
Crinipellis deutziae, Marasmius pinicola spp. nov., and C. rhizomaticola (Agaricales, Basidiomycota) New to China from Beijing

 and
and
Abstract
:1. Introduction
2. Materials and Methods
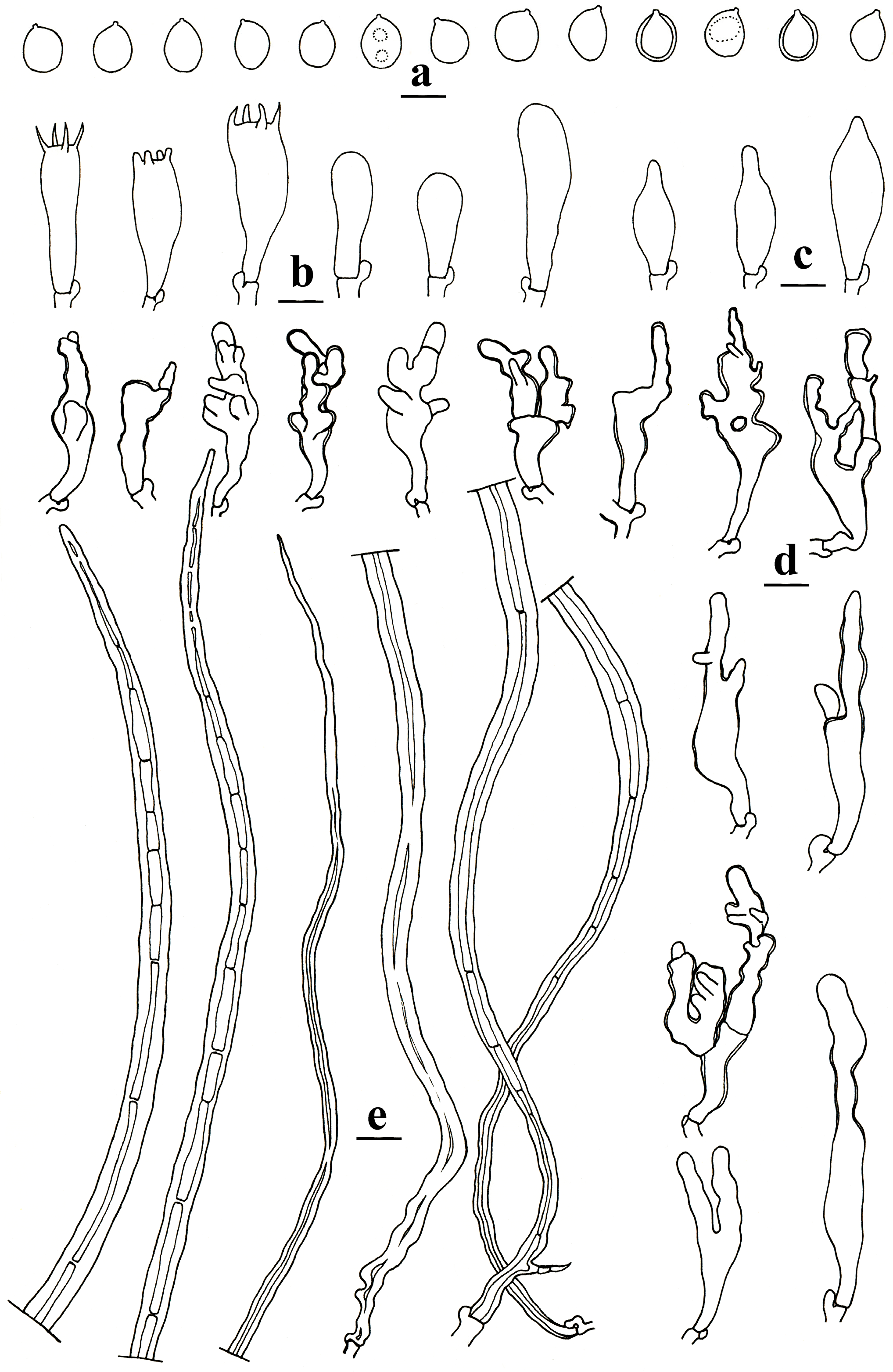
2.1. Morphological Studies
2.2. DNA Extraction and Sequencing
2.3. Phylogenetic Analyses
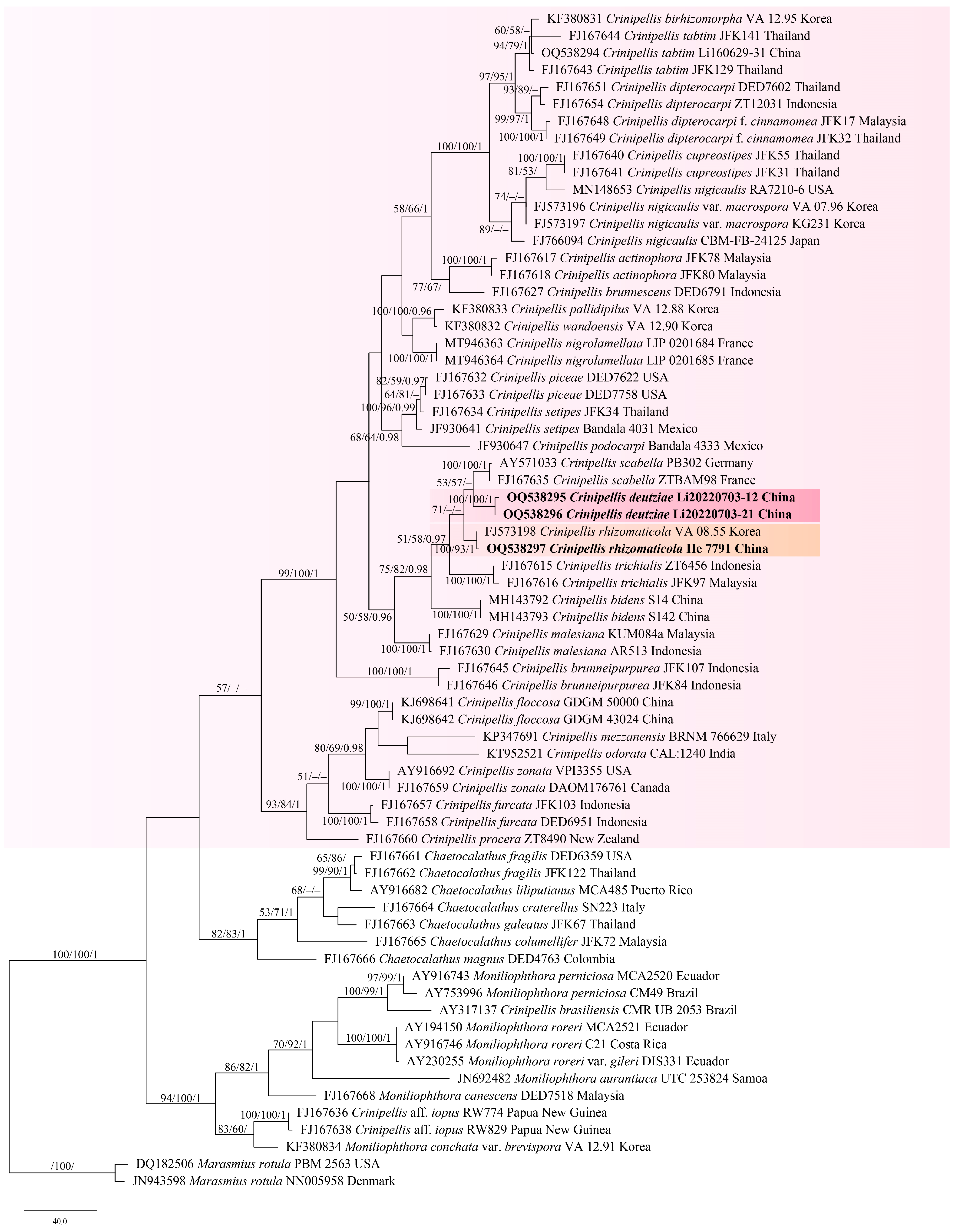
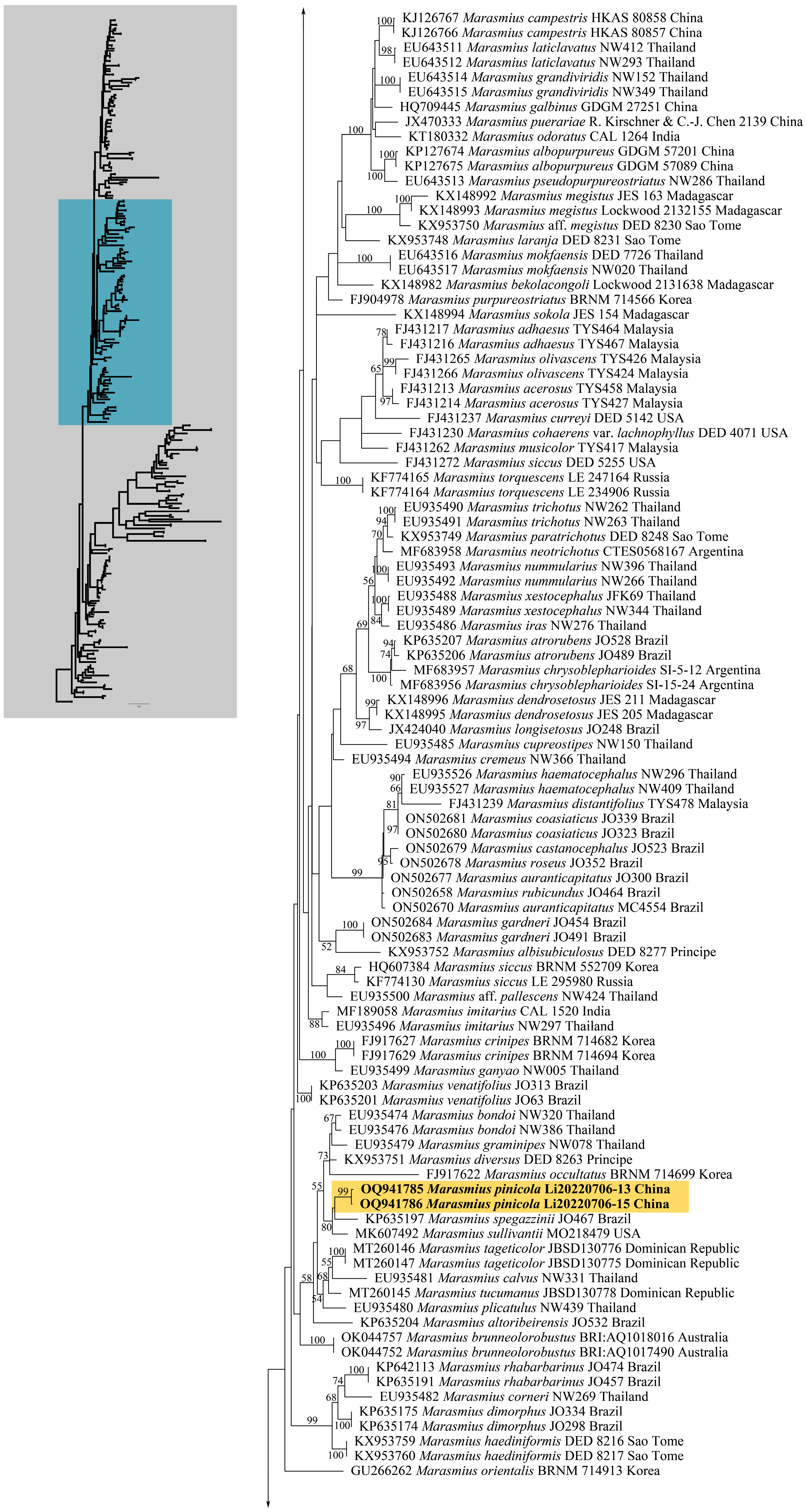
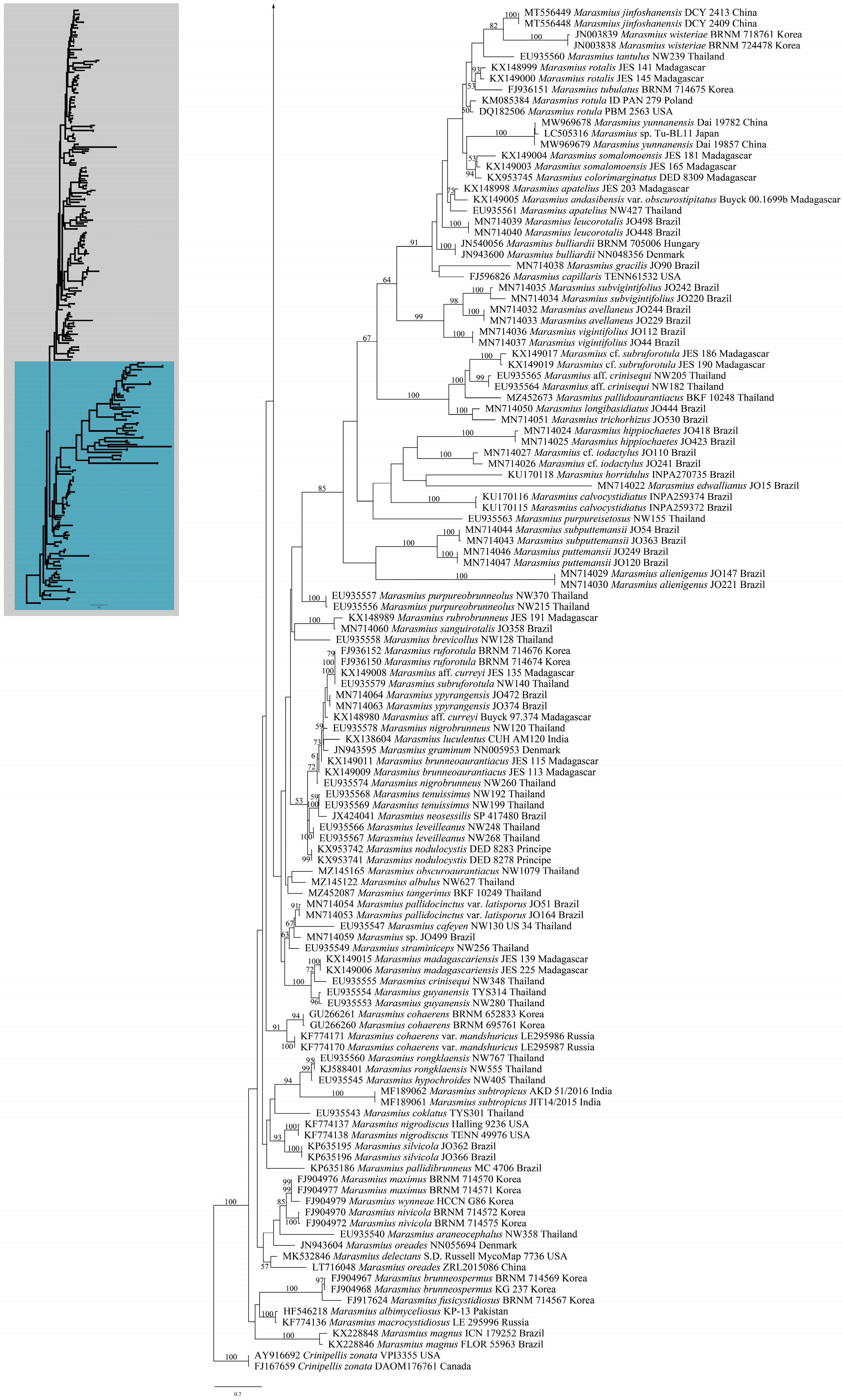
3. Results
3.1. Phylogenetic Analyses
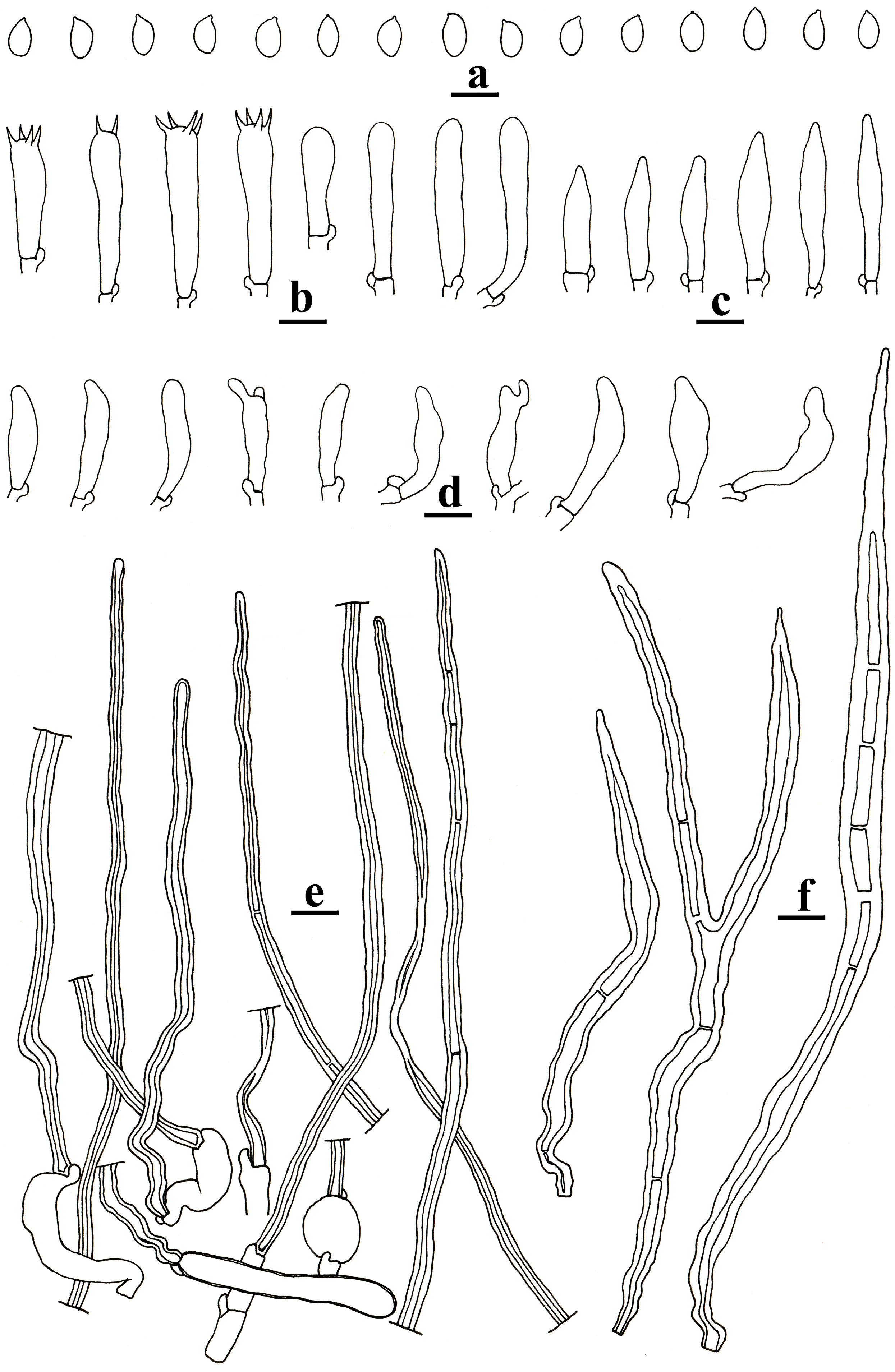
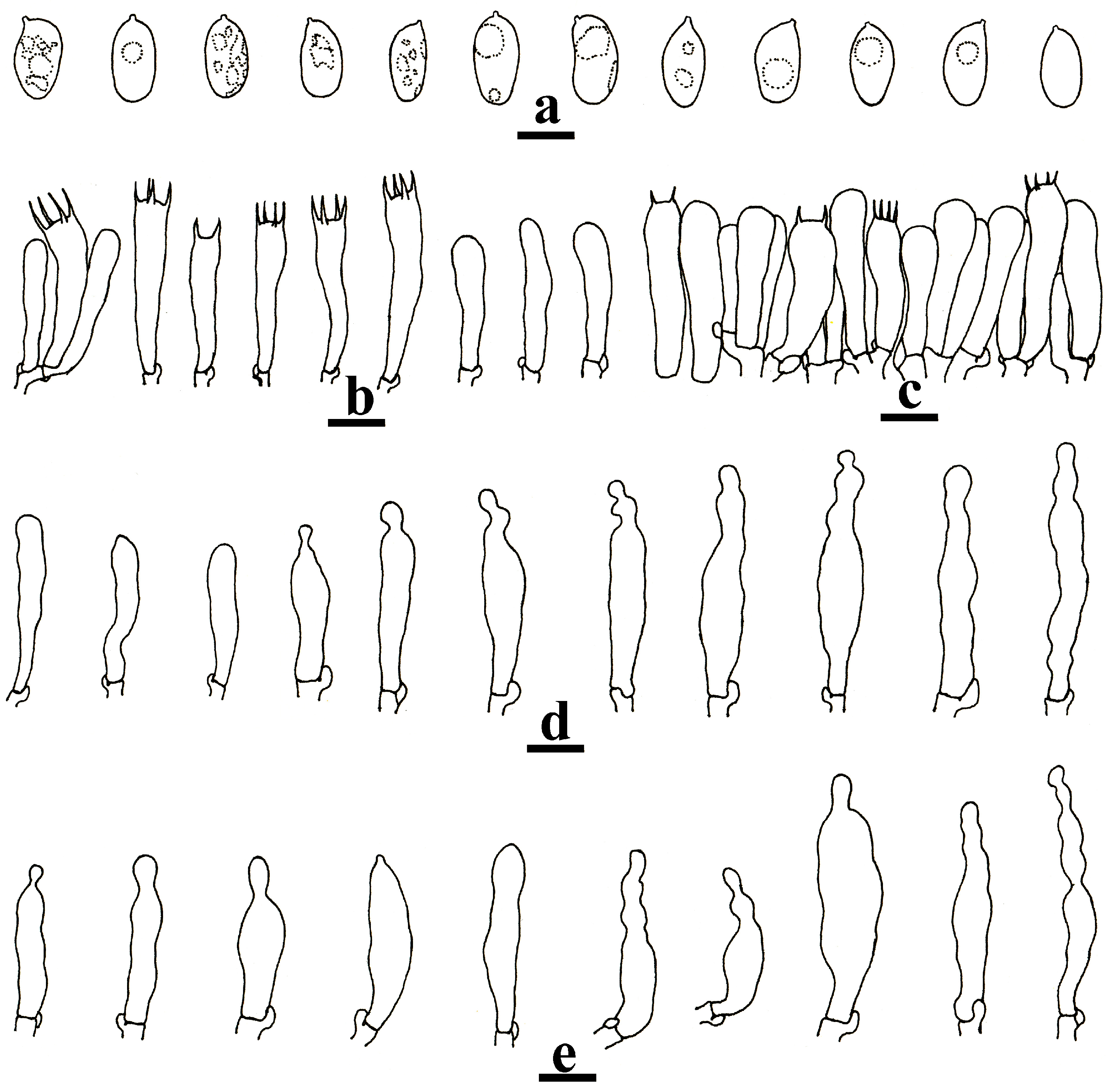
3.2. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kerekes, J.F.; Desjardin, D.E. A monograph of the genera Crinipellis and Moniliophthora from Southeast Asia including a molecular phylogeny of the nrITS region. Fungal Diver. 2009, 37, 101–152. [Google Scholar]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Koenigstein, Germany, 1986. [Google Scholar]
- Takahashi, H. Three new species of Crinipellis found in Iriomote Island, southwestern Japan, and central Honshu, Japan. Mycoscience 2000, 41, 171–182. [Google Scholar] [CrossRef]
- Takahashi, H. Four new species of Crinipellis and Marasmius in eastern Honshu, Japan. Mycoscience 2002, 43, 343–350. [Google Scholar] [CrossRef]
- Antonín, V.; Ryoo, R.; Shin, H.D. Marasmioid and gymnopoid fungi of the Republic of Korea. 1. Three interesting species of Crinipellis (Basidiomycota, Marasmiaceae). Mycotaxon 2009, 108, 429–440. [Google Scholar] [CrossRef]
- Antonín, V.; Ryoo, R.; Ka, K.H.; Sou, H.D. Three new species of Crinipellis and one new variety of Moniliophthora (Basidiomycota, Marasmiaceae) described from the Republic of Korea. Phytotaxa 2014, 170, 86–102. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.W.; Li, T.H.; Deng, W.Q.; Xu, J. A new Crinipellis species with floccose squamules from China. Mycoscience 2015, 56, 476–480. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.S.J.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef]
- Liu, L.N.; Razaq, A.; Atri, N.S.; Bau, T.; Belbahri, L.; Bouket, A.C.; Chen, L.P.; Deng, C.; Ilyas, S.; Khalid, A.N.; et al. Fungal systematics and evolution: FUSE 4. Sydowia 2018, 70, 211–286. [Google Scholar]
- Singer, R. Marasmieae (Basidiomycetes–Tricholomataceae). Flora Neotrop. Monogr. 1976, 17, 1–347. [Google Scholar]
- Desjardin, D.E. The Genus Marasmius from the Southern Appalachian Mountains. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, 1989. [Google Scholar]
- Wilson, A.W.; Desjardin, D.E. Phylogenetic relationships in the gymnopoid and marasmioid fungi (Basidiomycetes, euagaric clade). Mycologia 2005, 97, 667–679. [Google Scholar] [CrossRef]
- Antonín, V. Monograph of Marasmius, Gloiocephala, Palaeocephala and Setulipes in tropical Africa. Fungus Flora Trop. Afr. Meise 2007, 1, 1–164. [Google Scholar]
- Antonín, V.; Noordeloos, M.E. A Monograph of Marasmioid and Collybioid Fungi in Europe; IHW-Verlag: Eching, Germany, 2010. [Google Scholar]
- Lodge, D.J.; Chapela, I.; Samuels, G.; Uecker, F.A.; Desjardin, D.; Horak, E.; Milller, O.K.; Hennebert, G.L.; Decock, C.A.; Ammirati, J.; et al. A survey of patterns of diversity in non-lichenized fungi. Mitt. Eidgenöss. Forschungsanstalt Für Wald Schnee Und Landsch. 1995, 70, 157–173. [Google Scholar]
- Wannathes, N.; Desjardin, D.E.; Hyde, K.D.; Perry, B.A.; Lumyong, S. A monograph of Marasmius (Basidiomycota) from Northern Thailand based on morphological and molecular (ITS sequences) data. Fungal Divers. 2009, 37, 209–306. [Google Scholar]
