
The Multivariate Distribution of Stand Spatial Structure and Tree Size Indices Using Neighborhood-Based Variables in Coniferous and Broad Mixed Forest

Abstract
:1. Introduction
2. Materials and Methods
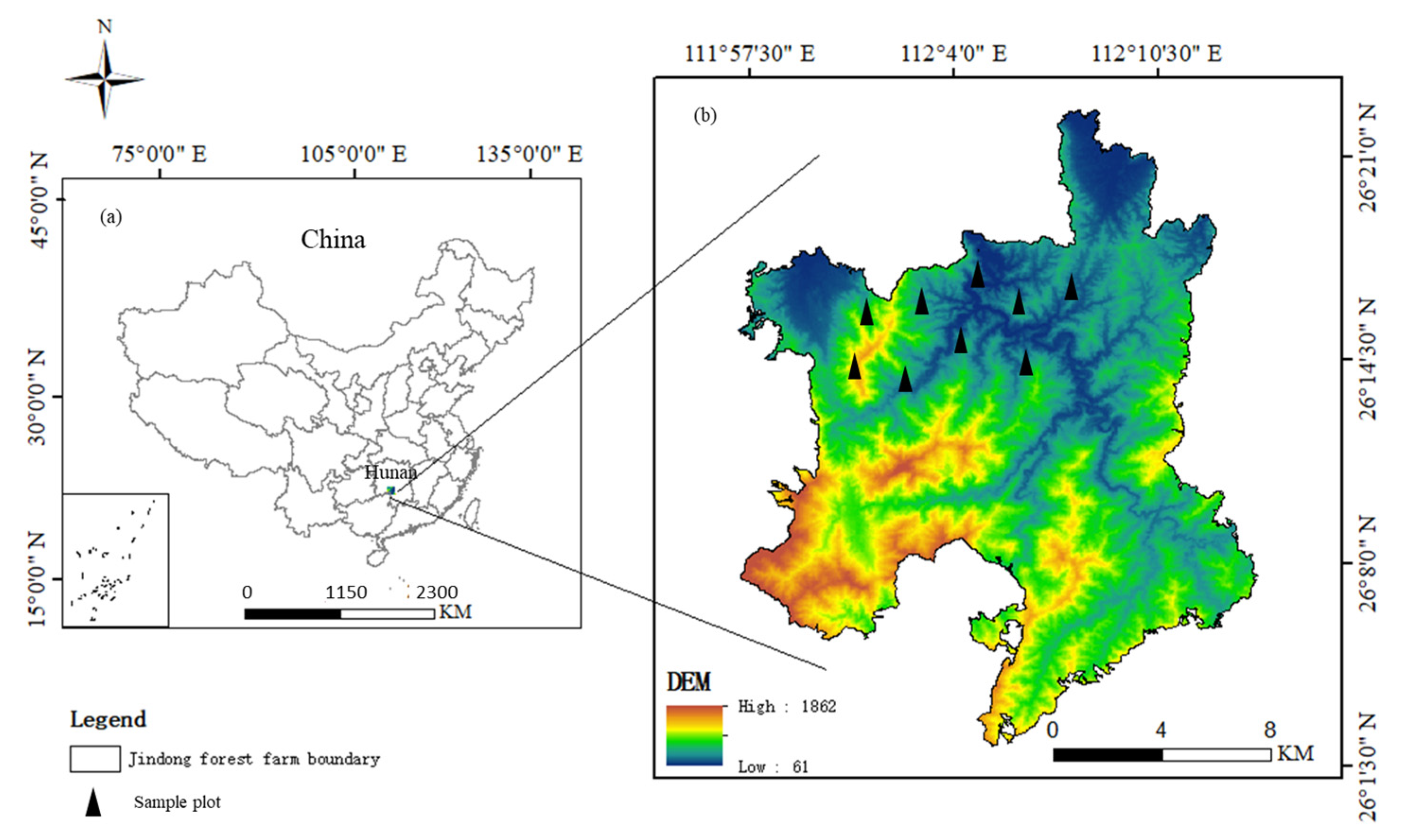
2.1. Study Area
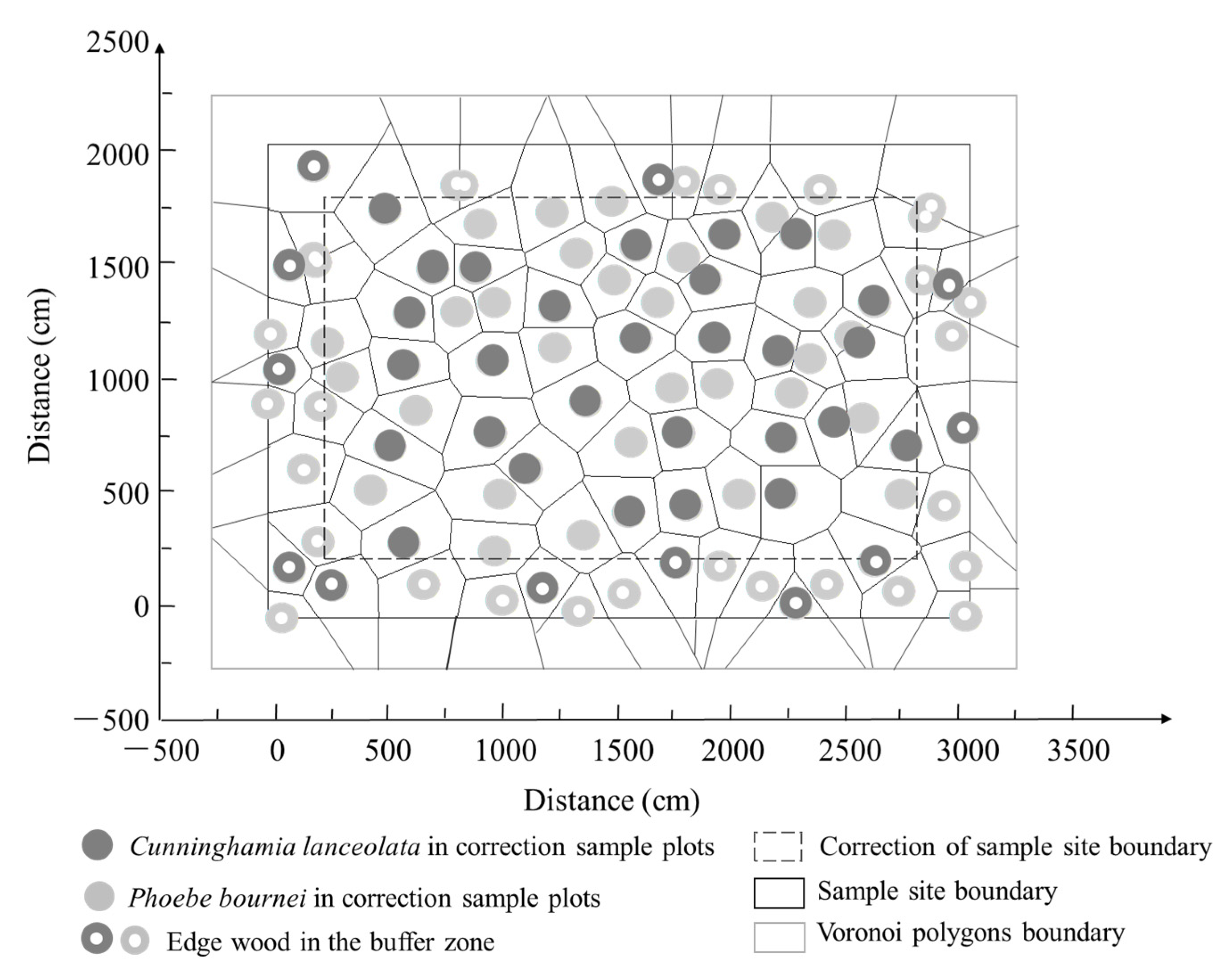
2.2. Study Design and Sampling
2.3. Forest Spatial Structure Parameters
2.4. Coupling Method
2.5. Statistical Analysis
3. Results
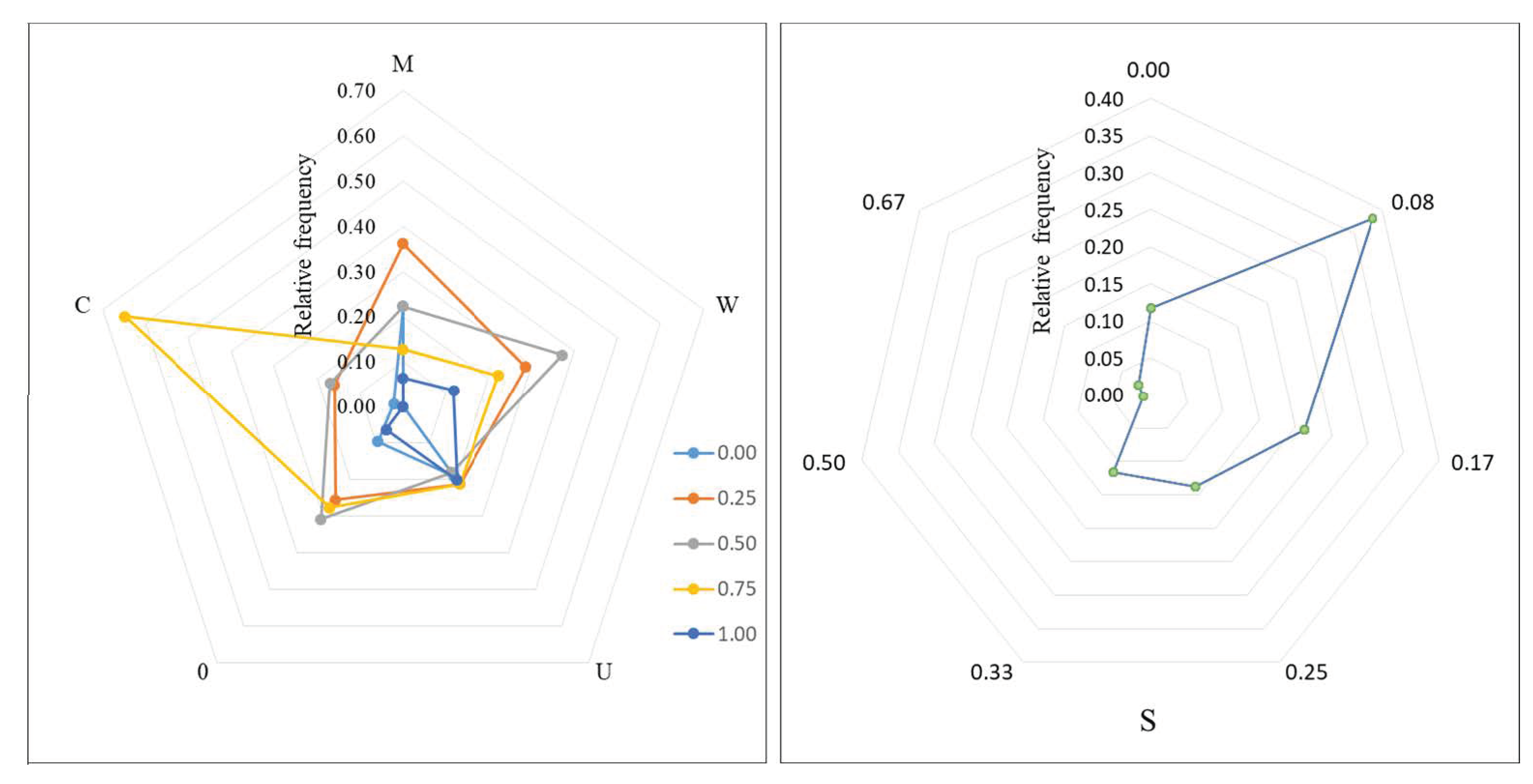
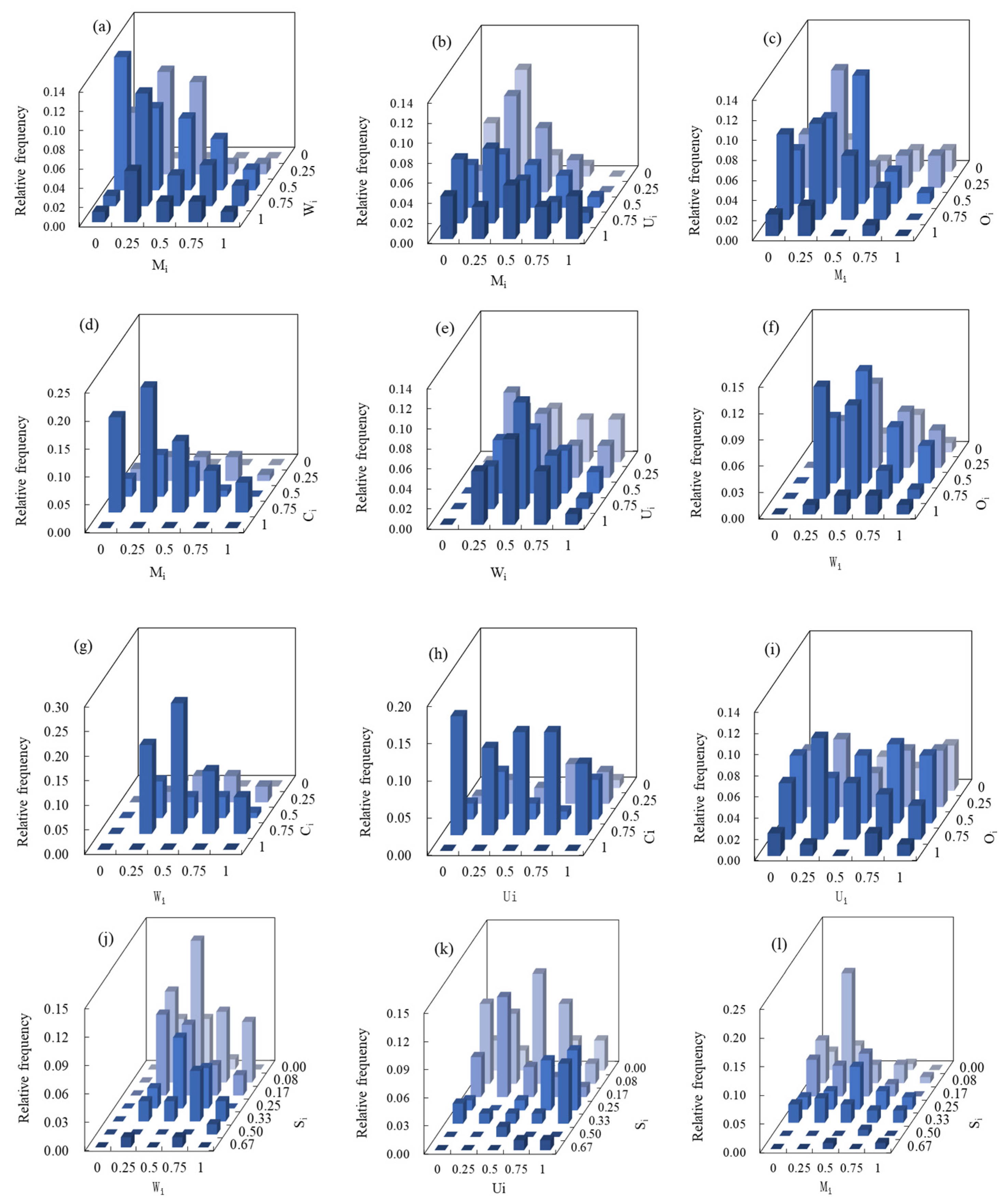
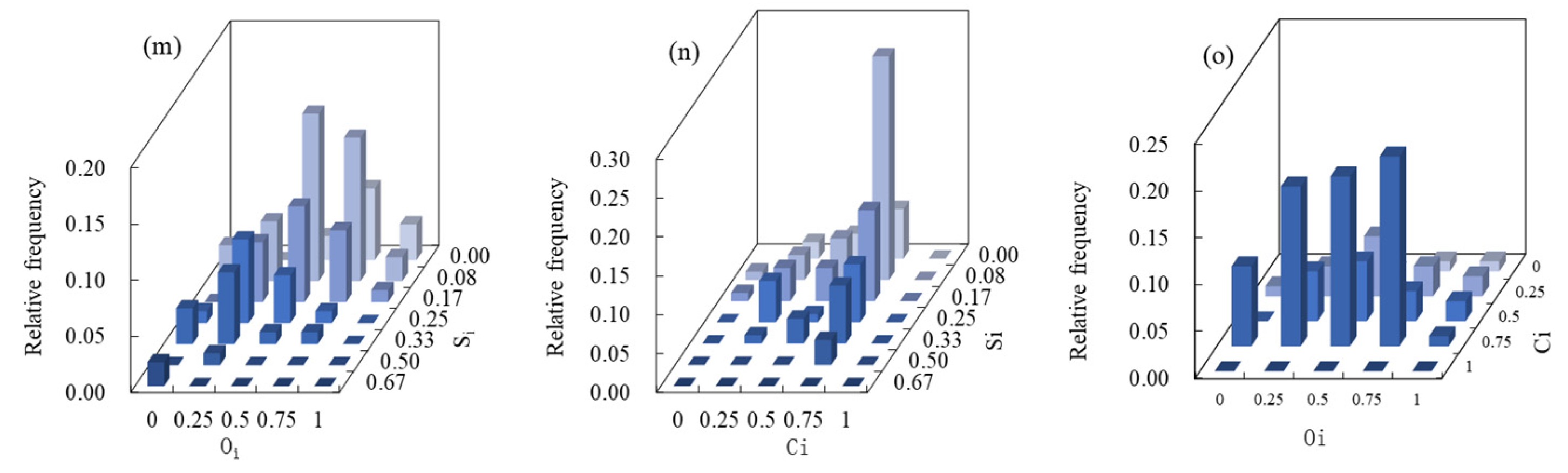
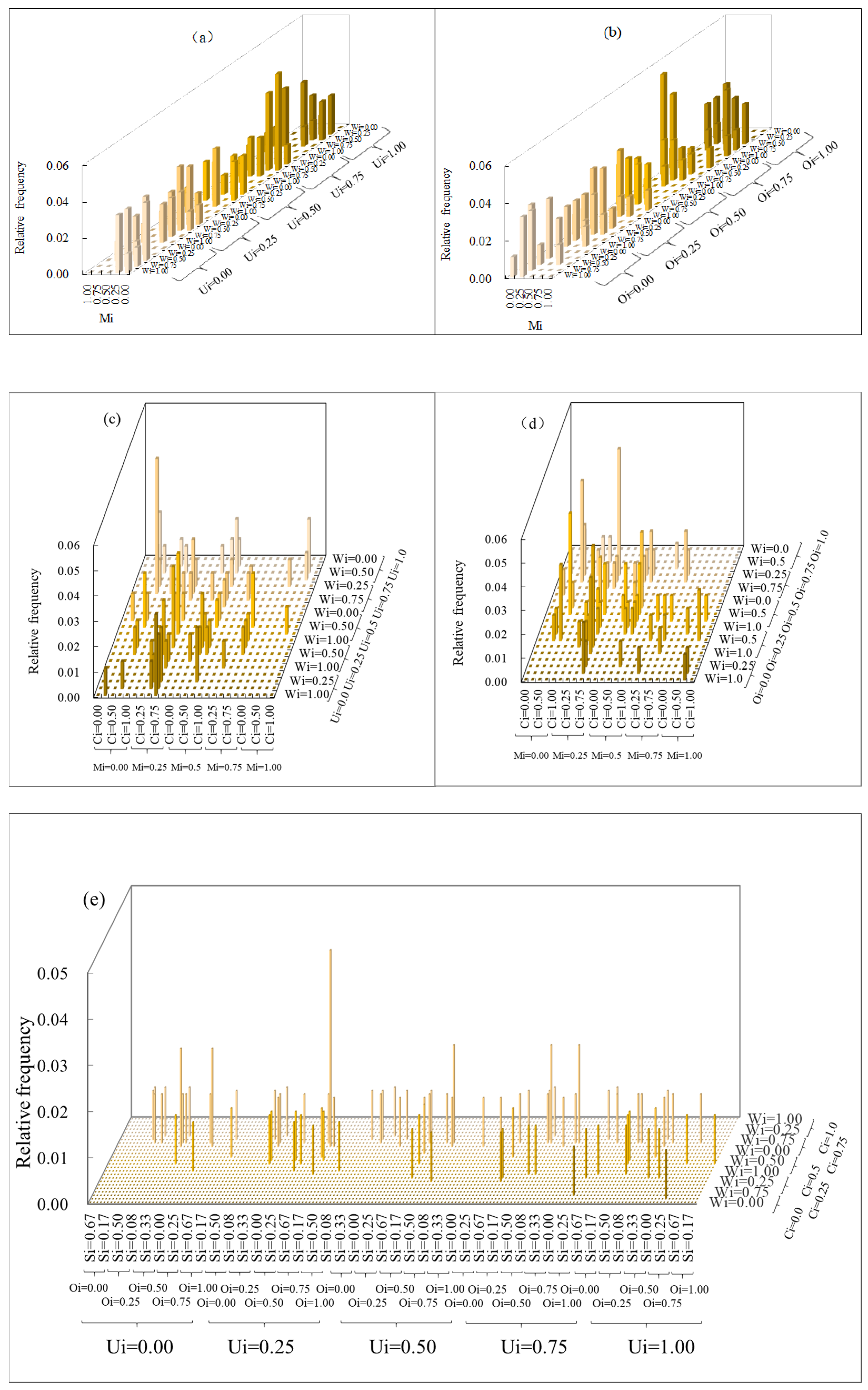
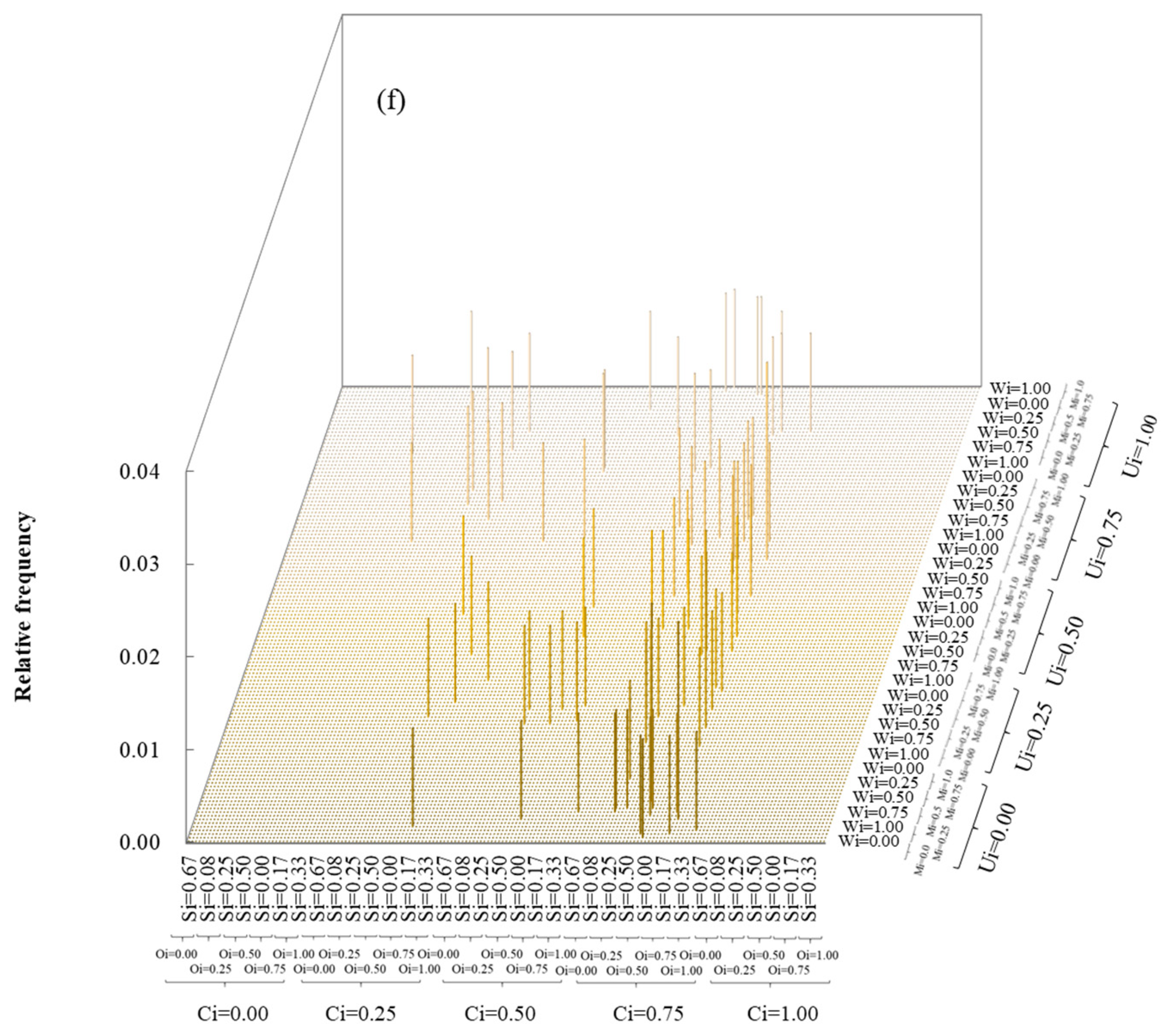
3.1. Multivariate Distribution of Spatial Structure Indices
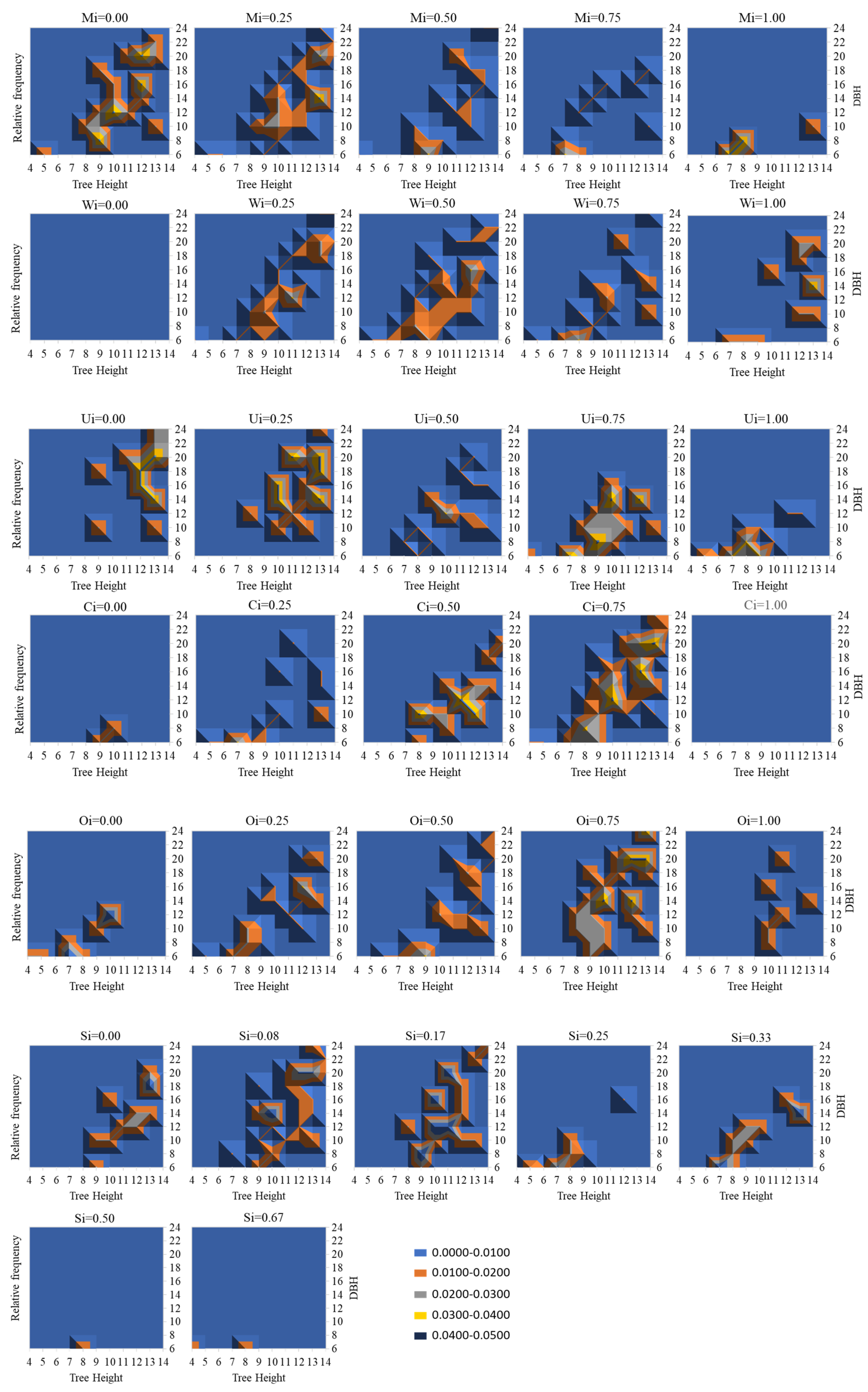
3.2. Coupling of Spatial Structure Indices and Tree Size
4. Discussion
4.1. The Superiority of the Six-Variable Distribution Method Compared to the Univariate Method
4.2. Relationship between Forest Spatial Structure and Tree Size
4.3. The Role of the N-Variable Distribution Method in Forest Structure Adjustment
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ehbrecht, M.; Seidel, D.; Annighöfer, P.; Kreft, H.; Köhler, M.; Zemp, D.C.; Puettmann, K.; Nilus, R.; Babweteera, F.; Willim, K.; et al. Global patterns and climatic controls of forest structural complexity. Nat. Commun. 2021, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and woodland stand structural complexity: Its definition and measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Franklin, J.F.; Fischer, J. