
Blackwellomyces kaihuaensis and Metarhizium putuoense (Hypocreales), Two New Entomogenous Fungi from Subtropical Forests in Zhejiang Province, Eastern China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Collection
2.2. Morphological Observation
2.3. DNA Isolation, PCR Amplification and Sequencing
2.4. Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher No. | Location | Host | nrSSU | nrLSU | tef1 | rpb1 | rpb2 | References |
|---|---|---|---|---|---|---|---|---|---|
| Ascopolyporus polychrous | P.C. 546 | Bolivia | Coccoideae | – | DQ118737 | DQ118745 | DQ127236 | – | [27] |
| A. villosus | ARSEF 6355 | Panama | Coccoideae | – | AY886544 | DQ118750 | DQ127241 | – | [27] |
| Blackwellomyces aurantiacus | BCC 85060 T | Thailand | Lepidoptera (larva) | – | MT003028 | MK411598 | MK411600 | MT017819 | [7] |
| B. aurantiacus | BCC 85061 | Thailand | Lepidoptera (larva) | – | MT003029 | MK411599 | MK411601 | MT017820 | [7] |
| B. calendulinus | BCC 68500 | Thailand | Coleoptera (larva) | – | MT003030 | MT017842 | MT017802 | MT017821 | [7] |
| B. calendulinus | BCC 68502 T | Thailand | Coleoptera (larva) | – | MT003031 | MT017843 | MT017803 | MT017822 | [7] |
| B. cardinalis | OSC 93610 | USA | Lepidoptera (larva) | AY184974 | AY184963 | EF469059 | EF469088 | EF469106 | [4] |
| B. cardinalis | CBS 113411 T | USA | Lepidoptera (larva) | NG_013131 | MH874496 | – | – | – | [28] |
| B. cardinalis | OSC 93609 | USA | Lepidoptera (larva) | AY184973 | AY184962 | DQ522325 | DQ522370 | DQ522422 | [4] |
| B. cardinalis | OSC 93619 | Japan | Lepidoptera (larva) | AY184975 | AY184964 | – | – | – | [4] |
| B. cardinalis | OSC 93620 | Japan | Lepidoptera (larva) | AY184976 | AY184965 | – | – | – | [4] |
| B. kaihuaensis | HMAS 285455 T | China | Lepidoptera (larva) | OQ981975 | OQ981968 | OQ980401 | OQ980409 | OQ980408 | This study |
| B. kaihuaensis | HMAS 285456 | China | Lepidoptera (larva) | OQ981976 | OQ981969 | OQ980402 | OQ980410 | – | This study |
| B. lateris | MFLU 18-0663 T | Thailand | Lepidoptera (larva) | NG_067678 | NG_067857 | MK069471 | MK084615 | MK079354 | [6] |
| B. minutus | BCC 88269 T | Thailand | Coleoptera (larva) | – | MT003032 | MT017844 | MT017804 | MT017823 | [7] |
| B. pseudomilitaris | TBRC 3662 | Thailand | Lepidoptera (larva) | – | MT003036 | MT017848 | MT017808 | – | [7] |
| B. pseudomilitaris | BCC 73634 | Thailand | Lepidoptera (larva) | – | MT017849 | MT017809 | MT017827 | [7] | |
| B. pseudomilitaris | BCC 1919 T | – | Lepidoptera (larva) | MF416588 | MF416534 | MF416478 | – | MF416440 | [3] |
| B. pseudomilitaris | BCC 2091 | – | Lepidoptera (larva) | MF416589 | MF416535 | MF416479 | – | MF416441 | [3] |
| B. pseudomilitaris | NBRC 101409 | Thailand | Lepidoptera (larva) | JN941748 | JN941393 | – | JN992482 | – | [29] |
| B. pseudomilitaris | NBRC 101410 | Thailand | Lepidoptera (larva) | JN941747 | JN941394 | – | JN992481 | – | [29] |
| B. pseudomilitaris | NBRC 101411 | Thailand | Lepidoptera (larva) | JN941746 | JN941395 | – | JN992480 | – | [29] |
| B. pseudomilitaris | NBRC 101413 | Thailand | Lepidoptera (larva) | JN941745 | JN941396 | – | JN992479 | – | [29] |
| B. roseostromatus | BCC 91358 T | Thailand | Lepidoptera (larva) | – | MT003033 | MT017845 | MT017805 | MT017824 | [7] |
| B. roseostromatus | BCC 91359 | Thailand | Lepidoptera (larva) | – | MT003034 | MT017846 | MT017806 | MT017825 | [7] |
| B. roseostromatus | BCC 91360 | Thailand | Lepidoptera (larva) | – | MT003035 | MT017847 | MT017807 | MT017826 | [7] |
| Lecanicillium psalliotae | CBS 101270 | UK | Soil | EF469128 | EF469081 | EF469066 | EF469095 | EF469113 | [30] |
| Gamszarea wallacei | CBS 101237 | Indonesia | Lepidoptera (pupa) | NG_062646 | NG_042398 | EF469073 | EF469102 | EF469119 | [30] |
| Aschersonia badia | BCC 8105 | Thailand | Hemiptera: scale insect | DQ522537 | DQ518752 | DQ522317 | DQ522363 | DQ522411 | [31] |
| A. placenta | BCC 7869 | Thailand | Hemiptera: scale insect | EF469121 | EF469074 | EF469056 | EF469085 | EF469104 | [30] |
| Balansia henningsiana | A.E.G. 96-27a | USA | Poaceae: Panicum sp. | AY545723 | AY545727 | AY489610 | AY489643 | DQ522413 | [20] |
| B. pilulaeformis | A.E.G. 94-2 | – | Poaceae | AF543764 | AF543788 | DQ522319 | DQ522365 | DQ522414 | [30] |
| Claviceps fusiformis | ATCC 26019 | – | Poaceae | DQ522539 | U17402 | DQ522320 | DQ522366 | – | [30] |
| C. paspali | ATCC 13892 | – | Poaceae | U32401 | U47826 | DQ522321 | DQ522367 | DQ522416 | [30] |