- Deng, S.F. Taxonomy of Gymnopus and Preliminary Study of Marasmiaceae Resource in South China. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2016. [Google Scholar]
- Zhang, Q.Y.; Si, J.; Li, H.J. A new Marasmius species (Agaricales, Marasmiaceae) with sessile basidiomata growing on wood, from Yunnan, China. Phytotaxa 2023, 578, 169–179. [Google Scholar] [CrossRef]
- Jenkinson, T.S.; Perry, B.A.; Schaefer, R.E.; Desjardin, D.E. Cryptomarasmius gen. nov. is established in the Physalacriaceae to accommodate members of Marasmius section Hygrometrici. Mycologia 2014, 106, 86–94. [Google Scholar] [CrossRef]
- He, M.Q.; Hyde, K.D.; Wei, S.L.; Xi, Y.L.; Cheewangkoon, R.; Zhao, R.L. Three new species of Agaricus section Minores from China. Mycosphere 2018, 9, 189–201. [Google Scholar] [CrossRef]
- Chen, C.C.; Chen, C.Y.; Wu, S.H. Species diversity, taxonomy and multi–gene phylogeny of phlebioid clade (Phanerochaetaceae, Irpicaceae, Meruliaceae) of Polyporales. Fungal Divers. 2021, 111, 337–442. [Google Scholar] [CrossRef]
- Ling, Z.L.; Zhou, J.L.; Parra, L.A.; De Kesel, A.; Callac, P.; Cao, B.; He, M.Q.; Zhao, R.L. Four new species of Agaricus subgenus Spissicaules from China. Mycologia 2021, 113, 476–491. [Google Scholar] [CrossRef]
- Zhou, H.; Li, J.Q.; Hou, C.L. Two new species of the genus Geastrum from the Yanshan Montains in China. Mycosystema 2022, 41, 1–16. [Google Scholar] [CrossRef]
- Li, Y.; He, S.H. Taxonomy and phylogeny of brown-rot corticioid fungi in China: Coniophora beijingensis and Veluticeps subfasciculata spp. nov. Front. Microbiol. 2023, 14, 1133236. [Google Scholar] [CrossRef]
- Li, Y.; Chen, C.C.; He, S.H. New corticioid taxa in Phanerochaetaceae (Polyporales, Basidiomycota) from East Asia. Front. Microbiol. 2023, 14, 1093096. [Google Scholar] [CrossRef] [PubMed]
- Man, X.W.; Dai, Y.C.; Bian, L.S.; Zhou, M.; Zhao, H.; Vlasák, J. Two new species of Haploporus (Polyporales, Basidiomycota) from China and Ecuador based on morphology and phylogeny. Front. Cell. Infect. Microbiol. 2023, 13, 1133839. [Google Scholar] [CrossRef]
- Petersen, J.H. The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Italy, 1996. [Google Scholar]
- Tang, L.X.; Zhang, Y.Z.; Liang, J.Q.; Jiang, S.F.; Si, J.; Li, H.J. Pyrofomes lagerstroemiae (Polyporaceae, Basidiomycota), a new species from Hubei Province, Central China. Phytotaxa 2023, 583, 191–198. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, bbx108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, R. A monographic study of the genera Crinipellis and Chaetocalathus. Lilloa 1942, 8, 441–534. [Google Scholar]
- Wannathes, N.; Desjardin, D.E.; Lumyong, S. Four new species of Marasmius section Globulares from Northern Thailand. Fungal Divers. 2009, 36, 155–163. [Google Scholar]
- Antonín, V.; Ryoo, R.; Shin, H.D. Marasmioid and gymnopoid fungi of the Republic of Korea. 2. Marasmius sect. Globulares. Persoonia 2010, 24, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.J.S.; Moncalvo, J.M.; Margaritescu, S.; Capelari, M. A morphological and phylogenetic evaluation of Marasmius sect. Globulares (Globulares-Sicci complex) with nine new taxa from the Neotropical Atlantic Forest. Persoonia 2020, 44, 240–277. [Google Scholar] [CrossRef]
- Treu, R. Marasmius oreades. IMI descriptions of fungi and bacteria. Mycopathologia 1996, 134, 39–60. [Google Scholar] [CrossRef]
