
Biocontrol Potential of Entomopathogenic Nematodes against Odontotermes obesus (Blattodea: Termitidae) under Laboratory and Field Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Culture
2.1.1. Termites
2.1.2. Entomopathogenic Nematodes
2.2. Laboratory Bioassays
Filter Paper Bioassay
2.3. Sawdust Bioassay
2.4. Field Bioassay
2.5. Statistical Analysis
3. Results
3.1. Mortality in Laboratory Trials
3.1.1. Bioassays (B)
3.1.2. Nematode Species (S)
3.1.3. Nematode Concentrations (C)
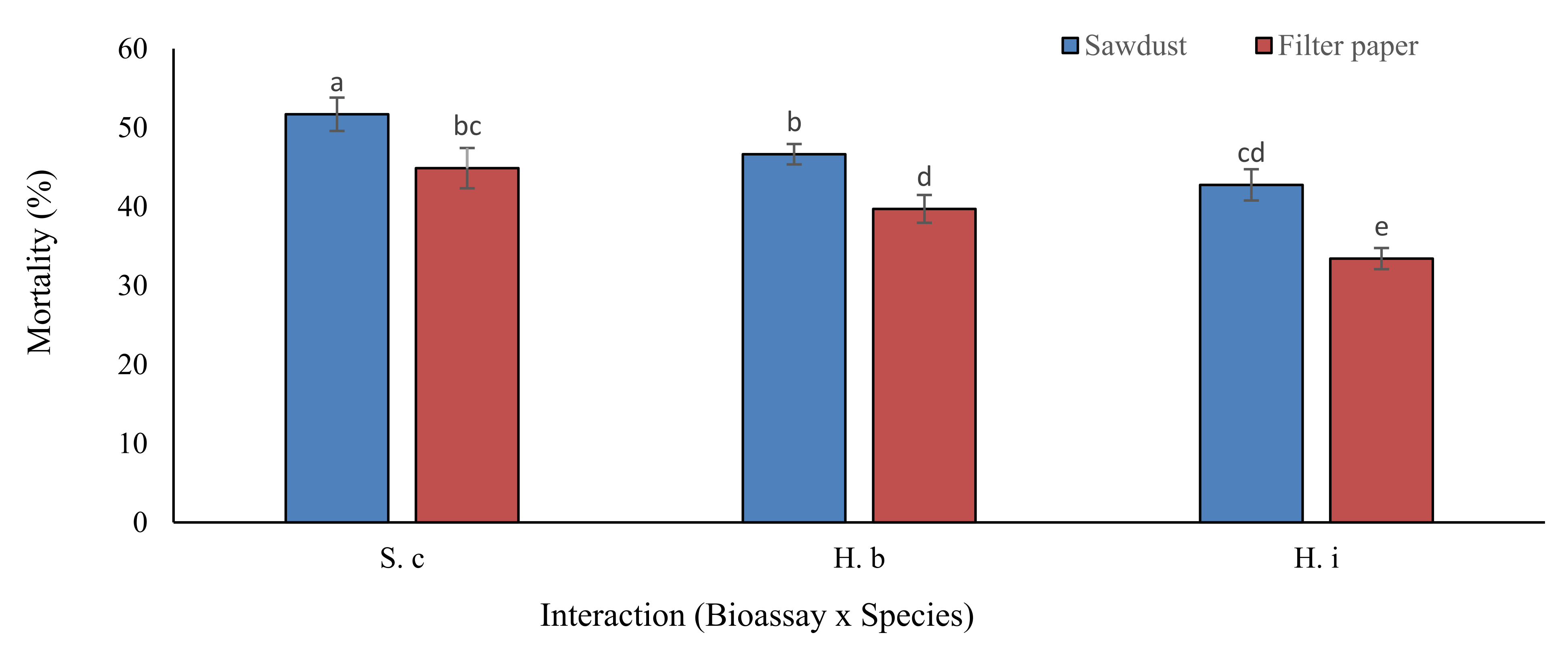
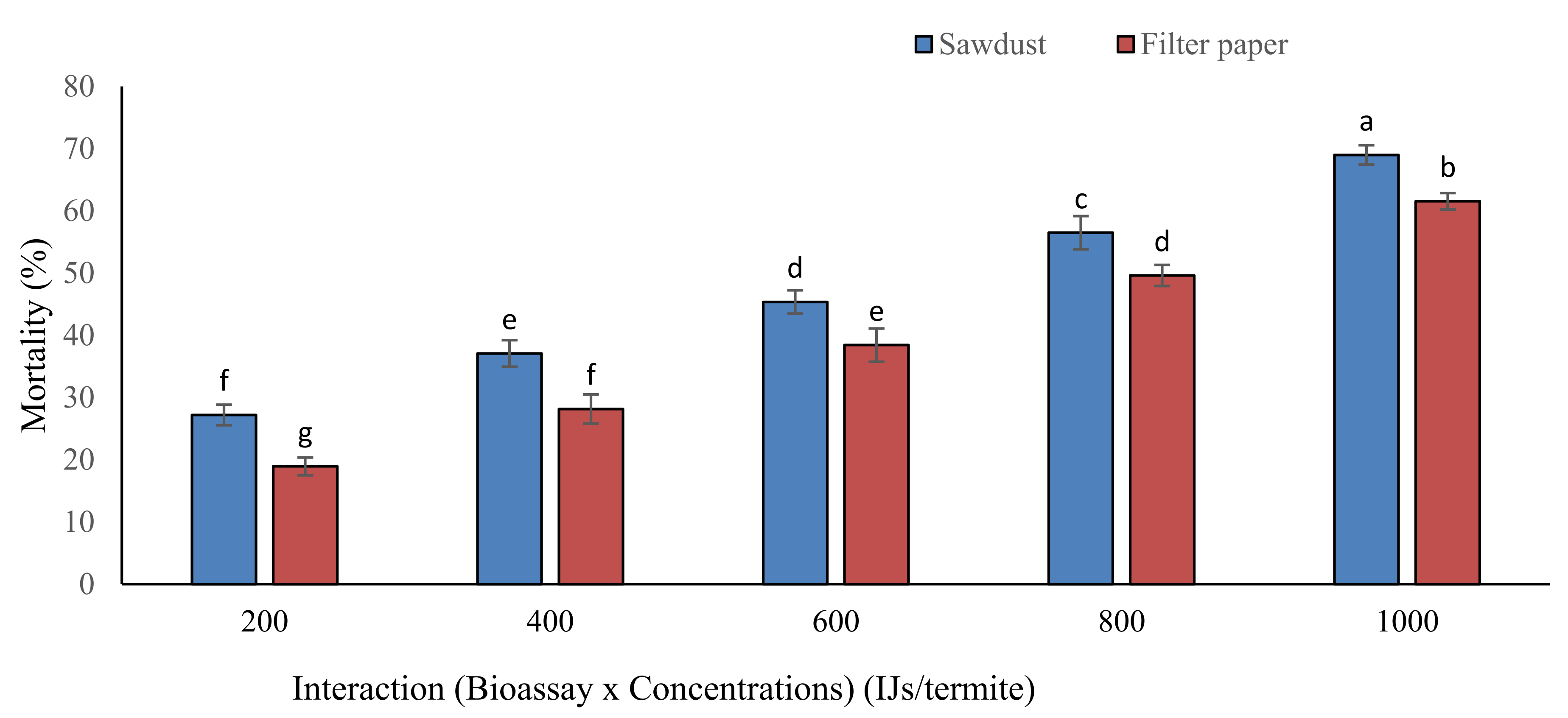
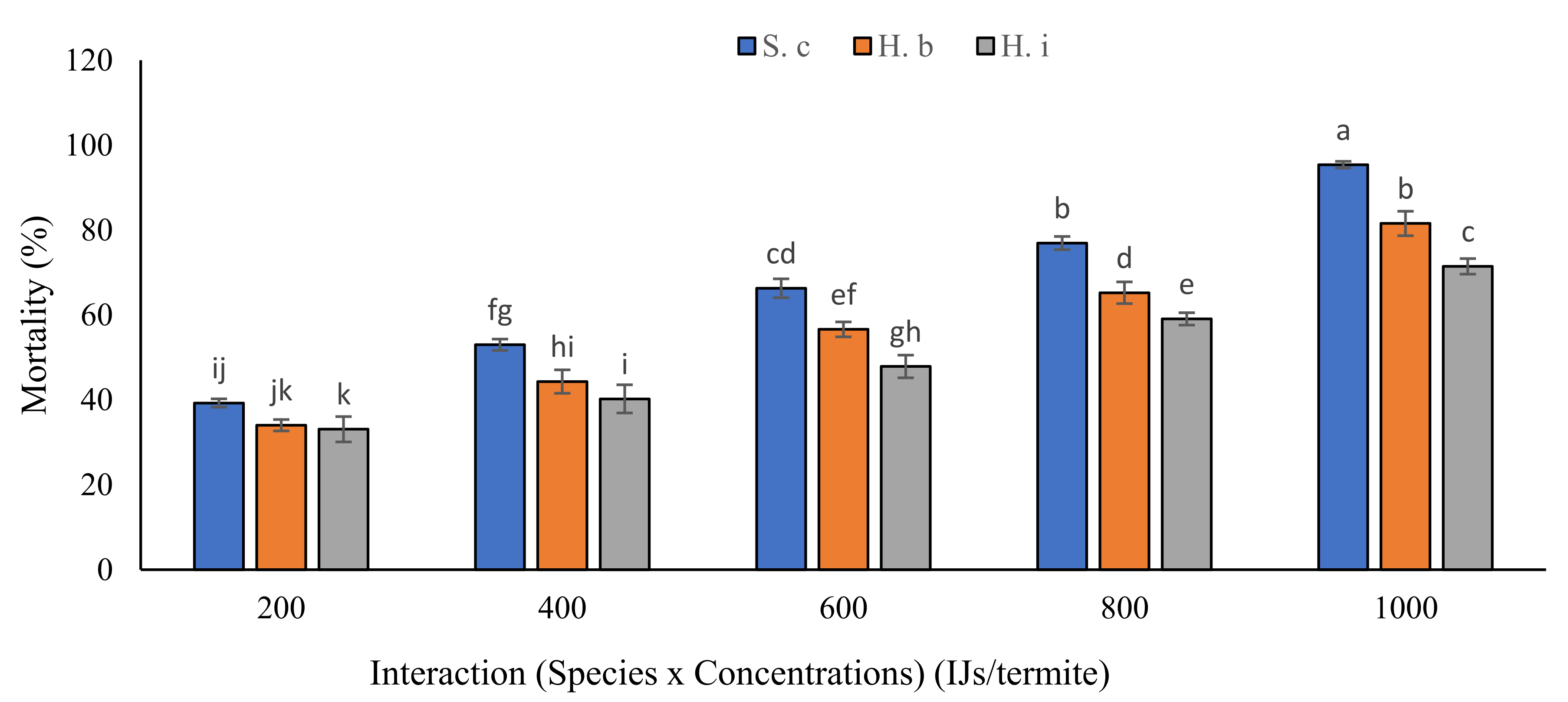
3.1.4. Interactions of Bioassay (B), Nematode Species (S) and Their Concentrations (C)
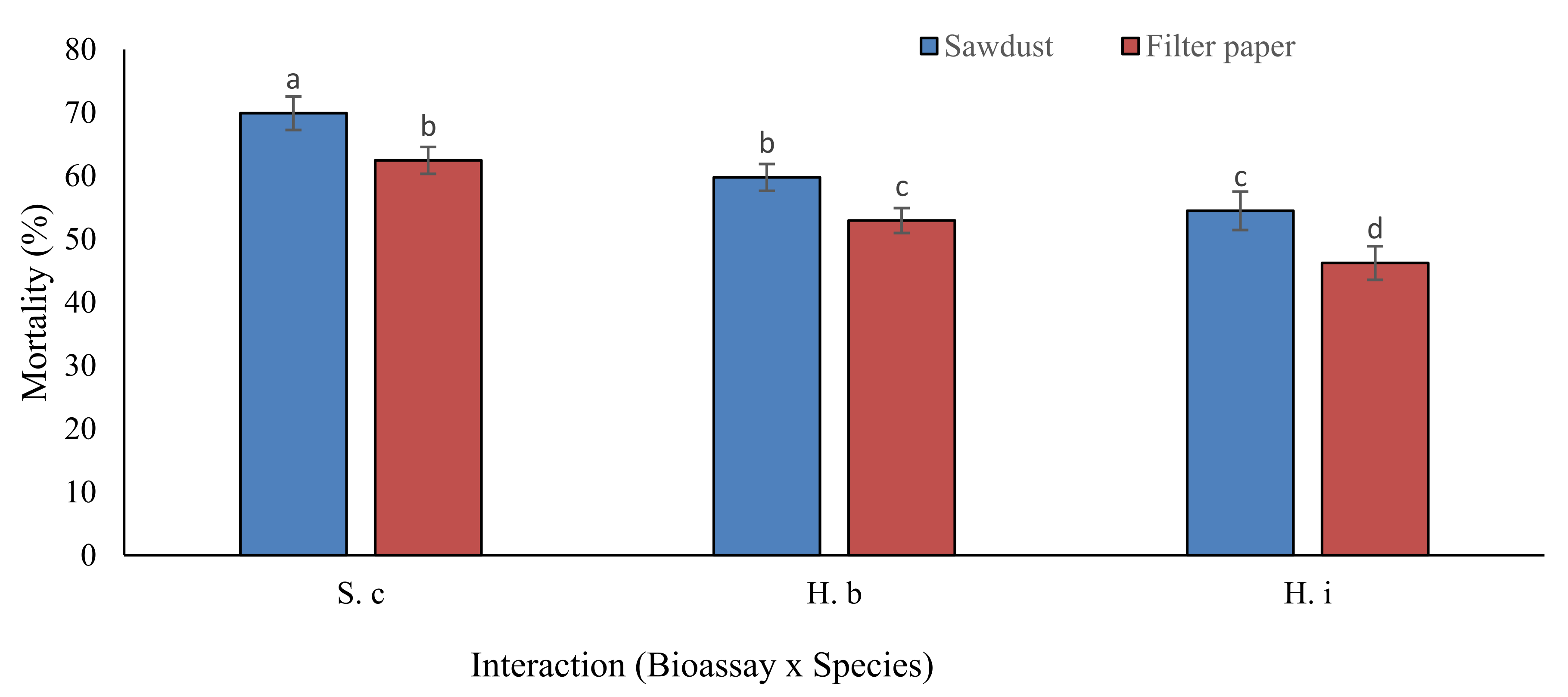
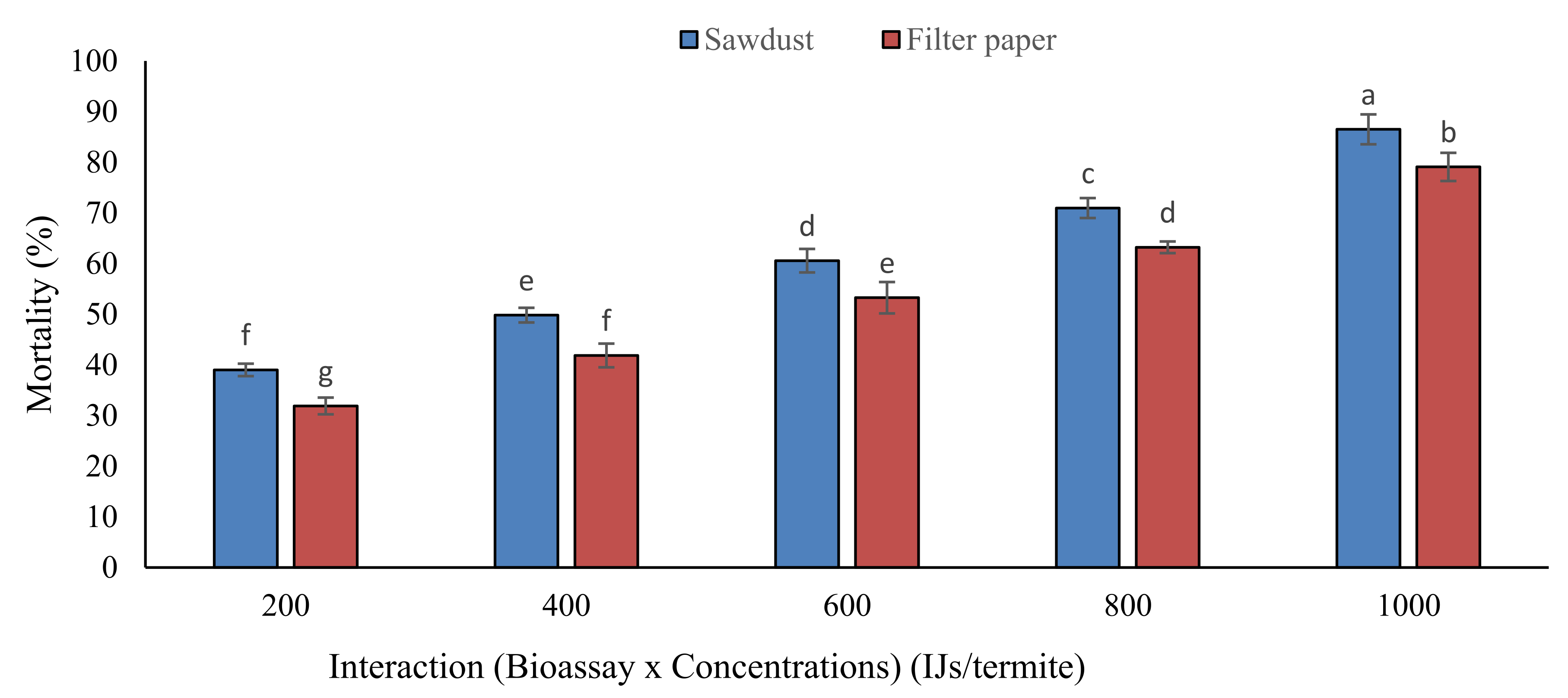
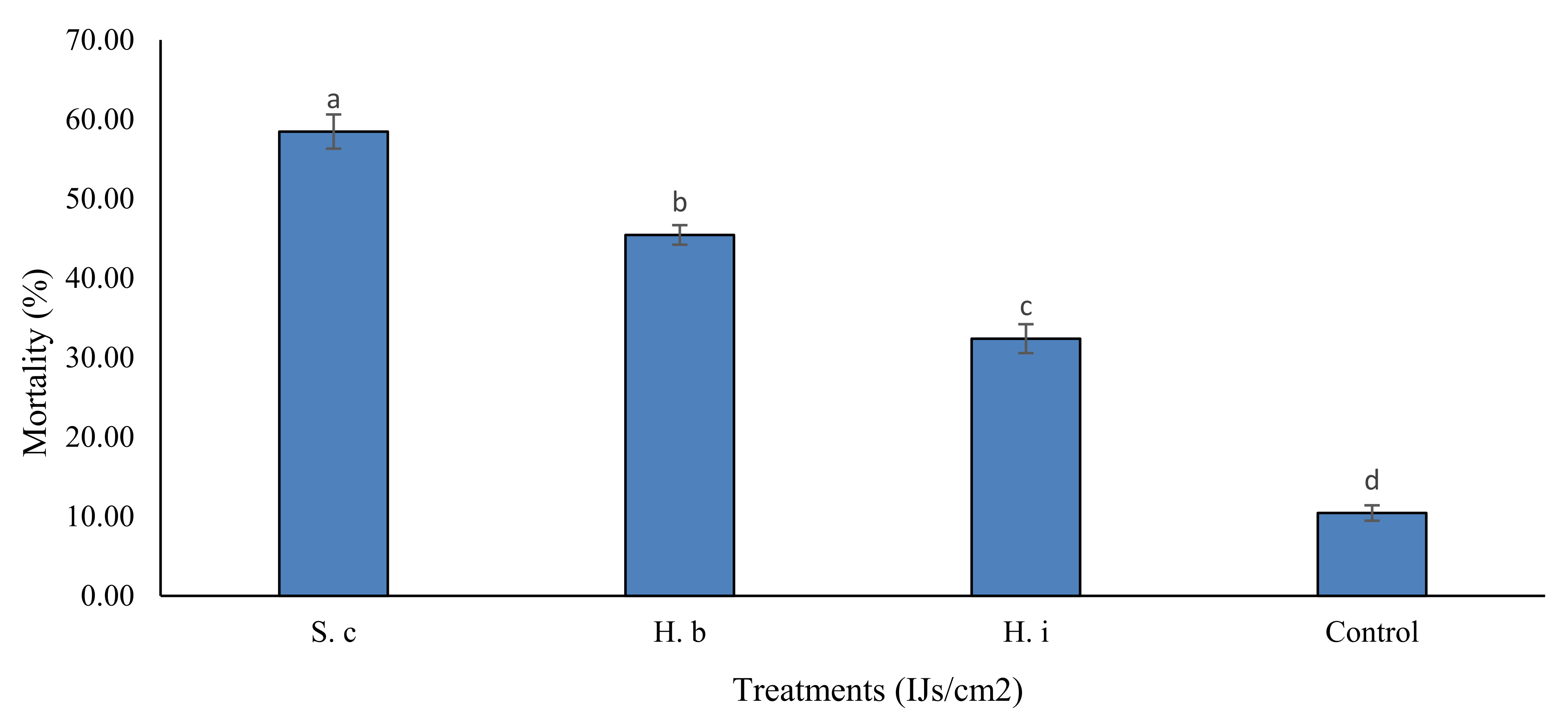
3.2. Mortality in Field Trials
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hickey, C.D. Effects of Disodium Octaborate Tetrahydrate in Ethylene Glycol on Consumption and Mortality of the Eastern Subterranean Termite. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2006. [Google Scholar]
- Ahmed, B.; French, J. An overview of termite control methods in Australia and their link to aspects of termite biology and ecology. Pak. Entomol. 2008, 30, 101–118. [Google Scholar]
- Krishna, K.; Grimaldi, D.A.; Krishna, V.; Engel, M.S. Treatise on the Isoptera of the World: Neoisoptera Excluding Termitidae. Bull. Am. Mus. Nat. Hist. 2013, 2013, 623–973. [Google Scholar] [CrossRef]
- Khan, I.A.; Zaman, M.; Akbar, R.; Ali, I.; Alam, M.; Saeed, M.; Farid, A. Efficacy of single and mixed particles sand size as physical barrier against Heterotermes indicola under laboratory conditions. J. Entomol. Zool. Stud. 2015, 3, 106–109. [Google Scholar]
- Jouquet, P.; Traoré, S.; Choosai, C.; Hartmann, C.; Bignell, D. Influence of termites on ecosystem functioning. Ecosystem services provided by termites. Eur. J. Soil Biol. 2011, 47, 215–222. [Google Scholar] [CrossRef]
- Govorushko, S. Economic and ecological importance of termites: A global review. Entomol. Sci. 2019, 22, 21–35. [Google Scholar] [CrossRef]
- Ahmad, F.; Fouad, H.; Liang, S.Y.; Hu, Y.; Mo, J.C. Termites and Chinese agricultural system: Applications and advances in integrated termite management and chemical control. Insect Sci. 2021, 28, 2–20. [Google Scholar] [CrossRef]
- Uys, V. A Guide to the Termite Genera of Southern Africa; Plant Protection Research Institute, Agricultural Research Council: Pretoria, South Africa, 2002. [Google Scholar]
- Sunitha, S.; Miranda, M. Population dynamics of termites with special reference to Odontotermes obesus (Desneux). Asian J. Anim. Sci. 2011, 6, 43–45. [Google Scholar]