| C. purpurea | S.A. cp11 | – | Poaceae | EF469122 | EF469075 | EF469058 | EF469087 | EF469105 | [30] |
| Cordyceps cylindrica | CBS 744.73 | Japan | Arachnida: spider | EF468987 | EF468841 | EF468786 | EF468892 | – | [30] |
| Hypocrella schizostachyi | BCC 14123 | Thailand | Hemiptera | DQ522557 | DQ518771 | DQ522346 | DQ522392 | DQ522447 | [30] |
| H. nectrioides | GJS 89-104 | – | Hemiptera: scale insect | – | DQ518772 | DQ522347 | DQ522393 | DQ522448 | [31] |
| Keithomyces acicularis | JCM 33284 | Japan | Soil | LC435738 | LC435741 | LC462188 | – | – | [32] |
| K. acicularis | JCM 33285 | Japan | Soil | LC435739 | LC435742 | LC462189 | – | – | [32] |
| K. carneus | CBS 239.32 | France | Sand dune | EF468988 | EF468843 | EF468789 | EF468894 | EF468938 | [31] |
| K. carneus | CBS 399.59 | USA | Soil | EF468989 | EF468842 | EF468788 | EF468895 | EF468939 | [31] |
| K. neogunnii | BUM 415 | China | Lepidoptera (larva) | MH143845 | MH143828 | MH143861 | MH143876 | MH143891 | [33] |
| K. neogunnii | GZUH SB13050305 | China | Lepidoptera (larva) | KU729724 | – | KU729729 | KU729734 | – | [34] |
| Keithomyces sp. | CBS 126563 | Tanzania | Soil | MT078871 | MT078856 | MT078848 | MT078864 | MT078921 | [23] |
| Keithomyces sp. | CBS 127407 | USA | Soil | MT078873 | MT078858 | MT078850 | MT078866 | MT078923 | [23] |
| Marquandomyces marquandii | CBS 127132 | USA | Soil | MT078872 | MT078857 | MT078849 | MT078865 | MT078922 | [23] |
| M. marquandii | CBS 129413 | USA | Soil | MT078874 | MT078859 | MT078851 | MT078867 | – | [23] |
| M. marquandii | CBS 182.27 | USA | Soil | EF468990 | EF468845 | EF468793 | EF468899 | EF468942 | [30] |
| Metacordyceps chlamydosporia | CBS 101244 | – | Diplopoda: egg of slug | DQ522544 | DQ518758 | DQ522327 | DQ522372 | DQ522424 | [31] |
| M. chlamydosporia | CBS 504.66 | Canada | Soil | AF339593 | AF339544 | EF469069 | EF469098 | EF469120 | [30] |
| Metapochonia bulbillosa | CBS 145.70 | Denmark | Picea abies | AF339591 | AF339542 | EF468796 | EF468902 | EF468943 | [30] |
| M. goniodes | CBS 891.72 | Germany | Fungi | AF339599 | AF339550 | DQ522354 | DQ522401 | DQ522458 | [30] |
| M. rubescens | CBS 464.88 | Scotland | Nematode eggs | AF339615 | AF339566 | EF468797 | EF468903 | EF468944 | [30] |
| Metarhizium acridum | ARSEF 7486 | Niger | Orthoptera | – | – | EU248845 | EU248897 | EU248925 | [10] |
| M. album | ARSEF 2082 | Indonesia | Hemiptera | DQ522560 | DQ518775 | DQ522352 | DQ522398 | DQ522452 | [30] |
| M. alvesii | CG1123 | Brazil | Soil | – | – | KY007614 | KY007612 | KY007613 | [35] |
| M. anisopliae | ARSEF 7487 | Ethiopia | Orthoptera | – | – | DQ463996 | DQ468355 | DQ468370 | [10] |
| M. anisopliae | CBS 130.71 | Ukraine | Avena sativa | MT078868 | MT078853 | MT078845 | MT078861 | MT078918 | [23] |
| M. argentinense | CEP424 | Argentina | Blaberidae: Epilamprinae | – | – | MF966624 | MF966625 | MF966626 | [36] |
| M. atrovirens | TNM F10184 | Japan | Coleoptera | JF415950 | JF415966 | – | JN049884 | – | [11] |
| M. baoshanense | BUM 63.4 | China | Soil | KY264178 | KY264175 | KY264170 | KY264181 | KY264184 | [37] |
| M. baoshanense | CCTCCM 2016589 | China | Soil | KY264177 | KY264174 | KY264169 | KY264180 | KY264183 | [37] |
| M. bibionidarum | CBS 648.67 | France | Coleoptera | – | – | LC126075 | LC125907 | LC125923 | [38] |
| M. bibionidarum | NBRC 112661 | Japan | Diptera | – | – | LC126076 | LC125908 | LC125924 | [38] |
| M. biotecense | BCC 51812 T | Thailand | Hemiptera: Delphacidae | MN781937 | MN781838 | MN781693 | MN781745 | MN781792 | [23] |
| M. biotecense | BCC 51813 | Thailand | Hemiptera: Delphacidae | MN781938 | MN781839 | MN781694 | MN781746 | MN781793 | [23] |
| M. blattodeae | MY00896 | Thailand | Blattodea | HQ165657 | HQ165719 | HQ165678 | HQ165739 | HQ165638 | [12] |
| M. brachyspermum | CM1 | Japan | Coleoptera | – | LC469749 | LC469751 | – | – | [39] |
| M. brasiliense | ARSEF 2948 | Brazil | Hemiptera | – | – | KJ398809 | KJ398620 | – | [40] |
| M. brittlebankisoides | Hn1 | China | Coleoptera | – | – | AB778556 | AB778555 | AB778554 | [41] |
| M. brunneum | ARSEF 2107 | USA | Coleoptera | – | – | EU248855 | EU248907 | EU248935 | [10] |
| M. candelabrum | BCC 29224 T | Thailand | Hemiptera: leafhopper | MN781952 | MN781853 | MN781708 | MN781755 | MN781804 | [23] |
| M. cercopidarum | BCC 31660 T | Thailand | Hemiptera: leafhopper | MN781953 | MN781854 | MN781709 | MN781756 | MN781805 | [23] |