- Takahashi, H. Two new species of Marasmius from eastern Honshu, Japan. Mycoscience 2000, 41, 539–543. [Google Scholar] [CrossRef]
- Nakasone, K.K.; Hibbett, D.S.; Goranova, G. Neocampanella, a new corticioid fungal genus, and a note on Dendrothele bispora. Botany 2009, 87, 875–882. [Google Scholar] [CrossRef] [Green Version]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Voucher | Locality | ITS |
|---|---|---|---|
| Crinipellis actinophora | JFK78 (KLU-M) | Malaysia | FJ167617 |
| C. actinophora | JFK80 (KLU-M) | Malaysia | FJ167618 |
| C. bidens | S14 | China | MH143792 |
| C. bidens | S142 | China | MH143793 |
| C. birhizomorpha | VA 12.95 (BRNM751593) | Korea | KF380831 |
| C. brasiliensis | CMR UB 2053 | Brazil | AY317137 |
| C. brunneipurpurea | JFK107 (KLU-M) | Indonesia | FJ167645 |
| C. brunneipurpurea | JFK84 (KLU-M) | Indonesia | FJ167646 |
| C. brunnescens | DED6791 (BO) | Indonesia | FJ167627 |
| C. cupreostipes | JFK55 (CMU) | Thailand | FJ167640 |
| C. cupreostipes | JFK31 (CMU) | Thailand | FJ167641 |
| C. deutziae | Li20220703-12 * | China | OQ538295 |
| C. deutziae | Li20220703-21 | China | OQ538296 |
| C. dipterocarpi | DED7602 (BO) | Thailand | FJ167651 |
| C. dipterocarpi | ZT12031(BO) | Indonesia | FJ167654 |
| C. dipterocarpi f. cinnamomea | JFK17 (KLU-M) | Malaysia | FJ167648 |
| C. dipterocarpi f. cinnamomea | JFK32 (KLU-M) | Thailand | FJ167649 |
| C. floccosa | GDGM 50000 | China | KJ698641 |
| C. floccosa | GDGM 43024 | China | KJ698642 |
| C. furcata | JFK103 (BO) | Indonesia | FJ167657 |
| C. furcata | DED6951 (BO) | Indonesia | FJ167658 |
| C. aff. iopus | RW774 (GENT) | Papua New Guinea | FJ167636 |
| C. aff. iopus | RW829 (GENT) | Papua New Guinea | FJ167638 |
| C. malesiana | KUM084a (KLU-M) | Malaysia | FJ167629 |
| C. malesiana | AR513 (BO) | Indonesia | FJ167630 |
| C. mezzanensis | BRNM 766629 | Italy | KP347691 |
| C. nigicaulis | CBM-FB-24125 | Japan | FJ766094 |
| C. nigicaulis | RA7210-6 | USA | MN148653 |
| C. nigicaulis var. macrospora | VA07.96 (BRNM712569) | Korea | FJ573196 |
| C. nigicaulis var. macrospora | KG231 (BRNM712580) | Korea | FJ573197 |
| C. nigrolamellata | LIP 0201684 | France | MT946363 |
| C. nigrolamellata | LIP 0201685 | France | MT946364 |
| C. odorata | CAL:1240 | India | KT952521 |
| C. pallidipilus | VA 12.88 (BRNM751595) | Korea | KF380833 |
| C. piceae | DED7622 (SFSU) | USA | FJ167632 |
| C. piceae | DED7758 (SFSU) | USA | FJ167633 |
| C. podocarpi | Bandala 4333 | Mexico | JF930647 |
| C. procera | ZT8490 (ZT) | New Zealand | FJ167660 |
| C. rhizomaticola | VA 08.55 (BRNM712570) | Korea | FJ573198 |
| C. rhizomaticola | He 7791 | China | OQ538297 |
| C. scabella | PB302 | Germany | AY571033 |
| C. scabella | ZTBAM98 (ZT) | France | FJ167635 |
| C. setipes | JFK34 (CMU) | Thailand | FJ167634 |
| C. setipes | Bandala 4031 | Mexico | JF930641 |
| C. tabtim | JFK129 (CMU) | Thailand | FJ167643 |
| C. tabtim | JFK141 (CMU) | Thailand | FJ167644 |
| C. tabtim | Li160629-31 | China | OQ538294 |
| C. trichialis | ZT6456 (ZT) | Indonesia | FJ167615 |
| C. trichialis | JFK97 (SFSU) | Malaysia | FJ167616 |
| C. wandoensis | VA 12.90 (BRNM751594) | Korea | KF380832 |
| C. zonata | VPI3355 | USA | AY916692 |
| C. zonata | DAOM176761 (SFSU) | Canada | FJ167659 |
| Chaetocalathus columellifer | JFK72 (SFSU) | Malaysia | FJ167665 |
