the seedlings were affected by water-stress conditions, which decreased the plant’s ability to achieve an ideal height performance.
N. cadamba was said to be more adaptable in alluvial and moist areas, between periodically flooded areas, or in swamps, as soil moisture can influence the progressive growth of the plant [
26]. Besides, the soil media used for the experiments in this study might have influenced the seedlings’ growth in both species. A previous study using different media types demonstrated that the second-best medium for
N. cadamba’s germination was a mix of soil and sand [
27]. These conclusions are limited by time and methodology resources, and further experiments are needed to demonstrate that environmental conditions significantly impact the height of those trees.
In this study,
O. sumatrana showed a positive incline of RCD under water-stress. This result contradicted previous knowledge affirming that RCD can be larger if the plant is exposed to higher soil moisture, as plants can take more water than they need [
28]. Water-stress can have different impacts on the plant as it can lead to deleterious or adaptive changes [
29]. However, this study was conducted over a short time. A more extended period of water limitation can lead to more significant changes in the structure of plants. Water limitation can restrain cell growth, resulting in a loss of cell turgor, thus decreasing the diameter of newly-formed adventitious roots [
30]. Understanding water transport is crucial, as RCD can vary depending on water availability [
31]. The RCD of
N. cadamba rose steadily under the control treatment compared to the water-logged stress, following the knowledge that optimal water availability can increase water-use efficiency [
32]. Similarly, the RCD was not significantly reduced by water-logged stress, as previous studies demonstrated that water uptake by the roots is slower under water-logged treatment than under control conditions [
33].
No noteworthy difference was observed in the means of the LAI between all three conditions. The leaf area indicates drought tolerance and is essential in metabolic activities [
17]. Water-stress allows adaptive action for immediate survival and structural plants’ ability to alter their gene expression, thus allowing improved functioning [
29].
O. sumatrana had an apparent increase of LAI under water-stress due to a reduction in the number of leaves produced but not in the total leaf surface. The capability to increase leaf area and the development of the leaf area itself were seemingly under genetic control [
34]. The highest mean of LAI in
N. cadamba was recorded under control treatment, probably due to seedlings producing leaves quickly because there was no water restriction [
35]. Water-stress treatment in
N. cadamba led to slightly lower mean values than in
O. sumatrana because small leaves endure turgor pressure better than larger leaves and can significantly contribute to turgor maintenance under stress conditions [
17]. The leaf growth changes caused by the deficiency of nutrients were well-demonstrated throughout these findings. Nutrient leaching occurs by the loss of water-soluble plain nutrients from the soil because of rainfall, except for water-stress treatment since samples were covered by transparent plastic. The observed positive effect could be sustained as the growth in all live individuals was stagnant [
36]. Therefore, the results observed for LAI do not necessarily mean that the growth is not optimal, as other parameters also influence growth performance.
Results obtained from the analysis of dried shoot biomass showed that the mean value at 8 WAT and 12 WAT was impacted by water-logged treatment. The shoot biomass is closely related to roots. The shoots’ dry weight reflects the roots’ capability to support the DSB in maintaining plant structure and form and absorbing nutrients and water [
37]. This is because roots play a significant role in the uptake of nutrients and water while providing support and anchorage to plants [
38]. Soil water availability is one of the features that restrict the production of plants [
39]. Water and nutrients can be absorbed efficiently when the root density is greater, which increases shoot biomass [
19]. The higher the moisture content in soil due to water availability, the more root growth can be improved, thus improving plant growth [
39]. Both species recorded their lowest reading at both harvest stages when submitted to a water-stress treatment. Water restriction leads to a deficit in the number of leaves per plant and green leaves present and a decrease in leaf longevity depending on the soil’s water potential [
17]. A stressful environment can be the principal reason that prevents maximal growth and biomass production [
25].