| M. chaiyaphumense | BCC 78198 | Thailand | Hemiptera: Cicadidae | KX369596 | KX369593 | KX369592 | KX369594 | KX369595 | [12] |
| M. chaiyaphumense | BCC 28241 | Thailand | Hemiptera: Cicadidae | MN781932 | MN781831 | MN781684 | MN781740 | MN781784 | [23] |
| M. cicadae | BCC 48696 | Thailand | Hemiptera: Cicadidae | MN781948 | MN781848 | MN781703 | – | MN781800 | [23] |
| M. cicadae | BCC 48881 T | Thailand | Hemiptera: Cicadidae | MN781949 | MN781849 | MN781704 | MN781752 | – | [23] |
| M. clavatum | BCC 84543 T | Thailand | Coleoptera (larva) | – | MN781834 | MN781689 | MN781741 | MN781789 | [23] |
| M. clavatum | BCC 84558 | Thailand | Coleoptera (larva) | – | MN781835 | MN781690 | MN781742 | – | [23] |
| M. culicidarum | BCC 7625 | Thailand | Diptera: Culicidae | – | MN781850 | MN781705 | – | MN781801 | [23] |
| M. culicidarum | BCC 2673 | Thailand | Diptera: Culicidae | MN781950 | MN781851 | MN781706 | MN781753 | MN781802 | [23] |
| M. culicidarum | BCC 7600 T | Thailand | Diptera: Culicidae | MN781951 | MN781852 | MN781707 | MN781754 | MN781803 | [23] |
| M. cylindrosporum | RCEF 3632 | China | Hemiptera: Cicadidae | JF415964 | JF415987 | JF416022 | – | – | [11] |
| M. cylindrosporum | TNS 16371 | Japan | Hemiptera: Cicadidae | JF415963 | JF415986 | JF416027 | JN049902 | – | [11] |
| M. dendrolimatilis | GZAC-IFR1006 | China | Lepidoptera | – | – | KT166031 | KT961694 | KT166032 | [42] |
| M. eburneum | BCC 79267 | Thailand | Lepidoptera (pupa) | – | MN781826 | – | MN781735 | – | [23] |
| M. eburneum | BCC 79252 T | Thailand | Lepidoptera (pupa) | – | MN781829 | MN781682 | MN781736 | – | [23] |
| M. ellipsoideum | BCC 49285 T | Thailand | Hemiptera (adult) | MN781957 | MN781858 | MN781713 | MN781759 | MN781808 | [23] |
| M. ellipsoideum | BCC 12847 | Thailand | Hemiptera (adult) | MN781959 | MN781860 | MN781715 | MN781761 | MN781810 | [23] |
| M. ellipsoideum | BCC 53509 | Thailand | Hemiptera (adult) | MN781958 | MN781859 | MN781714 | MN781760 | MN781809 | [23] |
| M. flavoviride | CBS 218.56 | Czech Republic | Coleoptera | – | – | KJ398787 | KJ398598 | – | [40] |
| M. flavoviride | CBS 125.65 | USA | Soil | MT078869 | MT078854 | MT078846 | MT078862 | MT078919 | [23] |
| M. flavum | BCC 90870 T | Thailand | Coleoptera (larva) | MN781965 | MN781874 | MN781731 | MN781776 | MN781822 | [23] |
| M. flavum | BCC 90874 | Thailand | Coleoptera (larva) | MN781966 | MN781875 | MN781732 | MN781777 | MN781823 | [23] |
| M. frigidum | ARSEF 4124 | Australia | Coleoptera | – | – | DQ464002 | DQ468361 | DQ468376 | [43] |
| M. fusoideum | BCC 41242 | Thailand | Psocoptera | MN781942 | MN781825 | MN781679 | – | MN781780 | [23] |
| M. fusoideum | BCC 53130 | Thailand | Psocoptera | MN781943 | MN781843 | MN781698 | – | MN781795 | [23] |
| M. fusoideum | BCC 28246 T | Thailand | Lepidoptera | MN781944 | MN781844 | MN781699 | MN781749 | MN781796 | [23] |
| M. gaoligongense | BUM 3.5 | China | Soil | KY087810 | KY087814 | KY087818 | KY087822 | – | [44] |
| M. gaoligongense | CCTCCM 2016588 | China | Soil | KY087812 | KY087816 | KY087820 | KY087824 | KY087826 | [44] |
| M. globosum | ARSEF 2596 | India | Lepidoptera | – | – | EU248846 | EU248898 | EU248926 | [11] |
| M. granulomatis | UAMH 11028 T | Denmark | Chamaeleo calyptratus | HM635076 | HM195304 | KJ398781 | – | – | [45] |
| M. granulomatis | UAMH 11176 | Denmark | Chamaeleo calyptratus | – | HM635078 | KJ398782 | KJ398593 | – | [45] |
| M. gryllidicola | BCC 37918 | Thailand | Orthoptera: Gryllidae | MN781935 | MN781836 | MN781691 | MN781743 | MN781790 | [46] |
| M. gryllidicola | BCC 82988 | Thailand | Orthoptera: Gryllidae | MK632117 | MK632091 | MK632062 | MK632166 | MK632143 | [46] |
| M. guizhouense | ARSEF 6238 | China | Lepidoptera | – | – | EU248857 | EU248909 | EU248937 | [10] |
| M. guizhouense | CBS 258.90 | China | Lepidoptera | – | – | EU248862 | EU248914 | EU248942 | [10] |
| M. huainamdangense | BCC 32190 | Thailand | Hemiptera: leafhopper | MN781954 | MN781855 | MN781710 | MN781757 | – | [23] |
| M. huainamdangense | BCC 44270 T | Thailand | Hemiptera: leafhopper | MN781956 | MN781857 | MN781712 | – | MN781807 | [23] |
| M. huainamdangense | BCC 7672 | Thailand | Hemiptera: leafhopper | MN781955 | MN781856 | MN781711 | MN781758 | MN781806 | [23] |
| M. humberi | IP46 | Brazil | Soil | – | – | MH837574 | MH837556 | MH837565 | [47] |