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biol. Conserv. 2006, 131, 433–445. [Google Scholar] [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Pastur, G.M.; Messier, C.; et al. Retention Forestry to Maintain Multifunctional Forests: A World Perspective. Bioscience 2012, 62, 633–645. [Google Scholar] [CrossRef]
- Chirici, G.; McRoberts, R.E.; Winter, S.; Barbati, A.; Brändli, U.B.; Abegg, M.; Beranova, J.; Rondeux, J.; Bertini, R.; Alberdi Asensio, I.; et al. National Forest Inventory Contributions to Forest Biodiversity Monitoring. For. Ecol. Manag. 2016, 356, 12–26. [Google Scholar] [CrossRef]
- Coates, K.D.; Haeussler, S. The Use of Stand Structural Attributes in Understanding and Managing Coast Interior Cedar-Hemlock Forests. For. Ecol. Manag. 2003, 172, 229–244. [Google Scholar]
- MacFarlane, D.W.; MacFarlane, D.M.; Keith, D.A. Towards consistent classification of structural forest attributes. For. Ecol. Manag. 2017, 394, 11–20. [Google Scholar]
- Lutz, J.A.; Larson, A.J.; Freund, J.A.; Swanson, M.E.; Bible, K.J. The Importance of Large-Diameter Trees to Forest Structural Heterogeneity. PLoS ONE 2012, 7, e36398. [Google Scholar] [CrossRef]
- Parresol, B.R. Assessing Tree and Stand Biomass: A Review with Examples and Critical Comparisons. For. Sci. 1999, 45, 573–593. [Google Scholar]
- Hui, G.Y.; Hu, Y.B.; Zhao, Z.H. Research Progress in Structured Forest Management. For. Sci. Res. 2018, 31, 85–93. [Google Scholar] [CrossRef]