- Rasib, K.Z.; Ashraf, H.; Afzal, M. Feeding preferences of Odontotermes obesus (Rambur) (Isoptera: Termitidae) on different commercial and non-commercial woods from Lahore, Pakistan, under laboratory and field conditions. Zool. Ecol. 2014, 24, 369–379. [Google Scholar] [CrossRef]
- Mandal, B.; Bashar, K.; Howlader, A.; Rahman, K. Incidence of termite infestation to tree species in Jahangirnagar University Campus, Bangladesh. Bangladesh J. Life Sci. 2010, 22, 7–15. [Google Scholar]
- Mugerwa, S.; Nyangito, M.; Mpairwe, D.; Bakuneeta, C.; Nderitu, J.; Zziwa, E. Termite assemblage structure on grazing lands in semi-arid Nakasongola. Agric. Biol. J. N. Am. 2011, 2, 848–859. [Google Scholar] [CrossRef]
- Rao, A.N.; Mahapatro, G.; Chatterjee, D. Termites as structural pest: Status in Indian Scenario. Proc. Natl. Acad. Sci. USA 2018, 88, 977–994. [Google Scholar]
- Wagner, T. US Forest Service termiticide tests. Sociobiology 2003, 41, 131–141. [Google Scholar]
- Haq, I.U.; Idrees, A.; Abbasi, A.; Ali, S.; Asad, M.; Li, C.; Liu, C.-Z.; Zhang, K.-X.; Yasin, M.; Asghar, M.A.; et al. Silicon Accumulation in Maize and its Effects on Demographical Traits of Fall armyworm, [Spodoptera frugiperda (J. E. Smith)]. Silicon 2022, 14, 1–13. [Google Scholar] [CrossRef]
- Al-Zaidawi, J.B.; Karimi, J.; Mahdikhani Moghadam, E. Entomopathogenic nematodes as potential biological control agents of Subterranean termite, Microcerotermes diversus (Blattodea: Termitidae) in Iraq. Environ. Entomol. 2020, 49, 412–421. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef] [Green Version]
- Delcour, I.; Spanoghe, P.; Uyttendaele, M. Literature review: Impact of climate change on pesticide use. Food Res. Int. 2015, 68, 7–15. [Google Scholar] [CrossRef]
- Ciancio, A.; Pieterse, C.M.; Mercado-Blanco, J. (Eds.) Harnessing Useful Rhizosphere Microorganisms for Pathogen and Pest Biocontrol; Frontiers Media SA: Lausanne, Switzerland, 2019; Volume 10, p. 1935. [Google Scholar]
- Ambele, F.C.; Bisseleua Daghela, H.; Babalola, O.O.; Ekesi, S. Soil-dwelling insect pests of tree crops in Sub-Saharan Africa, problems and management strategies—A review. J. Appl. Entomol. 2018, 142, 539–552. [Google Scholar] [CrossRef]
- de Oliveira Giannasi, A.; Roque Brambila, C.; Zart, M.; Guide, B.A.; Alves, V.S. Assessment of entomopathogenic nematodes in Agrotis ipsilon (Lepidoptera: Noctuidae) under laboratory and greenhouse conditions. Rev. Colomb. Entomol. 2018, 44, 25–31. [Google Scholar] [CrossRef]
- Viteri, D.M.; Linares, A.M.; Flores, L. Use of the entomopathogenic nematode Steinernema carpocapsae in combination with low-toxicity insecticides to control fall armyworm (Lepidoptera: Noctuidae) larvae. Fla. Entomol. 2018, 101, 327–329. [Google Scholar] [CrossRef] [Green Version]