| M. indigoticum | TNS F18553 | Japan | Lepidoptera | JF415953 | JF415968 | JF416010 | JN049886 | JF415992 | [11] |
| M. indigoticum | TNS F18554 | Japan | Lepidoptera | JF415952 | JF415969 | JF416011 | JN049887 | JF415993 | [11] |
| M. kalasinense | BCC 53582 | Thailand | Coleoptera (larva) | KC011175 | KC011183 | KC011189 | – | – | [12] |
| M. koreanum | ARSEF 2039 | Republic of Korea | Hemiptera: Delphacidae | – | – | KJ398806 | KJ398616 | – | [40] |
| M. koreanum | BCC 27998 | Thailand | Hemiptera: Fulgoromorpha | MN781945 | MN781845 | MN781700 | – | MN781797 | [23] |
| M. koreanum | BCC 30455 | Thailand | Hemiptera: Fulgoromorpha | MN781946 | MN781846 | MN781701 | MN781750 | MN781798 | [23] |
| M. lepidiotae | ARSEF 7488 | Australia | Coleoptera | – | – | EU248865 | EU248917 | EU248945 | [10] |
| M. majus | ARSEF 1015 | Japan | Lepidoptera | – | – | EU248866 | EU248918 | EU248946 | [10] |
| M. majus | ARSEF 1914 | Philippines | Coleoptera | – | – | EU248868 | EU248920 | EU248948 | [10] |
| M. megapomponiae | BCC 25100 T | Thailand | Hemiptera: Megopomponia | MN781947 | MN781847 | MN781702 | MN781751 | MN781799 | [23] |
| M. minus | ARSEF 2037 | Philippines | Hemiptera | AF339580 | AF339531 | DQ522353 | DQ522400 | DQ522454 | [30] |
| M. minus | ARSEF 1099 | Philippines | Hemiptera | – | – | KJ398799 | KJ398608 | KJ398706 | [40] |
| M. niveum | BCC 52400 T | Thailand | Hemiptera: Cicadidae | MN781933 | MN781832 | MN781685 | – | MN781785 | [23] |
| M. nornnoi | BCC 19364 | Thailand | Lepidoptera (larva) | MN781940 | MN781841 | MN781696 | MN781747 | – | [23] |
| M. nornnoi | BCC 25948 T | Thailand | Coleoptera (adult beetle) | MN781941 | MN781842 | MN781697 | MN781748 | – | [23] |
| M. novozealandicum | ARSEF 4661 | Australia | Soil | – | – | KJ398811 | KJ398622 | – | [40] |
| M. novozealandicum | ARSEF 4674 | Australia | Soil | – | – | KJ398812 | KJ398623 | – | [40] |
| M. ovoidosporum | BCC 29223 | Thailand | Hemiptera: Cercopidae | MN781960 | MN781861 | MN781716 | MN781762 | – | [23] |
| M. ovoidosporum | BCC 32600 T | Thailand | Hemiptera: Eurybrachidae | MN781961 | MN781862 | MN781717 | MN781763 | – | [23] |
| M. ovoidosporum | BCC 7634 | Thailand | Hemiptera (adult) | MN781962 | MN781863 | MN781718 | MN781764 | MN781811 | [23] |
| M. owariensis | NBRC 33258 | Japan | Hemiptera | HQ165669 | HQ165730 | HQ165689 | HQ165747 | – | [12] |
| M. pemphigi | ARSEF 7491 | UK | Hemiptera | – | – | KJ398819 | KJ398629 | DQ468379 | [40] |
| M. pemphigi | ARSEF 6569 | UK | Hemiptera | – | – | KJ398813 | KJ398624 | DQ468378 | [40] |
| M. phasmatodeae | BCC 49272 | Thailand | Orthoptera: Phasmatodea | MK632119 | MK632093 | MK632064 | – | MK632145 | [46] |
| M. phasmatodeae | BCC 2841 | Thailand | Orthoptera: Phasmatodea | MN781931 | MN781828 | MN781681 | MN781738 | MN781782 | [46] |
| M. phuwiangense | BCC 78206 | Thailand | Coleoptera (adult) | – | – | MN781719 | MN781765 | MN781812 | [23] |
| M. phuwiangense | BCC 85068 | Thailand | Coleoptera (adult) | – | MN781864 | MN781720 | MN781766 | MN781813 | [23] |
| M. phuwiangense | BCC 85069 T | Thailand | Coleoptera (adult) | – | MN781865 | MN781721 | MN781767 | MN781814 | [23] |
| M. pingshaense | CBS 257.90 | China | Coleoptera | – | – | EU248850 | EU248902 | EU248930 | [10] |
| M. prachinense | BCC 47950 | Thailand | Lepidoptera | KC011172 | KC011180 | KC011186 | KC011184 | – | [12] |
| M. prachinense | BCC 47979 | Thailand | Lepidoptera | KC011173 | KC011181 | KC011187 | KC011185 | – | [12] |
| M. pseudoatrovirens | TNS F16380 | Japan | Coleoptera | – | JF415977 | – | JN049893 | JF415997 | [11] |
| M. purpureogenum | MAFF 243305 | Japan | Soil | – | AB700552 | LC126078 | LC125913 | LC125920 | [38] |
| M. purpureogenum | MAFF 244762 | Japan | Soil | – | – | LC126079 | LC125911 | LC125922 | [38] |
| M. purpureonigrum | BCC 89247 T | Thailand | Coleoptera (larva) | – | – | MN781725 | MN781771 | MN781817 | [23] |
| M. purpureonigrum | BCC 89249 | Thailand | Coleoptera (larva) | MN781963 | MN781869 | MN781726 | MN781772 | MN781818 | [23] |
| M. purpureonigrum | BCC 89248 | Thailand | Coleoptera (larva) | MN781964 | MN781870 | MN781727 | – | MN781819 | [23] |
| M. purpureum | BCC 82173 | Thailand | Coleoptera (larva) | – | MN781866 | MN781722 | MN781768 | MN781815 | [23] |
| M. purpureum | BCC 82642 T | Thailand | Coleoptera (larva) | – | MN781867 | MN781723 | MN781769 | MN781816 | [23] |