| Ch. craterellus | SN223 (ZT) | Italy | FJ167664 |
| Ch. fragilis | DED6359 (SFSU) | Hawaii | FJ167661 |
| Ch. fragilis | JFK122 (SFSU) | Thailand | FJ167662 |
| Ch. galeatus | JFK67 (SFSU) | Thailand | FJ167663 |
| Ch. liliputianus | MCA485 (BPI) | Puerto Rico | AY916682 |
| Ch. magnus | DED4763(SFSU) | Colombia | FJ167666 |
| Moniliophthora aurantiaca | UTC 253824 | Samoa | JN692482 |
| Mo. canescens | DED7518 (SFSU) | Malaysia | FJ167668 |
| Mo. conchata var. brevispora | VA 12.91 (BRNM751596) | Korea | KF380834 |
| Mo. perniciosa | CM49 | Brazil | AY753996 |
| Mo. perniciosa | MCA2520 (BPI) | Ecuador | AY916743 |
| Mo. roreri | MCA2521 (BPI) | Ecuador | AY194150 |
| Mo. roreri | C21 (CATIE) | Costa Rica | AY916746 |
| Mo. roreri var. gileri | DIS331 | Ecuador | AY230255 |
| Marasmius acerosus | TYS458 | Malaysia | FJ431213 |
| M. acerosus | TYS427 | Malaysia | FJ431214 |
| M. adhaesus | TYS467 | Malaysia | FJ431216 |
| M. adhaesus | TYS464 | Malaysia | FJ431217 |
| M. albimyceliosus | KP-13 | Pakistan | HF546218 |
| M. albisubiculosus | DED 8277 | Principe | KX953752 |
| M. albopurpureus | GDGM 57201 | China | KP127674 |
| M. albopurpureus | GDGM 57089 | China | KP127675 |
| M. albulus | NW627 | Thailand | MZ145122 |
| M. alienigenus | JO221 | Brazil | MN714030 |
| M. alienigenus | JO147 | Brazil | MN714029 |
| M. altoribeirensis | JO532 | Brazil | KP635204 |
| M. ambicellularis | JO144 | Brazil | KP635181 |
| M. andasibensis var. obscurostipitatus | Buyck 00.1699b | Madagascar | KX149005 |
| M. apatelius | NW427 | Thailand | EU935561 |
| M. apatelius | JES 203 | Madagascar | KX148998 |
| M. araneocephalus | NW358 | Thailand | EU935540 |
| M. araucariae var. siccipes | NW364 | Thailand | EU935511 |
| M. atrorubens | JO489 | Brazil | KP635206 |
| M. atrorubens | JO528 | Brazil | KP635207 |
| M. auranticapitatus | MC4554 | Brazil | ON502670 |
| M. auranticapitatus | JO300 | Brazil | ON502677 |
| M. aurantioferrugineus | KG 254 | Korea | FJ904962 |
| M. aurantioferrugineus | HCCN 3571 | Korea | FJ904964 |
| M. auratus | NW076 | Thailand | EU935501 |
| M. auratus | NW175 | Thailand | EU935502 |
| M. avellaneus | JO244 | Brazil | MN714032 |
| M. avellaneus | JO229 | Brazil | MN714033 |
| M. bambusiniformis | NW329 | Thailand | EU935521 |
| M. bambusiniformis | NW368 | Thailand | EU935522 |
| M. bekolacongoli | Lockwood 2131638 | Madagascar | KX148982 |
| M. bellus | JO299 | Brazil | KP635208 |
| M. benghalensis | SOUMITRA 240 | India | MF189044 |
| M. benghalensis | SOUMITRA 205 | India | MF189043 |
| M. bondoi | NW320 | Thailand | EU935474 |
| M. bondoi | NW386 | Thailand | EU935476 |
| M. brevicollus | NW128 | Thailand | EU935558 |
| M. brunneoaurantiacus | JES 115 | Madagascar | KX149011 |
| M. brunneoaurantiacus | JES 113 | Madagascar | KX149009 |
| M. brunneolorobustus | BRI:AQ1018016 | Australia | OK044757 |
| M. brunneolorobustus | BRI:AQ1017490 | Australia | OK044752 |
| M. brunneoolivascens | NW112 | Thailand | EU935516 |
| M. brunneoolivascens | NW373 | Thailand | EU935517 |
| M. brunneospermus | BRNM 714569 | Korea | FJ904967 |
| M. brunneospermus | KG 237 | Korea | FJ904968 |
| M. bulliardii | NN048356 | Denmark | JN943600 |
| M. bulliardii | BRNM 705006 | Hungary | JN540056 |
| M. cafeyen | NW130 US 34 | Thailand | EU935547 |
| M. calvocystidiatus | INPA259374 | Brazil | KU170116 |
| M. calvocystidiatus | INPA259372 | Brazil | KU170115 |