- Laznik, Ž.; Tóth, T.; Lakatos, T.; Vidrih, M.; Trdan, S. Control of the Colorado potato beetle (Leptinotarsa decemlineata [Say]) on potato under field conditions: A comparison of the efficacy of foliar application of two strains of Steinernema feltiae (Filipjev) and spraying with thiametoxam. J. PlantDis. Prot. 2010, 117, 129–135. [Google Scholar] [CrossRef]
- Kepenekci, I.; Hazir, S.; Özdem, A. Evaluation of native entomopathogenic nematodes for the control of the European cherry fruit fly Rhagoletis cerasi L. (Diptera: Tephritidae) larvae in soil. Turk. J. Agric. For. 2015, 39, 74–79. [Google Scholar] [CrossRef]
- Yuksel, E.; Taskesen, Y.E.; Erarslan, D.; Canhilal, R. Effectiveness of different entomopathogenic nematode species against the variegated cutworm, Peridroma saucia (Hubner) (Lepidoptera: Noctuidae). Egypt. J. Biol. Pest Control 2018, 28, 8. [Google Scholar] [CrossRef] [Green Version]
- Acharya, R.; Yu, Y.-S.; Shim, J.-K.; Lee, K.-Y. Virulence of four entomopathogenic nematodes against the tobacco cutworm Spodoptera litura Fabricius. Biol. Control 2020, 150, 104348. [Google Scholar] [CrossRef]
- Bhat, A.H.; Thanwisai, A.; Muangpat, P.; Meesil, W.; Janthu, P.; Dumidae, A.; Subkrasae, C.; Ardpairin, J.; Tandhavanant, S.; Yoshino, T.P.; et al. Entomopathogenic Nematodes and Their Symbiotic Bacteria from the National Parks of Thailand and Larvicidal Property of Symbiotic Bacteria against Aedes aegypti and Culex quinquefasciatus. Biology 2022, 11, 1658. [Google Scholar]
- Adams, B.J.; Fodor, A.; Koppenhöfer, H.S.; Stackebrandt, E.; Stock, S.P.; Klein, M.G. Reprint of “Biodiversity and systematics of nematode–bacterium entomopathogens”. Biol. Control 2006, 38, 4–21. [Google Scholar] [CrossRef]
- Stuart, R.J.; Barbercheck, M.E.; Grewal, P.S. Entomopathogenic nematodes in the soil environment: Distributions, interactions and the influence of biotic and abiotic factors. In Nematode Pathogenesis of Insects and Other Pests; Springer: Berlin/Heidelberg, Germany, 2015; pp. 97–137. [Google Scholar]
- Campos-Herrera, R.; Martín, J.A.R.; Escuer, M.; García-González, M.T.; Duncan, L.W.; Gutiérrez, C. Entomopathogenic nematode food webs in an ancient, mining pollution gradient in Spain. Sci. Total Environ. 2016, 572, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Kergunteuil, A.; Campos-Herrera, R.; Sánchez-Moreno, S.; Vittoz, P.; Rasmann, S. The abundance, diversity, and metabolic footprint of soil nematodes is highest in high elevation alpine grasslands. Front. Ecol. Evol. 2016, 4, 84. [Google Scholar] [CrossRef] [Green Version]
- Kaya, H.K.; Aguillera, M.; Alumai, A.; Choo, H.Y.; De la Torre, M.; Fodor, A.; Ganguly, S.; Hazır, S.; Lakatos, T.; Pye, A. Status of entomopathogenic nematodes and their symbiotic bacteria from selected countries or regions of the world. Biol. Control 2006, 38, 134–155. [Google Scholar] [CrossRef]