| M. purpureum | BCC 83548 | Thailand | Coleoptera (larva) | – | MN781868 | MN781724 | MN781770 | – | [23] |
| M. putuoense | HMAS 285457 T | China | Coleoptera (larva) | OQ981977 | OQ981970 | OQ980403 | OQ980411 | – | This study |
| M. putuoense | HMAS 285458 | China | Coleoptera (larva) | OQ981978 | OQ981971 | OQ980404 | OQ980412 | – | This study |
| M. putuoense | HMAS 285459 | China | Coleoptera (larva) | OQ981979 | OQ981972 | OQ980405 | OQ980413 | – | This study |
| M. putuoense | HMAS 285460 | China | Coleoptera (larva) | OQ981980 | OQ981973 | OQ980406 | OQ980414 | – | This study |
| M. putuoense | LY 280 | China | Coleoptera (larva) | OQ981981 | OQ981974 | OQ980407 | – | – | This study |
| M. reniforme | IndGH96 | Philippines | – | HQ165670 | HQ165732 | – | – | HQ165649 | [12] |
| M. reniforme | ARSEF 429 | Philippines | Orthoptera: Tettigoniidae | HQ165671 | HQ165733 | HQ165690 | – | HQ165650 | [12] |
| M. rileyi | CBS 806.71 | USA | Trichoplusia ni | AY526491 | – | EF468787 | EF468893 | EF468937 | [30] |
| M. rileyi | NBRC 8560 | Japan | Lepidoptera | HQ165667 | HQ165729 | HQ165688 | – | – | [12] |
| M. robertsii | ARSEF 4739 | Australia | Soil | – | – | EU248848 | EU248900 | EU248928 | [10] |
| M. samlanense | BCC 17091 | Thailand | Hemiptera (adult) | HQ165665 | HQ165727 | HQ165686 | – | HQ165646 | [12] |
| M. samlanense | BCC 39752 | Thailand | Hemiptera (adult) | MN781939 | MN781840 | MN781695 | – | MN781794 | [12] |
| M. sulphureum | BCC 36585 | Thailand | Lepidoptera (larva) | – | – | MN781686 | – | MN781786 | [23] |
| M. sulphureum | BCC 39045 | Thailand | Lepidoptera (larva) | MK632120 | MK632095 | MK632066 | – | MK632147 | [23] |
| M. sulphureum | BCC 36592 T | Thailand | Lepidoptera (larva) | – | – | MN781687 | – | MN781787 | [23] |
| M. takense | BCC 30939 | Thailand | Hemiptera | HQ165659 | HQ165721 | – | HQ165741 | HQ165640 | [12] |
| M. takense | BCC 30934 | Thailand | Hemiptera | HQ165658 | HQ165720 | HQ165679 | HQ165740 | HQ165639 | [12] |
| M. viride | CBS 659.71 | – | Chameleo lateralis | HQ165673 | HQ165735 | HQ165692 | – | HQ165652 | [12] |
| M. viridulum | BCC 36261 | Thailand | Hemiptera: Cicadidae | MN781930 | MN781827 | MN781680 | MN781737 | MN781781 | [23] |
| M. viridulum | ARSEF 6927 | Taiwan | Hemiptera | – | – | KJ398815 | – | – | [40] |
| Myriogenospora atramentosa | A.E.G. 96-32 | – | Plant | AY489701 | AY489733 | AY489628 | AY489665 | DQ522455 | [30] |
| Nigelia aurantiaca | BCC 19950 | Thailand | Lepidoptera | GU979934 | GU979943 | GU979952 | GU979961 | GU979967 | [12] |
| N. aurantiaca | BCC 37621 | Thailand | Lepidoptera | GU979937 | GU979946 | GU979955 | GU979964 | GU979970 | [12] |
| N. martialis | EFCC 6863 | Republic of Korea | Lepidoptera | – | JF415975 | JF416016 | – | JF415995 | [12] |
| N. martialis | HMAS 197472 | China | Coleoptera: Cerambycidae | JF415955 | JF415973 | JF416015 | JN049892 | JF415994 | [12] |
| Papiliomyces liangshanensis | EFCC 1452 | Republic of Korea | Lepidoptera | EF468962 | EF468815 | EF468756 | – | – | [30] |
| P. liangshanensis | EFCC 1523 | Republic of Korea | Lepidoptera | EF468961 | EF468814 | EF468755 | – | EF468918 | [30] |
| P. shibinensis | GZUH SB13050311 | China | Lepidoptera | KR153588 | – | KR153589 | KR153590 | – | [48] |
| Petchia siamensis | BCC 68420 | Thailand | Ootheca of Mantidae | MK632113 | MK632087 | – | MK632163 | MK632140 | [46] |
| P. siamensis | BCC 73636 | Thailand | Ootheca of Mantidae | MK632115 | MK632089 | MK632060 | – | MK632138 | [46] |
| Purpureocillium lavendulum | FMR 10376 | Venezuela | Soil | – | FR775489 | FR775516 | FR775512 | – | [49] |
| P. lilacinus | CBS 284.36 | USA | Soil | AY526475 | FR775484 | EF468792 | EF468898 | EF468941 | [30] |
| P. takamizusanense | NHJ 3582 | Thailand | Hemiptera: Cicadidae | EU369097 | EU369034 | EU369015 | – | – | [50] |
| Purpureomyces khaoyaiense | BCC 1376 | Thailand | Lepidoptera (larva) | KX983468 | KX983462 | KX983457 | – | KX983465 | [12] |
| P. khaoyaiensis | BCC 14290 | Thailand | Lepidoptera (larva) | – | JF415970 | KJ398797 | JN049888 | – | [11] |
| P. maesotensis | BCC 88441 | Thailand | Lepidoptera (larva) | – | MN781877 | MN781734 | MN781779 | MN781824 | [23] |
| P. maesotensis | BCC 89300 T | Thailand | Lepidoptera (larva) | – | MN781876 | MN781733 | MN781778 | – | [23] |
| P. pyriformis | BCC 85074 T | Thailand | Lepidoptera (larva) | – | MN781873 | MN781730 | MN781775 | MN781821 | [23] |