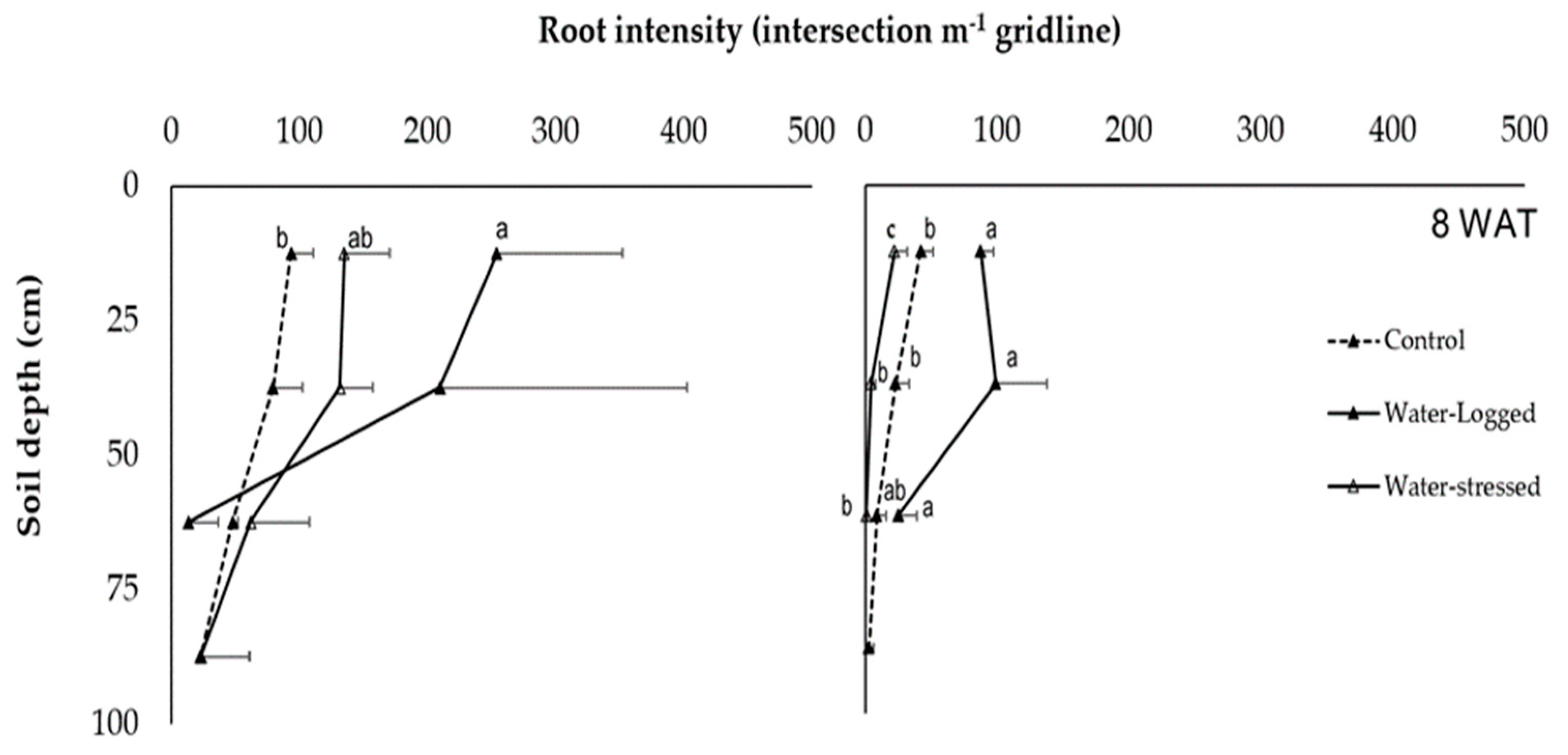
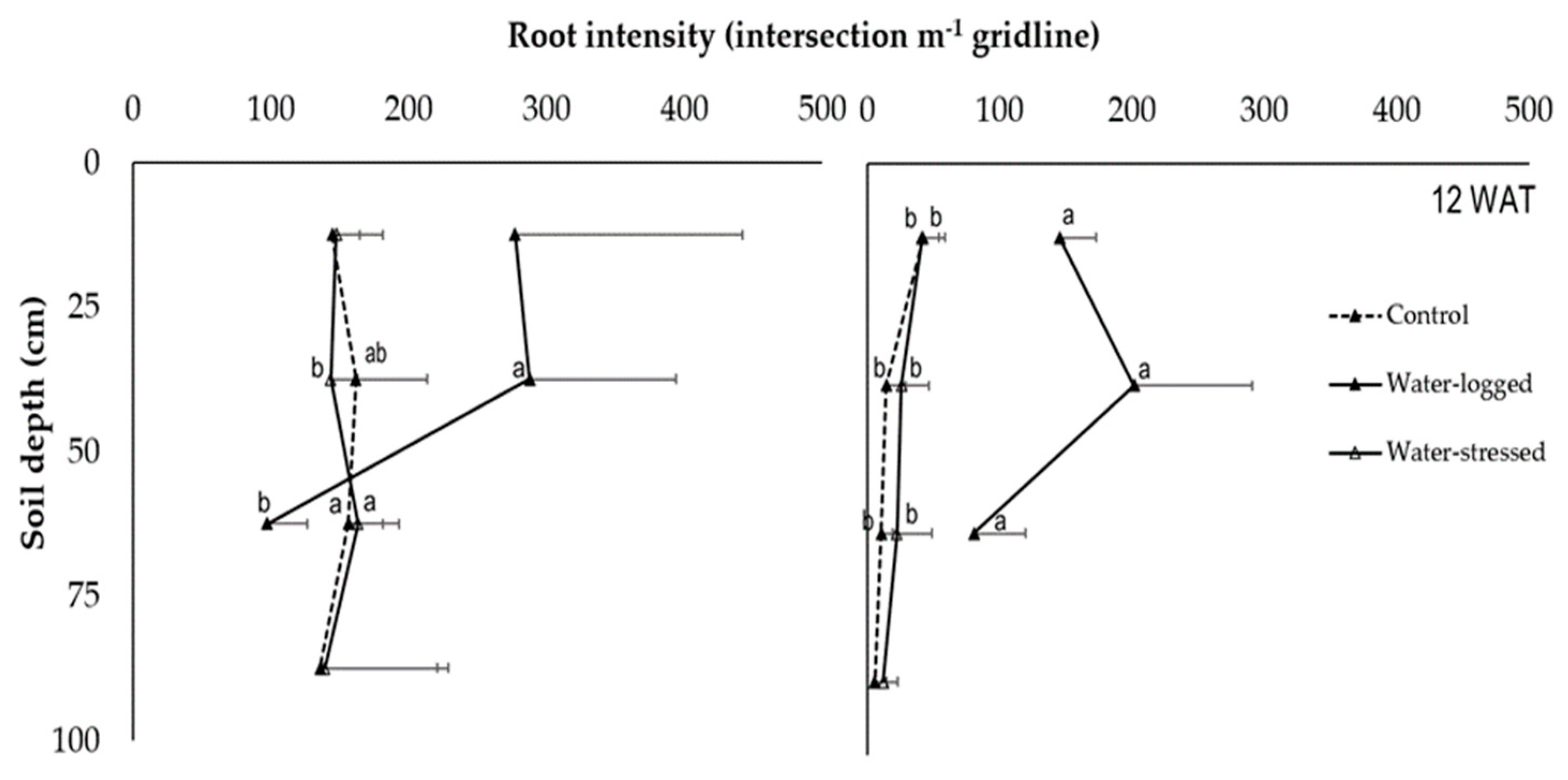
As for RD and RI, the data were presented according to roots visible at the rhizotron interface (
Table 6,
Figure 2,
Figure 3 and
Figure 4). Genetic and environmental conditions generally influence root depth. Root depth could be a significant feature in species in addition to their root biomass, as roots play an essential role in water uptake and nutrient absorption and can accommodate the shape of plants by hydraulic lift (HL). Additionally, the soil-water relationship is interconnected with the rooting condition, as the existing variation of optimal rooting depths depends on the species and its capability to adapt to various soil conditions [
40]. Based on the results, different root depths can be seen, with both species thriving excellently in particular conditions.
N. cadamba root depth generally approached 80 cm, demonstrating that this species’ root system is adaptive to water restriction pressure. A root system capable of penetrating deeper into the soil to extract water from deeper layers is an indispensable criterion for drought resistance. Its impact on the survival ability of the plant is significant as it allows efficient water use under drought stress [
30].
O. sumatrana prefers to penetrate high moisture of soil, approaching 65 cm of depth, which is almost 80% of
N. cadamba’s root depth. Under the water-stress condition, as root length is reduced due to hypoxia [
33], roots approach the third layer of soil (50–75 cm). Besides, it is believed that
N. cadamba and
O. sumatrana could not have more root extensions when approaching the deepest soil layer (75–100 cm) due to water clogging. This observation was supported by a previous study demonstrating that constant flooding leads to unsuitable conditions for
O. sumatrana to grow [
2]. Another study also stated that some trees of
N. cadamba in forest plantation areas, which have continuous access to excessive water, resulted in aboveground disadvantages and poor root systems [
9].