- Griffin, C.; Boemare, N.; Lewis, E.; Grewal, P.; Ehlers, R.; Shapiro-Ilan, D. (Eds.) Nematodes as Biocontrol Agents; CABI: Wallingford, UK, 2005. [Google Scholar]
- Haq, I.U.; Ashraf, H.J.; Abbasi, A.; Majid, M.; Inayat, R. Integration of silicon with entomopathogenic nematode against fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). In Proceedings of the 4th International Conference on Applied Zoology-2021 (ICAZ-2021), Islamabad, Pakistan, 24–25 November 2021. [Google Scholar]
- Stock, S.P.; Kusakabe, A.; Orozco, R.A. Secondary Metabolites Produced by Symbionts and Their Application in Agriculture: What We Know and What to Do Next. J. Nematol. 2017, 49, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Dillman, A.R.; Lima, S.; Bandi, C.; Wanji, S.; Diagne, M.; Casiraghi, M. An entomopathogenic nematode by any other name. PLoS Pathog. 2012, 223, 18. [Google Scholar] [CrossRef] [Green Version]
- Stock, S.P. Diversity, biology and evolutionary relationships. In Nematode Pathogenesis of Insects and Other Pests; Springer: Berlin/Heidelberg, Germany, 2015; pp. 3–27. [Google Scholar]
- Mauldin, J.K.; Beal, R.H. Entomogenous nematodes for control of subterranean termites, Reticulitermes spp. (Isoptera: Rhinotermitidae). J. Econ. Entomol. 1989, 82, 1638–1642. [Google Scholar] [CrossRef]
- Wang, C.; Powell, J.E.; Nguyen, K. Laboratory evaluations of four entomopathogenic nematodes for control of subterranean termites (Isoptera: Rhinotermitidae). Environ. Entomol. 2002, 31, 381–387. [Google Scholar] [CrossRef]
- Baker, N.R.; Hydar, B.; Gowen, S. Reproduction of entomopathogenic nematodes Steinernema carpocapsae and Heterorhabditis bacteriophora on the German cockroach Blatella germanica at different temperatures. Iraqi J. Sci. 2012, 53, 505–512. [Google Scholar]
- Epsky, N.D.; Capinera, J.L. Efficacy of the entomogenous nematode Steinernema feltiae against a subterranean termite, Reticulitermes tibialis (Isoptera: Rhinotermitidae). J. Econ. Entomol. 1988, 81, 1313–1317. [Google Scholar] [CrossRef]
- Guo, W.; Yan, X.; Zhao, G.; Han, R. Efficacy of entomopathogenic Steinernema and Heterorhabditis nematodes against white grubs (Coleoptera: Scarabaeidae) in peanut fields. J. Econ. Entomol. 2013, 106, 1112–1117. [Google Scholar] [CrossRef] [PubMed]
- Caccia, M.G.; Del Valle, E.; Doucet, M.E.; Lax, P. Susceptibility of Spodoptera frugiperda and Helicoverpa gelotopoeon (Lepidoptera: Noctuidae) to the entomopathogenic nematode Steinernema diaprepesi (Rhabditida: Steinernematidae) under laboratory conditions. Chil. J. Agric. Res. 2014, 74, 123–126. [Google Scholar] [CrossRef]