| P. pyriformis | BCC 85348 | Thailand | Lepidoptera (larva) | – | MN781871 | MN781728 | MN781773 | MN781820 | [23] |
| P. pyriformis | BCC 85349 | Thailand | Lepidoptera (larva) | – | MN781872 | MN781729 | MN781774 | – | [23] |
| Regiocrella camerunensis | ARSEF 7682 | – | Hemiptera | – | DQ118735 | DQ118743 | DQ127234 | – | [51] |
| Rotiferophthora angustispora | CBS 101437 | – | Bdelloid rotifers | AF339584 | AF339535 | AF543776 | DQ522402 | DQ522460 | [30] |
| Shimizuomyces paradoxus | EFCC 6279 | Republic of Korea | Smilax sieboldii | EF469131 | EF469084 | EF469071 | EF469100 | EF469117 | [30] |
| S. paradoxus | EFCC 6564 | Republic of Korea | Smilax sieboldii | EF469130 | EF469083 | EF469072 | EF469101 | EF469118 | [30] |
| Sungia yongmunensis | EFCC 2131 | Republic of Korea | Lepidoptera | EF468977 | EF468833 | EF468770 | EF468876 | – | [30] |
| S. yongmunensis | EFCC 2135 | Republic of Korea | Lepidoptera | EF468979 | EF468834 | EF468769 | EF468877 | – | [30] |
| Torrubiella luteorostrata | NHJ 11343 | Thailand | Hemiptera: scale insect | EF468995 | EF468850 | EF468801 | EF468906 | – | [30] |
| T. luteorostrata | NHJ 12516 | Thailand | Hemiptera: scale insect | EF468994 | EF468849 | EF468800 | EF468905 | EF468946 | [30] |
| T. petchii | NHJ 6240 | Thailand | Hemiptera: scale insect | EU369103 | EU369038 | EU369022 | EU369060 | EU369082 | [50] |
| T. petchii | NHJ 6209 | Thailand | Hemiptera: scale insect | EU369104 | EU369039 | EU369023 | EU369061 | EU369081 | [50] |
| T. tenuis | NHJ 6293 | Thailand | Hemiptera: scale insect | EU369112 | EU369044 | EU369029 | EU369068 | EU369087 | [50] |
| T. tenuis | NHJ 345.01 | Thailand | Hemiptera: scale insect | EU369111 | EU369045 | EU369030 | – | EU369088 | [50] |
| Tyrannicordyceps fratricida | TNS 19011 | – | Fungi | JQ257022 | JQ257023 | JQ257028 | JQ257016 | JQ257021 | [52] |
| Yosiokobayasia kusanaginensis | TNS F18494 | Japan | Coleoptera | JF415954 | JF415972 | JF416014 | JN049890 | – | [11] |
3. Results
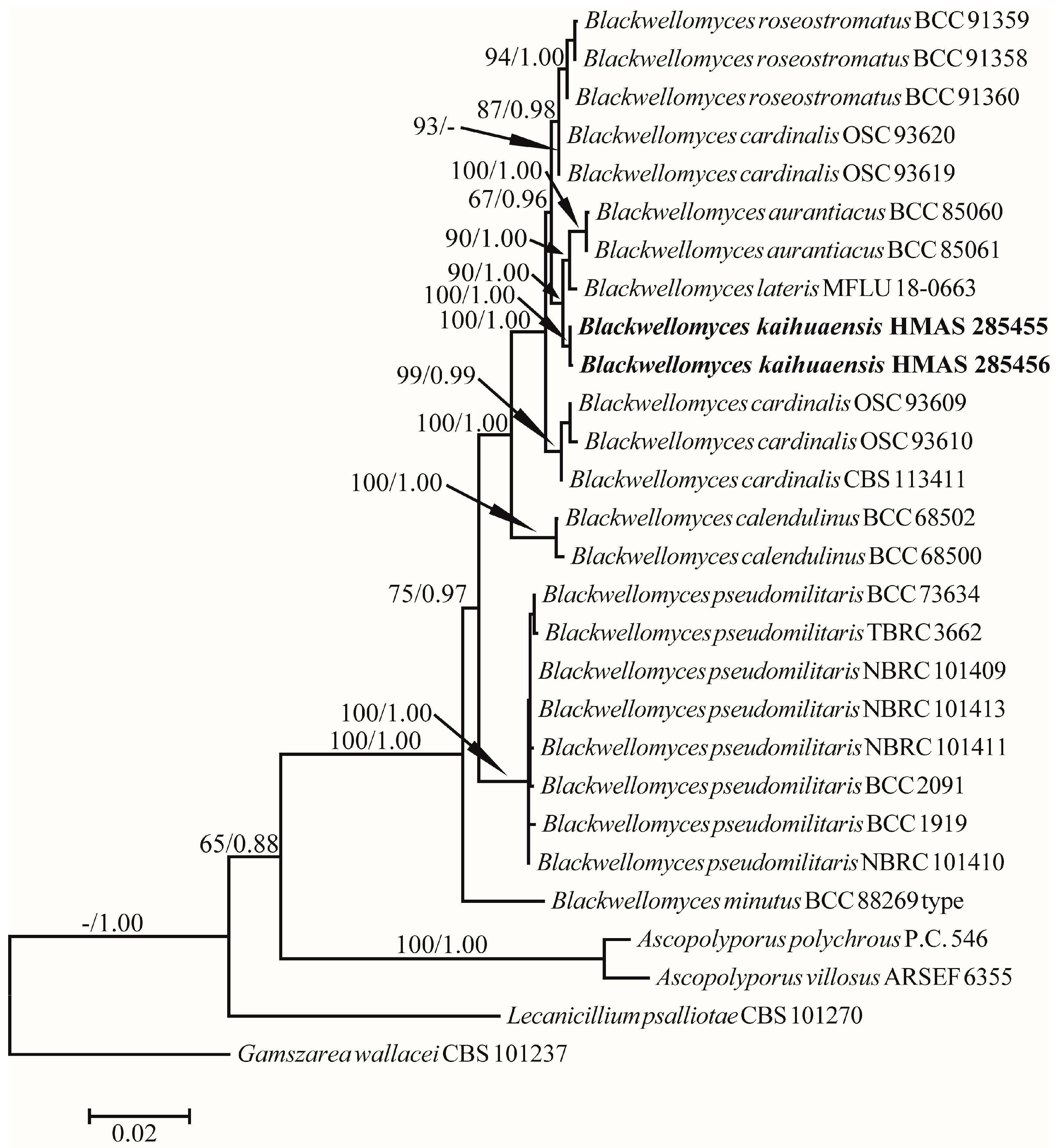
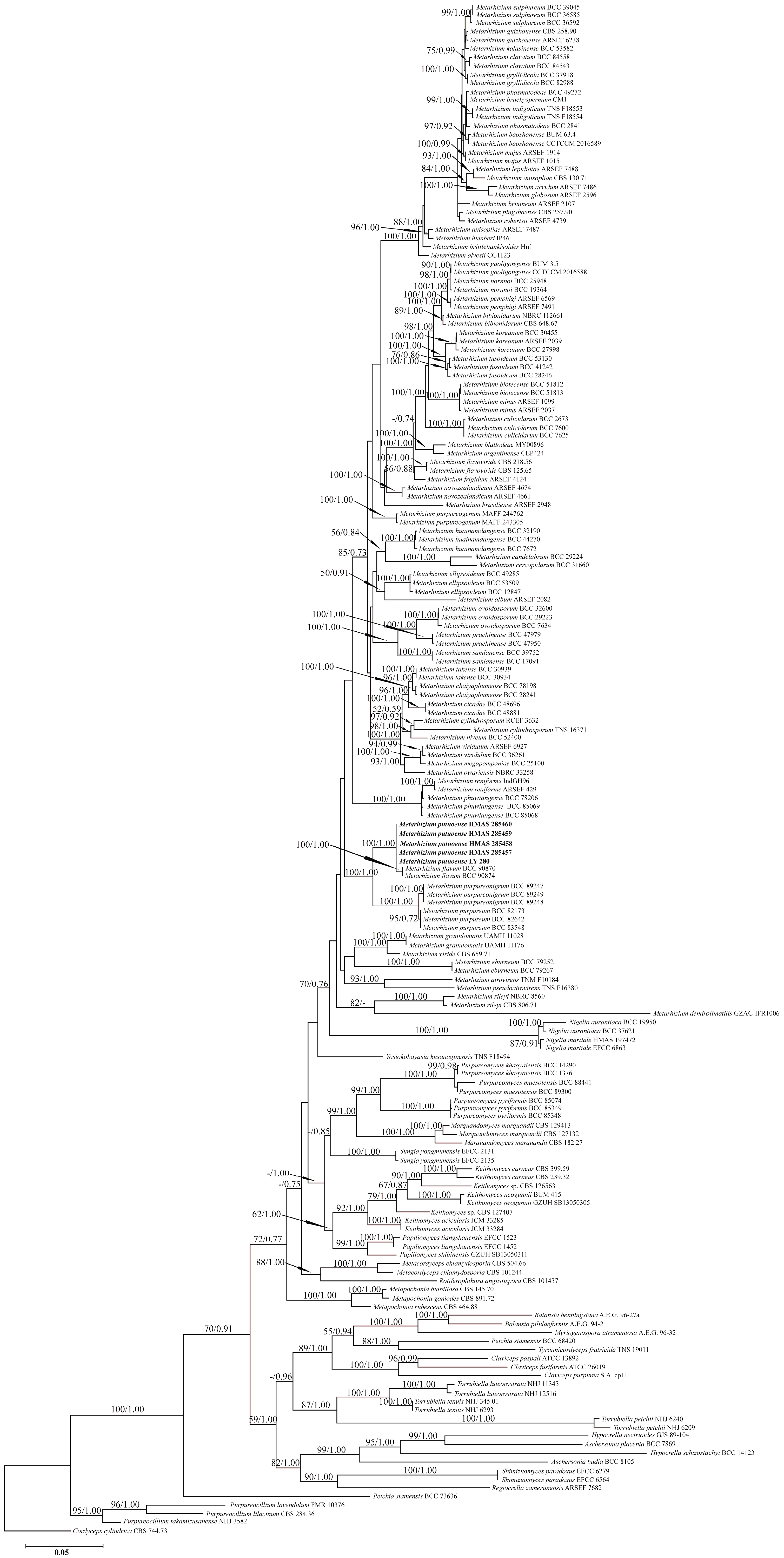
3.1. Phylogeny
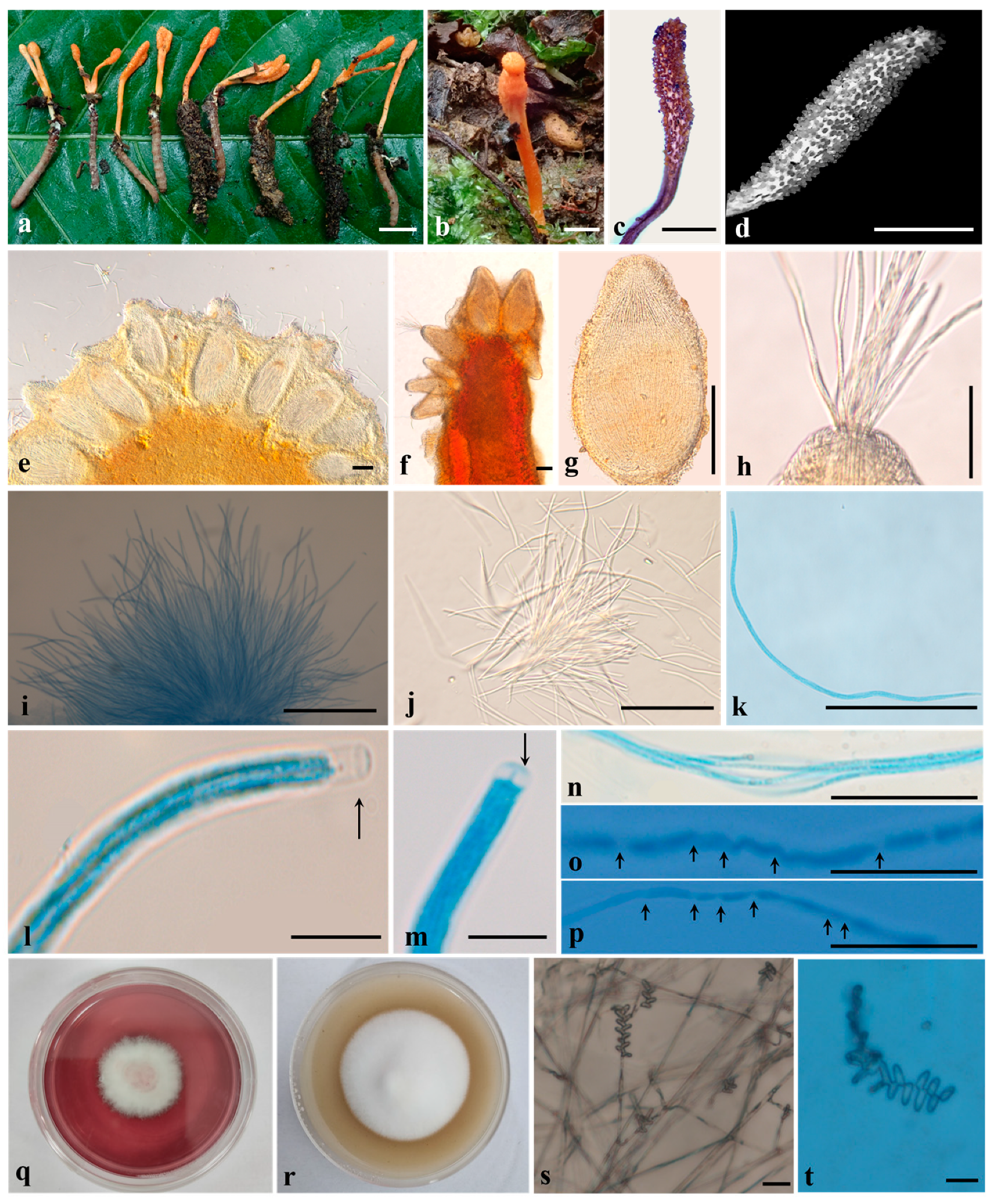
3.2. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic fungi: Unconventional applications. Rev. Environ. Sci. Bio 2020, 19, 23–42. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Eliopoulos, P.A. Endophytic entomopathogenic fungi: A valuable biological control tool against plant pests. Appl. Sci. 2020, 10, 360. [Google Scholar] [CrossRef]
- Kepler, R.M.; Luangsa-ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef]
- Sung, G.H.; Spatafora, J.W. Cordyceps cardinalis sp. nov., a new species of Cordyceps with an east Asian-eastern North American distribution. Mycologia 2004, 96, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Hywel-Jones, N.L. Cordyceps khaoyaiensis and C. pseudomilitaris, two new pathogens of lepidopteran larvae from Thailand. Mycol. Res. 1994, 98, 939–942. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Mongkolsamrit, S.; Noisripoom, W.; Tasanathai, K.; Khonsanit, A.; Thanakitpipattana, D.; Himaman, W.; Kobmoo, N.; Luangsa-ard, J.J. Molecular phylogeny and morphology reveal cryptic species in Blackwellomyces and Cordyceps (Cordycipitaceae) from Thailand. Mycol. Prog. 2020, 19, 957–983. [Google Scholar] [CrossRef]
- Zhang, S.P.; Li, Q.Y.; Zan, X.; Yan, Q.X.; Li, L.H. Studies of macrofungal resources in Micang Mountain Nature Reserve, Sichuan Province. Edible Fungi 2020, 42, 16–19. (In Chinese) [Google Scholar]
- Metschnikoff, E. Diseases of Wheat Chafers Zapiski imperatorskogo Obscestva Sel’skogo Hoziatistva Juznoi Roosi God Sorok Devjaytyi; Elsevier: Odessa, Ukraine, 1879; pp. 21–50. (In Russian) [Google Scholar]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. A multilocus phylogeny of the Metarhizium anisopliae lineage. Mycologia 2009, 101, 512–530. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Sung, G.H.; Ban, S.; Nakagiri, A.; Chen, M.J.; Huang, B.; Li, Z.; Spatafora, J.W. New teleomorph combinations in the entomopathogenic genus Metacordyceps. Mycologia 2012, 104, 182–197. [Google Scholar] [CrossRef]
- Luangsa-ard, J.J.; Mongkolsamrit, S.; Thanakitpipattana, D.; Khonsanit, A.; Tasanathai, K.; Noisripoom, W.; Humber, R.A. Clavicipitaceous entomopathogens: New species in Metarhizium and a new genus Nigelia. Mycol. Prog. 2017, 16, 369–391. [Google Scholar] [CrossRef]
- Zimmermann, G. The entomopathogenic fungus Metarhizium anisopliae and its potential as a biocontrol agent. Pestic. Sci. 1993, 37, 375–379. [Google Scholar] [CrossRef]