- Bélisle, C.; Desrochers, A. The Influence of Forest Stand Structure on Bird Communities in Boreal Forests of Quebec: An Assessment at Two Spatial Scales. Can. J. For. Res. 2002, 32, 388–399. [Google Scholar]
- Hui, G.; Gu, H. Enhancing forest diversity by combining different thinning approaches: A case study in northeastern China. For. Ecol. Manag. 2017, 389, 94–102. [Google Scholar]
- Pukkala, T.; Kolström, T. Assessment of structural diversity in uneven-sized stands. For. Ecol. Manag. 2000, 135, 147–155. [Google Scholar]
- Zhang, J.; Kang, S.; Zhang, Y. Spatio-temporal variations of structural diversity of Picea crassifolia forests in the Qilian Mountains, northwestern China. Ecol. Res. 2010, 25, 425–436. [Google Scholar]
- Gadow, K.V.; Hui, G.Y. Modeling Forest Development; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Lefsky, M.A.; Cohen, W.B.; Harding, D.J.; Parker, G.G.; Shugart, H.H.; Gower, S.T. Lidar Remote Sensing for Ecosystem Studies. BioScience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Kint, V.; Clarke, N.; Vesterdal, L. Quantifying fine-root biomass in Scots pine and Norway spruce stands using fine-root diameter and biomass equations. Tree Physiol. 2003, 23, 885–891. [Google Scholar]
- Li, Y.; Ye, S.; Hui, G.; Hu, Y.; Zhao, Z. Spatial structure of timber harvested according to structure-based forest management. For. Ecol. Manag. 2014, 322, 106–116. [Google Scholar] [CrossRef]