- Acharya, R.; Hwang, H.-S.; Shim, J.-K.; Yu, Y.-S.; Lee, K.-Y. Control efficacy of fungus gnat, Bradysia impatiens, enhanced by a combination of entomopathogenic nematodes and predatory mites. Biol. Control 2019, 138, 104071. [Google Scholar] [CrossRef]
- Platt, T.; Stokwe, N.; Malan, A. A review of the potential use of entomopathogenic nematodes to control above-ground insect pests in South Africa. S. Afr. J. Enol. Vitic. 2020, 41, 1–16. [Google Scholar] [CrossRef]
- Yan, X.; Shahid Arain, M.; Lin, Y.; Gu, X.; Zhang, L.; Li, J.; Han, R. Efficacy of entomopathogenic nematodes against the tobacco cutworm, Spodoptera litura (Lepidoptera: Noctuidae). J. Econ. Entomol. 2020, 113, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, J.; Han, X.; Moens, M. Effect of temperature on the pathogenicity of entomopathogenic nematodes (Steinernema and Heterorhabditis spp.) to Delia radicum. BioControl 2003, 48, 713–724. [Google Scholar] [CrossRef]
- Aydin, H.; Susurluk, A. Competitive abilities of the entomopathogenic nematodes Steinernema feltiae and Heterorhabditis bacteriophora in the same host at different temperatures. Turk. J. Biol. 2005, 29, 35–39. [Google Scholar]
- Ali, S.; Pervez, R.; Hussain, M.A.; Ahmad, R. Effect of temperature on survival of Steinernema seemae, S. masoodi and S. carpocapsae (Rhabditida: Steinernematidae) and their subsequent infectivity to prepupa of Helicoverpa armigera (Hübner). Arch. Phytopathol. Pl. Prot. 2007, 40, 183–187. [Google Scholar] [CrossRef]
- Bilgrami, A.; Gaugler, R. Effects of various stress factors on heat tolerance by Heterorhabditis bacteriophora and Steinernema carpocapsae. Nematology 2007, 9, 161–167. [Google Scholar] [CrossRef]
- Pervez, R.; Ali, S.; Ahmad, R. Effect of temperatures on the emergence of entomopathogenic nematodes. Int. J. Nematol. 2008, 18, 25–28. [Google Scholar]
- Susurluk, I.A. Influence of temperature on the vertical movement of the entomopathogenic nematodes Steinernema feltiae (TUR-S3) and Heterorhabditis bacteriophora (TUR-H2), and infectivity of the moving nematodes. Nematology 2008, 10, 137–141. [Google Scholar] [CrossRef]
- Rashid, P.; Eapen, S.J.; Devasahayam, S.; Jacob, T. Effect of temperature on the infectivity of entomopathogenic nematodes against shoot borer (Conogethes punctiferalis Guen.) infesting ginger (Zingiber officinale Rosc.). J. Biol. Control 2015, 29, 187–193. [Google Scholar]
- Hu, X.P.; Dhang, P. Urban Pest Management: An Environmental Perspective; CABI: Wallingford, UK, 2011. [Google Scholar]
- Quarcoo, F.Y.; Appel, A.G.; Ping Hu, X. Effects of indoxacarb concentration and exposure time on onset of abnormal behaviors, morbundity, and death in eastern subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2010, 103, 762–769. [Google Scholar] [CrossRef]