| M. calvus | NW331 | Thailand | EU935481 |
| M. campestris | HKAS 80857 | China | KJ126766 |
| M. campestris | HKAS 80858 | China | KJ126767 |
| M. capillaris | TENN61532 | USA | FJ596826 |
| M. castanocephalus | JO523 | Brazil | ON502679 |
| M. chrysoblepharioides | SI-15-24 | Argentina | MF683956 |
| M. chrysoblepharioides | SI-5-12 | Argentina | MF683957 |
| M. cladophyllus var. glaberripes | JO87 | Brazil | KP635163 |
| M. cladophyllus var. glaberripes | JO518 | Brazil | KP635164 |
| M. coarctatus | NW315 | Thailand | EU935541 |
| M. coarctatus | NW385 | Thailand | EU935542 |
| M. coasiaticus | JO323 | Brazil | ON502680 |
| M. coasiaticus | JO339 | Brazil | ON502681 |
| M. cohaerens | BRNM 695761 | Korea | GU266260 |
| M. cohaerens | BRNM 652833 | Korea | GU266261 |
| M. cohaerens var. lachnophyllus | DED 4071 | USA | FJ431230 |
| M. cohaerens var. mandshuricus | LE295987 | Russia | KF774170 |
| M. cohaerens var. mandshuricus | LE295986 | Russia | KF774171 |
| M. coklatus | TYS301 | Thailand | EU935543 |
| M. colorimarginatus | DED 8309 | Madagascar | KX953745 |
| M. conchiformis | JO117 | Brazil | JX424038 |
| M. conchiformis | JO45 | Brazil | KF741996 |
| M. congregatus | JO122 | Brazil | KP635165 |
| M. congregatus | JO468 | Brazil | KP635168 |
| M. corneri | NW269 | Thailand | EU935482 |
| M. corrugatiformis | DED 8233 | Sao Tome | KX953757 |
| M. corrugatiformis | DED 8326 | Sao Tome | KX953756 |
| M. corrugatus | JO336 | Brazil | KP635170 |
| M. corrugatus | JO456 | Brazil | KP635171 |
| M. cremeus | NW366 | Thailand | EU935494 |
| M. crinipes | BRNM 714682 | Korea | FJ917627 |
| M. crinipes | BRNM 714694 | Korea | FJ917629 |
| M. crinisequi | NW348 | Thailand | EU935555 |
| M. aff. crinisequi | NW205 | Thailand | EU935565 |
| M. aff. crinisequi | NW182 | Thailand | EU935564 |
| M. cupreostipes | NW150 | Thailand | EU935485 |
| M. curreyi | DED 5142 | USA | FJ431237 |
| M. aff. curreyi | JES 135 | Madagascar | KX149008 |
| M. aff. curreyi | Buyck 97.374 | Madagascar | KX148980 |
| M. cystidiatus | 1672 | India | MH216042 |
| M. cystidiatus | CAL 1669 | India | MH216191 |
| M. delectans | S.D. Russell MycoMap 7736 | USA | MK532846 |
| M. dendrosetosus | JES 205 | Madagascar | KX148995 |
| M. dendrosetosus | JES 211 | Madagascar | KX148996 |
| M. dimorphus | JO298 | Brazil | KP635174 |
| M. dimorphus | JO334 | Brazil | KP635175 |
| M. distantifolius | TYS478 | Malaysia | FJ431239 |
| M. diversus | DED 8263 | Principe | KX953751 |
| M. edwallianus | JO15 | Brazil | MN714022 |
| M. elaeocephaliformis | DED 8213 | Sao Tome | KX953758 |
| M. elaeocephalus | DED 8254 | Sao Tome | KX953754 |
| M. ferruginoides | JES 209 | Madagascar | KX148983 |
| M. fusicystidiosus | BRNM 714567 | Korea | FJ917624 |
| M. galbinus | GDGM 27251 | China | HQ709445 |
| M. ganyao | NW005 | Thailand | EU935499 |
| M. gardneri | JO491 | Brazil | ON502683 |
| M. gardneri | JO454 | Brazil | ON502684 |
| M. gracilis | JO90 | Brazil | MN714038 |
| M. graminicola | BRNM 714701 | Korea | FJ917621 |
| M. graminicola | BRNM 714685 | Korea | FJ917617 |
| M. graminipes | NW078 | Thailand | EU935479 |
| M. graminum | NN005953 | Denmark | JN943595 |
| M. grandisetulosus | DED 8225 | Sao Tome | KX953743 |
| M. grandisetulosus | DED 8257 | Sao Tome | KX953744 |
| M. grandiviridis | NW152 | Thailand | EU643514 |
| M. grandiviridis | NW349 | Thailand | EU643515 |
| M. griseoroseus | JO465 | Brazil | KJ173479 |