The rhizotron is one of the methods used to calculate the RI through two-dimensional root imaging (captured image), also known as a non-destructive method [
41]. The mean value for RI for both species at 75–100 cm was the lowest under water-logged treatment. The same condition applied to different species also showed that root development was severely restricted under complete water-logged treatment. The finding showed that roots could have different functions and growth if the plant survives under water-logged treatment as the roots go into hypoxia [
42]. That is why the RI mean value gap between water-logged treatment and control treatment is slightly different; both species can survive in an environment with high soil moisture content but not in a fully clogged environment in the long term [
17]. Usually, the growth rate of roots is gravely decreased when plants undergo water-logged treatment [
43]. Therefore, root traits significantly improve the relationship between below-ground activities and plant growth [
41].
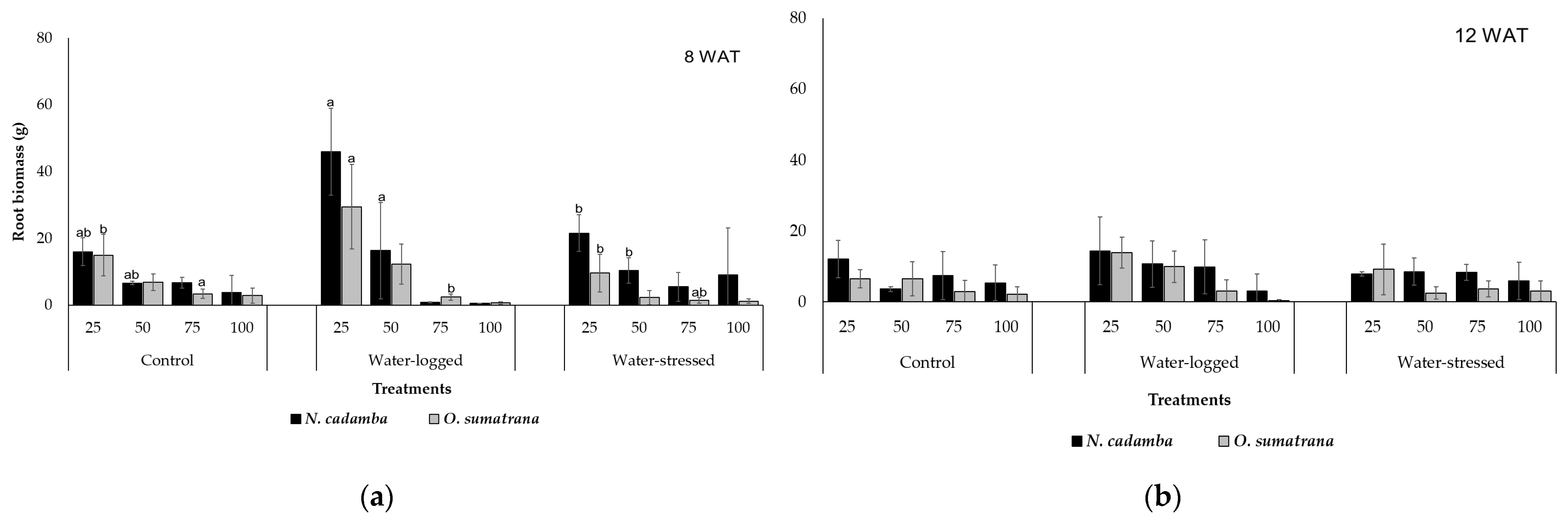
In each treatment, root biomass production was affected to a different extent. Through the analysis of both species in the first and second harvests under water-stress, water-logged, and control treatment, the RB in the upper layer (0–25 cm) and the second layer of soil (25–50 cm) presented the highest average of RB. On the other hand, in soil depths of 50–75 cm and 75–100 cm, only a low amount of root biomass was produced—about one-third of the root mass growing in the upper layer. In the third and fourth depths of the soil, a root biomass reduction is expected at the end of the harvesting stage. Indeed, root respiration decreases as water-logging produces delayed effects and can damage the root biomass [
43]. The higher the depth of the soil and, thus, the water content, the lower the root biomass production [
44]. Low root biomass production is a primary element that significantly impacts growth patterns and water relations [
33]. Hypoxic or anoxic conditions inhibit root growth, making their primary purpose difficult [
45]. On the other hand, roots need to absorb more water in the first and second layers of soil; soil moisture must be maintained to support the distribution of roots. In addition, higher root biomass in upper soil layers is caused by a lower soil water content at the surface than in subsurface layers [
44].