- Zhang, G.; Hui, G.; Zhang, G.; Zhao, Z.; Hu, Y. Telescope method for characterizing the spatial structure of a pine-oak mixed forest in the Xiaolong Mountains, China. Scand. J. For. Res. 2019, 34, 751–762. [Google Scholar] [CrossRef]
- Howard, J.L.; Gagne, M.; Morin, A.J.S.; Forest, J. Using Bifactor Exploratory Structural Equation Modeling to Test for a Continuum Structure of Motivation. Management 2016, 44, 2638–2664. [Google Scholar] [CrossRef]
- Perry, D.A.; Lotan, J.E. A framework for the development of forest spatial patterns. For. Sci. 1975, 21, 370–380. [Google Scholar]
- Aguirre, O.; Hui, G.; von Gadow, K.; Jiménez, J. An analysis of spatial forest structure using neighbourhood-based variables. For. Ecol. Manag. 2003, 183, 137–145. [Google Scholar] [CrossRef]
- Hui, G.Y.; Hu, Y.B. Measuring species spatial isolation in mixed forests. For. Res. 2001, 14, 23–27. [Google Scholar]
- Hui, G.Y.; Zhao, Z.H.; Hu, Y.B.; Zhang, G.Q.; Cheng, S.P.; Xu, X.F. Comprehensive evaluation of forest stand spatial structure based on the mean value of structural parameters. For. Res. 2023, 36, 12–21. [Google Scholar] [CrossRef]
- Hui, G.Y.; Gadow, K.V.; Zhao, Z.H.; Hu, Y.B.; Xu, H.; Li, Y.F.; Zhang, L.J.; Zhang, G.Q.; Liu, W.Z.; Yuan, S.Y. Principles of Structure-Based Forest Management; China Forestry Press: Beijing, China, 2016; Volume 6. [Google Scholar]
- Zhu, J.; Liu, S.; Wu, F.; Liu, W. Spatial Structure and Allometric Growth of Major Tree Species in a Subtropical Evergreen Broad-Leaved Forest. PLoS ONE 2016, 11, e0155275. [Google Scholar]