| M. griseoroseus var. diminutus | JO390 | Brazil | JX424044 |
| M. guyanensis | TYS314 | Thailand | EU935554 |
| M. guyanensis | NW280 | Thailand | EU935553 |
| M. haediniformis | DED 8216 | Sao Tome | KX953759 |
| M. haediniformis | DED 8217 | Sao Tome | KX953760 |
| M. haematocephalus | NW409 | Thailand | EU935527 |
| M. haematocephalus | NW296 | Thailand | EU935526 |
| M. hinnuleus | JES 217 | Madagascar | KX148988 |
| M. hippiochaetes | JO423 | Brazil | MN714025 |
| M. hippiochaetes | JO418 | Brazil | MN714024 |
| M. horridulus | INPA270735 | Brazil | KU170118 |
| M. hypochroides | NW405 | Thailand | EU935545 |
| M. hypophaeus | NW285 | Thailand | EU935484 |
| M. imitarius | CAL 1520 | India | MF189058 |
| M. imitarius | NW297 | Thailand | EU935496 |
| M. indojasminodorus | AKD 135/2015 | India | KY785172 |
| M. insolitus | LE 289497 | Russia | KF774162 |
| M. insolitus | LE 289498 | Russia | KF774163 |
| M. inthanonensis | NW353 | Thailand | EU935514 |
| M. cf. iodactylus | JO241 | Brazil | MN714026 |
| M. cf. iodactylus | JO110 | Brazil | MN714027 |
| M. iras | NW276 | Thailand | EU935486 |
| M. jalapensis | 414F | Paraguay | MT441867 |
| M. jasminodorus | NW294 | Thailand | EU935513 |
| M. jasminodorus | NW414 | Thailand | EU935515 |
| M. jinfoshanensis | DCY 2409 | China | MT556448 |
| M. jinfoshanensis | DCY 2413 | China | MT556449 |
| M. katangensis | JES 227 | Madagascar | KX148991 |
| M. koreanus | BRNM 714697 | Korea | FJ917619 |
| M. koreanus | BRNM 714700 | Korea | FJ917620 |
| M. laranja | DED 8231 | Sao Tome | KX953748 |
| M. laticlavatus | NW293 | Thailand | EU643512 |
| M. laticlavatus | NW412 | Thailand | EU643511 |
| M. leoninus | JO320 | Brazil | KP635162 |
| M. leoninus | JO84 | Brazil | KP635209 |
| M. leucorotalis | JO498 | Brazil | MN714039 |
| M. leucorotalis | JO448 | Brazil | MN714040 |
| M. leveilleanus | NW248 | Thailand | EU935566 |
| M. leveilleanus | NW268 | Thailand | EU935567 |
| M. linderioides | JO286 | Brazil | JX424037 |
| M. longibasidiatus | JO444 | Brazil | MN714050 |
| M. longisetosus | JO248 | Brazil | JX424040 |
| M. luculentus | CUH AM120 | India | KX138604 |
| M. luteolus | NW138 | Thailand | EU935506 |
| M. luteolus | NW304 | Thailand | EU935507 |
| M. macrocystidiosus | LE 295996 | Russia | KF774136 |
| M. madagascariensis | JES 225 | Madagascar | KX149006 |
| M. madagascariensis | JES 139 | Madagascar | KX149015 |
| M. magnus | ICN 179252 | Brazil | KX228848 |
| M. magnus | FLOR 55963 | Brazil | KX228846 |
| M. makok | NW201 | Thailand | EU935524 |
| M. margallensis | SIM35 | Pakistan | OP804130 |
| M. margallensis | SIM34 | Pakistan | OP804131 |
| M. maximus | BRNM 714570 | Korea | FJ904976 |
| M. maximus | BRNM 714571 | Korea | FJ904977 |
| M. megistus | JES 163 | Madagascar | KX148992 |
| M. megistus | Lockwood 2132155 | Madagascar | KX148993 |
| M. aff. megistus | DED 8230 | Sao Tome | KX953750 |
| M. mokfaensis | DED 7726 | Thailand | EU643516 |
| M. mokfaensis | NW020 | Thailand | EU643517 |
| M. musicolor | TYS417 | Malaysia | FJ431262 |
| M. neosessilis | SP 417480 | Brazil | JX424041 |
| M. neotrichotus | CTES0568167 | Argentina | MF683958 |
| M. neotropicalis | JO325 | Brazil | KP635185 |
| M. neotropicalis | JO69 | Brazil | KP635183 |
| M. nigrobrunneus | NW260 | Thailand | EU935574 |
| M. nigrobrunneus | NW120 | Thailand | EU935578 |
| M. nigrodiscus | Halling 9236 | USA | KF774137 |