- Woodring, J.L.; Kaya, H.K. Steinernematid and Heterorhabditid Nematodes: A Handbook of Biology and Techniques; Southern cooperative series bulletin (USA); Arkansas Agricultural Experiment Station: Fayetteville, AR, USA, 1988. [Google Scholar]
- Zadji, L.; Baimey, H.; Afouda, L.; Moens, M.; Decraemer, W. Characterization of biocontrol traits of heterorhabditid entomopathogenic nematode isolates from South Benin targeting the termite pest Macrotermes bellicosus. BioControl 2014, 59, 333–344. [Google Scholar] [CrossRef]
- Razia, M.; Baïmey, H.; Zadji, L.; Afouda, L.; Fanou, A.; Kotchofa, R.; Decraemer, W. Searching for better methodologies for successful control of termites using entomopathogenic nematodes. In Nematology-Concepts, Diagnosis and Control; Intech: London, UK, 2017; p. 53. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: Hoboken, NJ, USA, 1984. [Google Scholar]
- Kuehl, R.O. Designs of Experiments: Statistical Principles of Research Design and Analysis; Duxbury Press: London, UK, 2000. [Google Scholar]
- Kaya, H.K.; Gaugler, R. Entomopathogenic nematodes. Ann Rev. Entomol. 1993, 38, 181–206. [Google Scholar] [CrossRef]
- Bhairavi, K.S.; Bhattacharyya, B.; Devi, G.; Bhagawati, S.; Das, P.P.G.; Devi, E.B.; Manpoong, N.S. Evaluation of two native entomopathogenic nematodes against Odontotermes obesus (Rambur)(Isoptera: Termitidae) and Agrotis ipsilon (Hufnagel) (Lepidoptera: Noctuidae). Egypt J. Biol. Pest Control 2021, 31, 111. [Google Scholar] [CrossRef]
- Danthanarayana, W.; Vitarana, S.I. Control of the live-wood tea termite Glyptotermes dilatatus using Heterorhabditis sp. (Nemat.). Agric. Ecosyst. Environ. 1987, 19, 333–342. [Google Scholar] [CrossRef]
- Glazer, I.; Navon, A. Activity and persistence of entomoparasitic nematodes tested against Heliothis armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 1990, 83, 1795–1800. [Google Scholar] [CrossRef]
- Peters, A.; Ehlers, R.-U. Susceptibility of leatherjackets (Tipula paludosa and Tipula oleracea; Tipulidae; Nematocera) to the entomopathogenic nematode Steinernema feltiae. J. Invertebr. Pathol. 1994, 63, 163–171. [Google Scholar] [CrossRef]
- Radhakrishnan, S.; Shanmugam, S. Bioefficacy of entomopathogenic nematodes against Spodoptera litura (Lepidoptera: Noctuidae) in Bhendi. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2314–2319. [Google Scholar] [CrossRef] [Green Version]
- Burman, M. Neoaplectana carpocapsae: Toxin production by axenic insect parasitic nematodes. Nematologica 1982, 28, 62–70. [Google Scholar] [CrossRef]
- Javed, S.; Ali, A.; Khanum, T.A. Biocontrol potential of the entomopathogenic nematodes (Rhabditida: Steinernematidae and Heterorhabditidae) against the termite species, Microtermes obesi (Holmgren) (Blattodea: Termitidae). Egypt J. Biol. Pest Control. 2021, 31, 99. [Google Scholar] [CrossRef]