- Qian, H.; Zhang, J.; Wu, Z.; Yin, L.; Li, H. Incorporating species composition into models for stand volume using spatial linear models. For. Ecol. Manag. 2014, 328, 38–44. [Google Scholar]
- Zhang, L.; Liang, H.; Xu, M. Spatial distributions and correlations of forest structure, composition, and soil nutrients along an altitudinal gradient in a subtropical evergreen broadleaved forest. For. Ecol. Manag. 2018, 418, 41–48. [Google Scholar]
- Fortin, M.J.; Dale, M. Spatial Analysis: A Guide for Ecologists; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Turner, W.; Rondinini, C.; Pettorelli, N.; Mora, B.; Leidner, A.K.; Szantoi, Z.; Chauvenet, A. Free and open-access satellite data are key to biodiversity conservation. Biol. Conserv. 2015, 182, 173–176. [Google Scholar] [CrossRef]
- Levin, N.; Lechner, A.M.; Brown, G.; Mörtberg, U.M.; Mörtberg, M.; Lechner, A.M. A review of airborne LiDAR applications for 3D modelling and analysis of forests. Remote Sens. 2017, 9, 1070. [Google Scholar]
- He, H.S.; Shang, B.Z.; Crow, T.R.; Gustafson, E.J.; Shifley, S.R.; Fraser, J.S. Simulating forest fuel and fire risk dynamics across landscapes—LANDIS fuel module design. Ecol. Model. 2007, 201, 675–690. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A.; Naves, J. Using process-based models to simulate tree population dynamics in monodominant tropical forests. Ecology 1999, 80, 1214–1224. [Google Scholar]