- Oreste, M.; Bubici, G.; Poliseno, M.; Tarasco, E. Effect of Beauveria bassiana and Metarhizium anisopliae on the Trialeurodes vaporariorum-Encarsia formosa system. J. Pest Sci. 2016, 89, 153–160. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J. Methuen Handbook of Colour Fletcher; Fletcher & Son: Norwich, UK, 1981; pp. 1–252. [Google Scholar]
- Wang, W.J.; Wang, X.L.; Li, Y.; Xiao, S.R.; Kepler, R.M.; Yao, Y.J. Molecular and morphological studies of Paecilomyces sinensis reveal a new clade in clavicipitaceous fungi and its new systematic position. Syst. Biodivers. 2012, 10, 221–232. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 31, 315–322. [Google Scholar]
- Vilgalys, R.; Sun, B.L. Ancient and recent patterns of geographic speciation in the oyster mushroom Pleurotus revealed by phylogenetic analysis of ribosomal DNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 4599–4603. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Castlebury, L.A.; Rossman, A.Y.; Sung, G.H.; Hyten, A.S.; Spatafora, J.W. Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus. Mycol. Res. 2004, 108, 864–872. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef]
- Mongkolsamrit, S.; Khonsanit, A.; Thanakitpipattana, D.; Tasanathai, K.; Noisripoom, W.; Lamlertthon, S.; Himaman, W.; Houbraken, J.; Samson, R.A.; Luangsa-ard, J. Revisiting Metarhizium and the description of new species from Thailand. Stud. Mycol. 2020, 95, 171–251. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Bischoff, J.F.; Chaverri, P.; White, J.F. Clarification of the host substrate of Ascopolyporus and description of Ascopolyporus philodendrus sp. nov. Mycologia 2005, 97, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.; Seifert, K.; Huhndorf, S.; Robert, V.; Spouge, J.; Levesque, C.; Chen, W.; Fungal Barcoding Consortium. The internal transcribed spacer as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; White, J.F. Phylogenetic evidence for an animal pathogen origin of ergot and the grass endophytes. Mol. Ecol. 2007, 16, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, H.; Tokiwa, T.; Shiina, M.; Asami, Y.; Shiomi, K.; Omura, S.; Nonaka, K. Metarhizium aciculare sp. nov. for euvesperins A and B producing Metarhizium strains. Mycoscience 2019, 60, 313–318. [Google Scholar] [CrossRef]
- Chen, Z.H.; Yang, X.N.; Sun, N.J.; Xu, L.; Zheng, Y.; Yang, Y.M. Species diversity and vertical distribution characteristics of Metarhizium in Gaoligong Mountains, southwestern China. Biodivers. Sci. 2018, 26, 1308–1317. [Google Scholar] [CrossRef]
- Wen, T.C.; Xiao, Y.P.; Han, Y.; Huang, S.K.; Zha, L.S.; Hyde, K.; Kang, J.C. Multigene phylogeny and morphology reveal that the Chinese medicinal mushroom ‘Cordyceps gunnii’ is Metacordyceps neogunnii sp. nov. Phytotaxa 2017, 302, 27. [Google Scholar] [CrossRef]
- Lopes, R.B.; Souza, D.A.; Rocha, L.F.N.; Montalva, C.; Luz, C.; Humber, R.A.; Faria, M. Metarhizium alvesii sp. nov.: A new member of the Metarhizium anisopliae species complex. J. Invertebr. Pathol. 2018, 151, 165–168. [Google Scholar] [CrossRef]
- Gutierrez, A.C.; Leclerque, A.; Manfrino, R.G.; Luz, C.; Ferrari, W.A.O.; Barneche, J.; Garcia, J.J.; Lastra, C.C.L. Natural occurrence in Argentina of a new fungal pathogen of cockroaches, Metarhizium argentinense sp. nov. Fungal Biol. 2019, 123, 364–372. [Google Scholar] [CrossRef]
- Chen, Z.H.; Xu, L.; Yang, X.N.; Zhang, Y.G.; Yang, Y.M. Metarhizium baoshanense sp. nov., a new entomopathogen fungus from Southwestern China. Pak. J. Zool. 2018, 50, 1739–1746. [Google Scholar] [CrossRef]
- Nishi, O.; Shimizu, S.; Sato, H. Metarhizium bibionidarum and M. purpureogenum: New species from Japan. Mycol. Prog. 2017, 16, 987–998. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ohmae, M.; Orihara, T. Metarhizium brachyspermum sp. nov. (Clavicipitaceae), a new species parasitic on Elateridae from Japan. Mycoscience 2020, 61, 37–42. [Google Scholar] [CrossRef]
- Kepler, R.M.; Humber, R.A.; Bischoff, J.F.; Rehner, S.A. Clarification of generic and species boundaries for Metarhizium and related fungi through multigene phylogenetics. Mycologia 2014, 106, 811–829. [Google Scholar] [CrossRef]
- Nishi, O.; Iiyama, K.; Yasunaga-Aoki, C.; Shimizu, S. Phylogenetic status and pathogenicity of Metarhizium majus isolated from a fruit beetle larva in Japan. Mycol. Prog. 2015, 14, 10. [Google Scholar] [CrossRef]