- Maketon, M.; Hominchan, A.; Hotaka, D. Control of American cockroach (Periplaneta americana) and German cockroach (Blattella germanica) by entomopathogenic nematodes. Rev. Colomb. Entomol. 2010, 36, 249–253. [Google Scholar] [CrossRef]
- Kamali, S.; Karimi, J.; Hosseini, M.; Campos-Herrera, R.; Duncan, L.W. Biocontrol potential of the entomopathogenic nematodes Heterorhabditis bacteriophora and Steinernema carpocapsae on cucurbit fly, Dacus ciliatus (Diptera: Tephritidae). Biocontrol Sci. Technol. 2013, 23, 1307–1323. [Google Scholar] [CrossRef]
- Mahar, A.N.; Munir, M.; Elawad, S.; Gowen, S.R.; Hague, N.G.M. Pathogenicity of bacterium, Xenorhabdus nematophila isolated from entomopathogenic nematode (Steinernema carpocapsae) and its secretion against Galleria mellonella larvae. J. Zhejiang Univ. Sci. 2005, 6, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, E.; Gaugler, R.; Harrison, R. Entomopathogenic nematode host finding: Response to host contact cues by cruise and ambush foragers. Parasitology 1992, 105, 309–315. [Google Scholar] [CrossRef]
- Gaugler, R.; Kaya, H.K. Entomopathogenic Nematodes in Biological Control; CRC Press: Boca Raton, FL, USA, 1990; Volume 227. [Google Scholar]
- Glazer, I.; Alekseev, E.; Samish, M. Factors affecting the virulence of entomopathogenic nematodes to engorged female Boophilus annulatus ticks. J. Parasitol. 2001, 87, 808–812. [Google Scholar] [CrossRef]
- Usman, A.; Ali, M.I.; Shah, M.; e Amin, F.; Sarwar, J. 8. Comparative efficacy of indigenous plant extracts and a synthetic insecticide for the management of tomato fruit worm (Helicoverpa armigera Hub.) and their effect on natural enemies in tomato crop. Pure Appl. Biol. (PAB) 2018, 7, 1014–1020. [Google Scholar] [CrossRef]
- Gaugler, R.; Lewis, E.; Stuart, R.J. Ecology in the service of biological control: The case of entomopathogenic nematodes. Oecologia 1997, 109, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Hanif, C.; Shahid, M.; Mansha, M.Z.; Aatif, H.M.; Ijaz, M.; Ikram, K.; Khan, A.A.; Ismail, M.; Ashir, M. Aptness of entomogenus fungi with diatomaceous earth against various stored grain insect pests. Egypt. J. Biol. Pest Control 2022, 32, 14. [Google Scholar] [CrossRef]
- Langford, E.A.; Nielsen, U.N.; Johnson, S.N.; Riegler, M. Susceptibility of Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae), to entomopathogenic nematodes. Biol. Control 2014, 69, 34–39. [Google Scholar] [CrossRef]