- Pélissier, R.; Goreaud, F. A practical approach to the study of spatial structure in simple cases of heterogeneous vegetation. J. Veg. Sci. 2001, 12, 99–108. [Google Scholar] [CrossRef]
- Li, Y.; Lei, X.; Ma, S. Analysis of forest structure using remote sensing data: A review. For. Ecol. Manag. 2020, 458, 117762. [Google Scholar]
- Pommerenig, J.; Stoyan, D. Optimizing spatial forest structure under habitat constraints. Environ. Ecol. Stat. 2008, 15, 69–82. [Google Scholar]
- Packard, K.C.; Radosevich, S.R. Relating vegetation and environmental gradients through quantile regression. For. Ecol. Manag. 2001, 154, 347–359. [Google Scholar]
- Perry, D.A.; Amaranthus, M.P. The role of spatial processes in canopy dynamics. Can. J. For. Res. 2006, 36, 243–252. [Google Scholar]
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of global forest area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Kobe, R.K. Sapling growth as a function of light and landscape-level variation in soil water and foliar nitrogen in northern Michigan. Oecologia 2006, 147, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Canham, C.D.; Finzi, A.C. Disentangling changes in the spectral reflectance of forest canopy and understory due to natural and anthropogenic effects. Remote Sens. Environ. 2013, 132, 51–57. [Google Scholar]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef]