| M. nigrodiscus | TENN 49976 | USA | KF774138 |
| M. nivicola | BRNM 714575 | Korea | FJ904972 |
| M. nivicola | BRNM 714572 | Korea | FJ904970 |
| M. nodulocystis | DED 8278 | Principe | KX953741 |
| M. nodulocystis | DED 8283 | Principe | KX953742 |
| M. nummularius | NW266 | Thailand | EU935492 |
| M. nummularius | NW396 | Thailand | EU935493 |
| M. obscuroaurantiacus | NW1079 | Thailand | MZ145165 |
| M. occultatiformis | VLA M-19639 | Russia | KF774160 |
| M. occultatiformis | LE 295995 | Russia | KF774157 |
| M. occultatus | BRNM 714699 | Korea | FJ917622 |
| M. ochroleucus | LE 295978 | Russia | KF912952 |
| M. odoratus | CAL 1264 | India | KT180332 |
| M. oreades | NN055694 | Denmark | JN943604 |
| M. oreades | ZRL2015086 | China | LT716048 |
| M. orientalis | BRNM 714913 | Korea | GU266262 |
| M. olivascens | TYS424 | Malaysia | FJ431266 |
| M. olivascens | TYS426 | Malaysia | FJ431265 |
| M. aff. pallescens | NW424 | Thailand | EU935500 |
| M. pallidibrunneus | MC 4706 | Brazil | KP635186 |
| M. pallidoaurantiacus | BKF 10248 | Thailand | MZ452673 |
| M. pallidocinctus var. latisporus | JO164 | Brazil | MN714053 |
| M. pallidocinctus var. latisporus | JO51 | Brazil | MN714054 |
| M. paratrichotus | DED 8248 | Sao Tome | KX953749 |
| M. pellucidus | NW321 | Thailand | EU935508 |
| M. pellucidus | NW342 | Thailand | EU935509 |
| M. pinicola | Li20220706-13 | China | OQ941785 |
| M. pinicola | Li20220706-15 * | China | OQ941786 |
| M. plicatulus | NW439 | Thailand | EU935480 |
| M. pseudoniveoaffinis | JO60 | Brazil | KP635187 |
| M. pseudoniveoaffinis | JO70 | Brazil | KP635188 |
| M. pseudopellucidus | NW186 | Thailand | EU935504 |
| M. pseudopellucidus | NW305 | Thailand | EU935505 |
| M. pseudopurpureostriatus | NW286 | Thailand | EU643513 |
| M. puerariae | R. Kirschner & C.-J. Chen 2139 | China | JX470333 |
| M. pulcherripes | RA705-12b | USA | MK217466 |
| M. pulcherripes | iNAT:30809942 | USA | MZ267775 |
| M. purpureisetosus | NW155 | Thailand | EU935563 |
| M. purpureobrunneolus | NW370 | Thailand | EU935557 |
| M. purpureobrunneolus | NW215 | Thailand | EU935556 |
| M. purpureostriatus | BRNM 714566 | Korea | FJ904978 |
| M. puttemansii | JO249 | Brazil | MN714046 |
| M. puttemansii | JO120 | Brazil | MN714047 |
| M. rhabarbarinus | JO457 | Brazil | KP635191 |
| M. rhabarbarinus | JO474 | Brazil | KP642113 |
| M. rhodopurpureus | BRNM 724483 | Korea | HQ607382 |
| M. rongklaensis | NW555 | Thailand | KJ588401 |
| M. rongklaensis | NW767 | Thailand | EU935560 |
| M. roseus | JO352 | Brazil | ON502678 |
| M. rotalis | JES 145 | Madagascar | KX149000 |
| M. rotalis | JES 141 | Madagascar | KX148999 |
| M. rotula | ID PAN 279 | Poland | KM085384 |
| M. rotula | PBM 2563 | USA | DQ182506 |
| M. rotula | NN005958 | Denmark | JN943598 |
| M. ruber | DED 8669 | Brazil | KP635193 |
| M. rubicundus | JO464 | Brazil | ON502658 |
| M. rubrobrunneus | JES 191 | Madagascar | KX148989 |
| M. ruforotula | BRNM 714676 | Korea | FJ936152 |
| M. ruforotula | BRNM 714674 | Korea | FJ936150 |
| M. sanguirotalis | JO358 | Brazil | MN714060 |
| M. siccus | BRNM 552709 | Korea | HQ607384 |
| M. siccus | LE 295980 | Russia | KF774130 |
| M. siccus | DED 5255 | USA | FJ431272 |
| M. silvicola | JO362 | Brazil | KP635195 |
| M. silvicola | JO366 | Brazil | KP635196 |
| M. somalomoensis | JES 181 | Madagascar | KX149004 |
| M. somalomoensis | JES 165 | Madagascar | KX149003 |