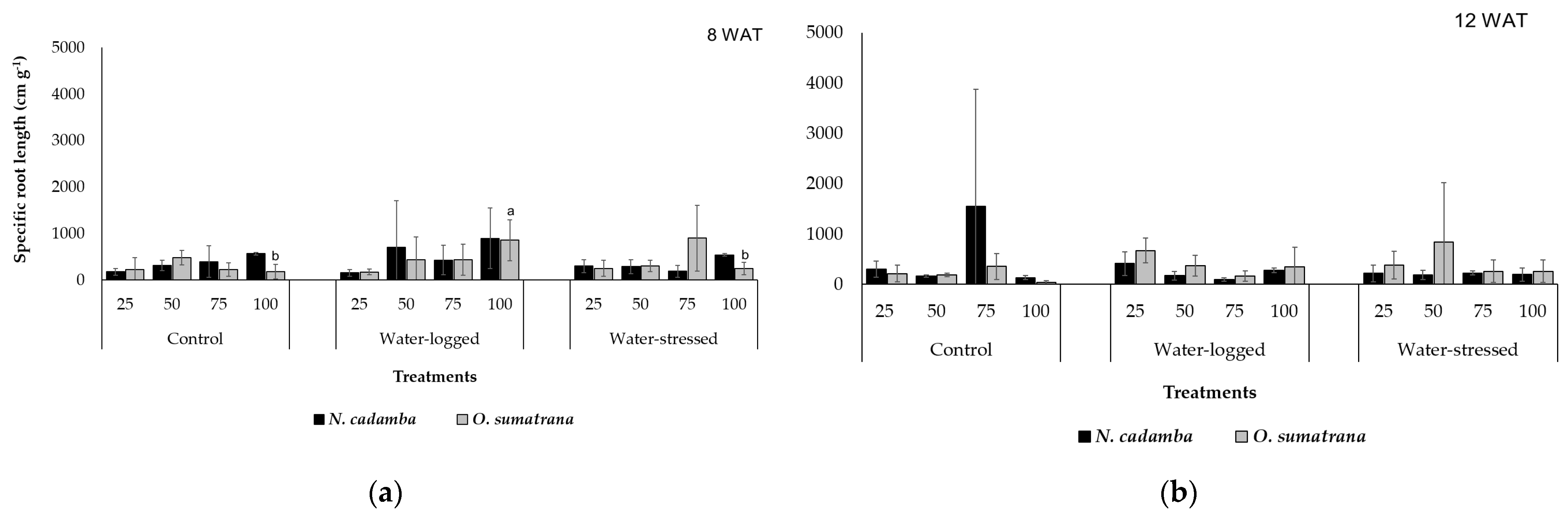
Generally, the SRL observed in
N. cadamba was higher than that of
O. sumatrana. SRL is the root length ratio to fine roots’ dry mass [
19]. SRL increases simultaneously with RI [
46]. SRL is a complex parameter that includes variations in root diameter and root tissue density [
47]. At 8 WAT,
N. cadamba and
O. sumatrana presented their highest SRL under water-logged treatment, which increases the root diameter [
43].
N. cadamba, at the second harvest stage, presented its highest SRL under the control treatment. It was argued that roots in normal conditions are more productive in deeper soils since roots often occupy shallow layers [
48]. A different situation was observed with
O. sumatrana at 12 WAT, with a steady increase under water-stress treatment. Root elongation is necessary when plants are grown in conditions with restricted supplies of water in the soil [
49]. A greater SRL is obtained by having a smaller diameter of roots, and reaching water under water-stress requires an increase in root length or root hair density and a decrease in plant carbon investment [
50]. Thus, the analysis must be emphasized by a better understanding of drought adaptation mechanisms, as roots are the least affected compared to vegetative, reproductive, and aerial organs [
30].