- Trdan, S.; Vidrih, M.; Andjus, L.; Laznik, Ž. Activity of four entomopathogenic nematode species against different developmental stages of Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera, Chrysomelidae). Helminthologia 2009, 46, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Rohde, C.; Moino Jr, A.; da Silva, M.A.; Carvalho, F.D.; Ferreira, C.S. Influence of soil temperature and moisture on the infectivity of entomopathogenic nematodes (Rhabditida: Heterorhabditidae, Steinernematidae) against larvae of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Neotrop. Entomol. 2010, 39, 608–611. [Google Scholar] [CrossRef] [Green Version]
- Foelkel, E.; Monteiro, L.B.; Voss, M. Virulence of nematodes against larvae of the south-American fruit fly in laboratory using soil from Porto Amazonas, Paraná, Brazil, as substrate. Ciênc Rural 2016, 46, 405–410. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Mortality (%) (Mean ± SE) |
|---|---|
| Bioassays (B) | |
| Sawdust | 47.02 ± 2.79 a |
| Filter paper | 39.33 ± 2.34 b |
| Nematode species (S) | |
| Steinernema carpocapsae | 48.28 ± 3.12 a |
| Heterorhabditis bacteriophora Heterorhabditis indica | 43.17 ± 2.63 b 38.08 ± 2.19 c |
| Nematode concentrations (IJs/termite) (C) | |
| 200 | 23.06 ± 2.06 e |
| 400 | 32.62 ± 2.25 d |
| 600 | 41.89 ± 1.95 c |
| 800 | 53.05 ± 3.54 b |
| 1000 | 65.27 ± 2.89 a |
| F-value (B) | 47.20 ** |
| F-value (S) | 27.65 ** |
| F-value (C) | 175.91 ** |
| F-value (B × S) | 0.54 * |
| F-value (B × C) | 0.12 * |
| F-value (S × C) | 0.80 NS |
| F-value (B × S × C) | 0.02 NS |
| Factors | Mortality (%) (Mean ± SE) |
|---|---|
| Bioassays (B) | |
| Sawdust | 61.37 ± 3.12 a |
| Filter paper | 53.86 ± 2.51 b |
| Nematode species (S) | |
| Steinernema carpocapsae | 66.17 ± 2.73 a |
| Heterorhabditis bacteriophora Heterorhabditis indica | 56.34 ± 1.74 b 50.34 ± 1.34 c |
| Nematode concentrations (IJs/termite) (C) | |
| 200 | 35.45 ± 1.82 e |
| 400 | 45.84 ± 3.48 d |
| 600 | 56.92 ± 2.52 c |
| 800 | 67.08 ± 3.30 b |
| 1000 | 82.79 ± 2.79 a |
| F-value (B) | 60.43 ** |
| F-value (S) | 91.10 ** |
| F-value (C) | 289.68 ** |
| F-value (B × S) | 0.19 * |
| F-value (B × C) | 0.02 * |
| F-value (S × C) | 3.27 * |
| F-value (B × S × C) | 0.31 NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, A.; Chi, D.-F.; Abbasi, A.; Arshad, M. Biocontrol Potential of Entomopathogenic Nematodes against Odontotermes obesus (Blattodea: Termitidae) under Laboratory and Field Conditions. Forests 2023, 14, 580. https://doi.org/10.3390/f14030580
Aslam A, Chi D-F, Abbasi A, Arshad M. Biocontrol Potential of Entomopathogenic Nematodes against Odontotermes obesus (Blattodea: Termitidae) under Laboratory and Field Conditions. Forests. 2023; 14(3):580. https://doi.org/10.3390/f14030580
Chicago/Turabian StyleAslam, Asad, De-Fu Chi, Asim Abbasi, and Muhammad Arshad. 2023. "Biocontrol Potential of Entomopathogenic Nematodes against Odontotermes obesus (Blattodea: Termitidae) under Laboratory and Field Conditions" Forests 14, no. 3: 580. https://doi.org/10.3390/f14030580