| M. sokola | JES 154 | Madagascar | KX148994 |
| M. spegazzinii | JO467 | Brazil | KP635197 |
| M. straminiceps | NW256 | Thailand | EU935549 |
| M. strobiluriformis | BRNM 714914 | Korea | GU266263 |
| M. strobiluriformis | BRNM 714915 | Korea | GU266264 |
| M. subarborescens | DED 8215 | Sao Tome | KX953755 |
| M. subputtemansii | JO363 | Brazil | MN714043 |
| M. subputtemansii | JO54 | Brazil | MN714044 |
| M. subruforotula | NW140 | Thailand | EU935579 |
| M. cf. subruforotula | JES 186 | Madagascar | KX149017 |
| M. cf. subruforotula | JES 190 | Madagascar | KX149019 |
| M. subtangerinus | BRNM 718756 | Korea | HQ607380 |
| M. subtropicus | JIT14/2015 | India | MF189061 |
| M. subtropicus | AKD 51/2016 | India | MF189062 |
| M. subvigintifolius | JO242 | Brazil | MN714035 |
| M. subvigintifolius | JO220 | Brazil | MN714034 |
| M. sullivantii | MO218479 | USA | MK607492 |
| M. suthepensis | JO329 | Brazil | KP635198 |
| M. suthepensis | JO469 | Brazil | KP635199 |
| M. tageticolor | JBSD130776 | Dominican Republic | MT260146 |
| M. tageticolor | JBSD130775 | Dominican Republic | MT260147 |
| M. tangerinus | BKF 10249 | Thailand | MZ452087 |
| M. tantulus | NW239 | Thailand | EU935560 |
| M. tenuissimus | NW192 | Thailand | EU935568 |
| M. tenuissimus | NW199 | Thailand | EU935569 |
| M. torquescens | LE 234906 | Russia | KF774164 |
| M. torquescens | LE 247164 | Russia | KF774165 |
| M. trichorhizus | JO530 | Brazil | MN714051 |
| M. trichotus | NW262 | Thailand | EU935490 |
| M. trichotus | NW263 | Thailand | EU935491 |
| M. tricystidiatus | NR 100 | Argentina | MT441866 |
| M. tricystidiatus | CB T6-02 | Argentina | MF683959 |
| M. trinitatis | JO306 | Brazil | KP635200 |
| M. tubulatus | BRNM 714675 | Korea | FJ936151 |
| M. tucumanus | JBSD130778 | Dominican Republic | MT260145 |
| M. venatifolius | JO63 | Brazil | KP635201 |
| M. venatifolius | JO313 | Brazil | KP635203 |
| M. vigintifolius | JO112 | Brazil | MN714036 |
| M. vigintifolius | JO44 | Brazil | MN714037 |
| M. wisteriae | BRNM 724478 | Korea | JN003838 |
| M. wisteriae | BRNM 718761 | Korea | JN003839 |
| M. wynneae | HCCN G86 | Korea | FJ904979 |
| M. xestocephalus | JFK69 | Thailand | EU935488 |
| M. xestocephalus | NW344 | Thailand | EU935489 |
| M. ypyrangensis | JO472 | Brazil | MN714064 |
| M. ypyrangensis | JO374 | Brazil | MN714063 |
| M. yunnanensis | Dai 19857 | China | MW969679 |
| M. yunnanensis | Dai 19782 | China | MW969678 |
| Marasmius sp. 1 | Tu-BL11 | Japan | LC505316 |
| Marasmius sp. 2 | JO499 | Brazil | MN714059 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; He, S.-H.; Zhang, Y.-Z.; Liang, J.-Q.; Sheng, W.-Y.; Si, J.; Li, H.-J. Crinipellis deutziae, Marasmius pinicola spp. nov., and C. rhizomaticola (Agaricales, Basidiomycota) New to China from Beijing. Forests 2023, 14, 1480. https://doi.org/10.3390/f14071480
Li Y, He S-H, Zhang Y-Z, Liang J-Q, Sheng W-Y, Si J, Li H-J. Crinipellis deutziae, Marasmius pinicola spp. nov., and C. rhizomaticola (Agaricales, Basidiomycota) New to China from Beijing. Forests. 2023; 14(7):1480. https://doi.org/10.3390/f14071480
Chicago/Turabian StyleLi, Yue, Shuang-Hui He, Yi-Zhe Zhang, Jia-Qi Liang, Wen-Yue Sheng, Jing Si, and Hai-Jiao Li. 2023. "Crinipellis deutziae, Marasmius pinicola spp. nov., and C. rhizomaticola (Agaricales, Basidiomycota) New to China from Beijing" Forests 14, no. 7: 1480. https://doi.org/10.3390/f14071480