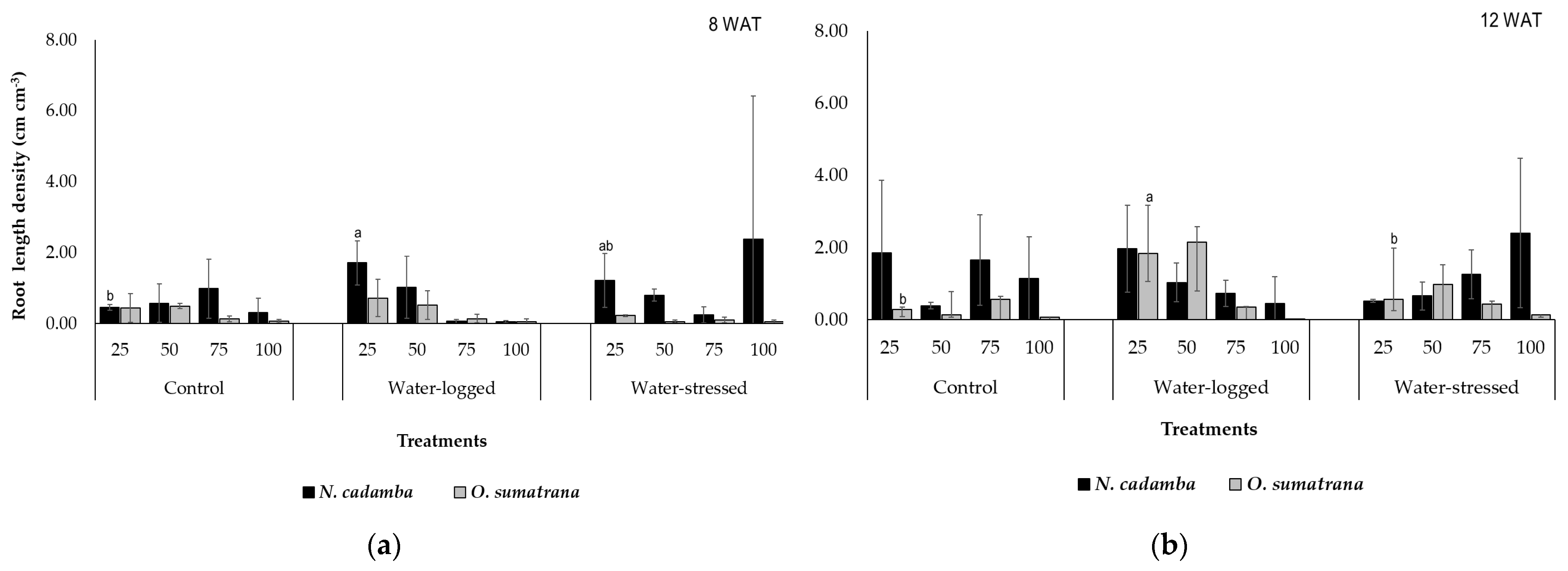
RLD is essential as it was used to evaluate the root system by exploring the soil in search of nutrients and water [
51]. A low RLD was observed in
O. sumatrana in the fourth layer of soil (75–100 cm) under the water-logged treatment because compacted soil is more prone to a complete water-logged condition where hypoxia is likely to impede root growth [
49]. Indeed the lowest RLD was observed in the third and fourth layers of soil (50–100 cm). A previous study demonstrated that if the root system is exposed to flooding, the mortality rate of the plant increases [
52]. However,
O. sumatrana recorded its highest RLD at the second layer of soil under the water-logged treatment compared to other treatments. Water from rainfall is usually kept deep in the soil, and the rapid growth of roots penetrating through the soil increases the capture of water and nitrogen [
53]. Roots can change based on different soil depths depending on the availability of nutrients and water [
54]. RLD can drastically increase, and roots can spread in the upper layer of soil that is not water-logged [
42]. The higher RLD of
N. cadamba observed under the water-stress treatment at 12 WAT in the third layer of soil (50–75 cm) is assumed to be a potential adaptation feature [
55]. Roots are more present in drought conditions where plants try to survive, showing their capability to adapt to marginal lands. Exploring root increase has considerable potential in genetic research to optimize root system architecture (RSA) in water-stress conditions [
56]. Besides, the mean value was slightly higher at a soil depth of 50–75 cm compared to the upper layers. Nevertheless, a previous study suggested that the reallocation of carbon under water-stress impacts root growth, decreasing RLD in upper layers [
51]. Roots tend to penetrate deep soil because of the limited water availability in top soil layers [
57]. This phenomenon is part of xerotropism, where the root gravity response is enhanced in response to desiccation [
58].

 and
and 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}