- Mladenoff, D.J.; Stearns, F. Eastern hemlock regeneration and deer browsing in the northern great lakes region: A re-examination and model simulation. Conserv. Biol. 1993, 7, 889–900. [Google Scholar] [CrossRef]
- Canham, C.D.; Finzi, A.C.; Pacala, S.W.; Burbank, D.H. Causes and consequences of resource heterogeneity in forests: Interspecific variation in light transmission by canopy trees. Can. J. For. Res. 1994, 24, 337–349. [Google Scholar] [CrossRef]
- Pacala, S.W.; Deutschman, D.H. Details that matter: The spatial distribution of individual trees maintains forest ecosystem function. Oikos 1995, 74, 357–365. [Google Scholar] [CrossRef]
- Pommerening, A.; Stoyan, D. Edge-correction needs in estimating indices of spatial forest structure. Can. J. For. Res. 2008, 38, 878–889. [Google Scholar] [CrossRef]
- Li, Y.; Weiskittel, A.R. Multivariate analysis of forest structure and composition: A systematic review and synthesis. For. Ecol. Manag. 2018, 427, 309–318. [Google Scholar]
- Gadow, K.V.; Hui, G.Y. Modeling Forest Development; Academic Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Li, H.; Lei, X.; Tang, J. Optimizing forest structure through spatially explicit simulation and multivariate analysis of forest attributes in Northeast China. For. Ecol. Manag. 2018, 424, 409–418. [Google Scholar]
- Pretzsch, H. Forest Dynamics, Growth, and Yield: From Measurement to Model; Springer Science Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Pukkala, T.; Kurttila, M.; Mehtätalo, L. A method for integrating stand growth models and harvest scheduling models. Eur. J. For. Res. 2009, 128, 1–11. [Google Scholar]