- Chen, W.H.; Han, Y.F.; Liang, J.D.; Liang, Z.Q.; Jin, D.C. Metarhizium dendrolimatilis, a novel Metarhizium species parasitic on Dendrolimus sp. larvae. Mycosphere 2017, 8, 31–37. [Google Scholar] [CrossRef]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. Metarhizium frgidum sp. nov.: A cryptic species of M. anisopliae and a member of the M. flavoviride complex. Mycologia 2006, 98, 737–745. [Google Scholar] [CrossRef]
- Chen, Z.H.; Zhang, Y.G.; Yang, X.N.; Chen, K.; Liu, Q.; Xu, L. A new fungus Metarhizium gaoligongense from China. Int. J. Agric. Biol. 2018, 20, 2271–2276. [Google Scholar]
- Sigler, L.; Gibas, C.F.C.; Kokotovic, B.; Bertelsen, M.F. Disseminated mycosis in veiled chameleons (Chamaeleo calyptratus) caused by Chamaeleomyces granulomatis, a new fungus related to Paecilomyces viridis. J. Clin. Microbiol. 2010, 48, 3182–3192. [Google Scholar] [CrossRef] [PubMed]
- Thanakitpipattana, D.; Tasanathai, K.; Mongkolsamrit, S.; Khonsanit, A.; Lamlertthon, S.; Luangsa-ard, J.J. Fungal pathogens occurring on Orthopterida in Thailand. Persoonia 2020, 44, 140–160. [Google Scholar] [CrossRef]
- Luz, C.; Rocha, L.F.N.; Montalva, C.; Souza, D.A.; Botelho, A.; Lopes, R.B.; Faria, M.; Delalibera, I. Metarhizium humberi sp. nov. (Hypocreales: Clavicipitaceae), a new member of the PARB clade in the Metarhizium anisopliae complex from Latin America. J. Invertebr. Pathol. 2019, 166, 9. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.C.; Zha, L.S.; Xiao, Y.P.; Wang, Q.; Kang, J.C.; Hyde, K.D. Metacordyceps shibinensis sp. nov. from larvae of Lepidoptera in Guizhou Province, southwest China. Phytotaxa 2015, 226, 51–62. [Google Scholar] [CrossRef]
- Perdomo, H.; Cano, J.; Gené, J.; García, D.; Hernández, M.; Guarro, J. Polyphasic analysis of Purpureocillium lilacinum isolates from different origins and proposal of the new species Purpureocillium lavendulum. Mycologia 2013, 105, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Sung, G.H.; Hywel-Jones, N.L.; Luangsa-Ard, J.J.; Bischoff, J.F.; Kepler, R.M.; Spatafora, J.W. Systematics and evolution of the genus Torrubiella (Hypocyeales, Ascomycota). Mycol. Res. 2009, 113, 279–289. [Google Scholar] [CrossRef]
- Chaverri, P.; Bischoff, J.F.; Evans, H.C.; Hodge, K.T. Regiocrella, a new entomopathogenic genus with a pycnidial anamorph and its phylogenetic placement in the Clavicipitaceae. Mycologia 2005, 97, 1225–1237. [Google Scholar] [CrossRef]
- Kepler, R.M.; Sung, G.H.; Harada, Y.; Tanaka, K.; Tanaka, E.; Hosoya, T.; Bischoff, J.F.; Spatafora, J.W. Host jumping onto close relatives and across kingdoms by Tyrannicordyceps (Clavicipitaceae) gen. nov. and Ustilaginoidea_(Clavicipitaceae). Am. J. Bot. 2012, 99, 552–561. [Google Scholar] [CrossRef]
- Sung, G.H.; Shrestha, B.; Han, S.K.; Kim, S.Y.; Sung, J.M. Growth and cultural characteristics of Cordyceps cardinalis collected from Korea. Mycobiology 2010, 38, 274–281. [Google Scholar] [CrossRef]
- Evans, H.C. Entomogenous fungi in tropical forest ecosystems: An appraisal. Ecol. Entomol. 1982, 7, 47–60. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Kitsiou, F.; Natsiopoulos, D.; Eliopoulos, P.A. Entomopathogenic fungi: Interactions and applications. Encyclopedia 2022, 2, 646–656. [Google Scholar] [CrossRef]
- Wang, Y.H.; Ban, S.; Wang, W.J.; Li, Y.; Wang, K.; Kirk, P.M.; Yao, Y.J. Pleurocordyceps gen. nov. for a clade of fungi previously included in Polycephalomyces based on molecular phylogeny and morphology. J. Syst. Evol. 2021, 59, 1065–1080. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhao, X.-C.; Wu, L.-X.; Wang, Y.; Xu, A.; Lin, W.-F. Blackwellomyces kaihuaensis and Metarhizium putuoense (Hypocreales), Two New Entomogenous Fungi from Subtropical Forests in Zhejiang Province, Eastern China. Forests 2023, 14, 2333. https://doi.org/10.3390/f14122333
Li Y, Zhao X-C, Wu L-X, Wang Y, Xu A, Lin W-F. Blackwellomyces kaihuaensis and Metarhizium putuoense (Hypocreales), Two New Entomogenous Fungi from Subtropical Forests in Zhejiang Province, Eastern China. Forests. 2023; 14(12):2333. https://doi.org/10.3390/f14122333
Chicago/Turabian StyleLi, Yi, Xin-Chang Zhao, Li-Xia Wu, Yu Wang, Ao Xu, and Wen-Fei Lin. 2023. "Blackwellomyces kaihuaensis and Metarhizium putuoense (Hypocreales), Two New Entomogenous Fungi from Subtropical Forests in Zhejiang Province, Eastern China" Forests 14, no. 12: 2333. https://doi.org/10.3390/f14122333