- Temesgen, H.; Monleon, V.J. Characterizing crown structure in old-growth and mature conifer stands using terrestrial lidar data and a structural clustering algorithm. Can. J. For. Res. 2015, 45, 199–208. [Google Scholar]
- Li, S.; Fang, X.; Chen, J.; Li, L.; Gu, X.; Liu, Z.; Zhang, S. Effects of different degrees of anthropogenic disturbance on biomass and spatial distribution in Subtropical forests in Central Southern China. Acta Ecol. Sin. 2018, 38, 6111–6124. [Google Scholar] [CrossRef]
- Spies, T.A.; Turner, W. Dynamic forest mosaics. In Forest Landscape Ecology: Transferring Knowledge to Practice; McDonnell, M.J., Pickett, S.T.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 155–171. [Google Scholar]
- Pretzsch, H.; Schütze, G. Transgressive overyielding in mixed compared with pure stands of Norway spruce and European beech in Central Europe: Evidence on stand level and explanation on individual tree level. Eur. J. For. Res. 2009, 128, 183–204. [Google Scholar] [CrossRef]
- Liang, J.; Buongiorno, J.; Monserud, R.A.; Kruger, E.L. Effects of diversity of tree species and size on forest basal area growth, recruitment, and mortality. For. Ecol. Manag. 2007, 243, 116–127. [Google Scholar] [CrossRef]
- Reed, D.D.; Jones, D.A.; Schuler, T.M. Economic return on long-term forest productivity: A synthesis of results from 35 years of experimental management on 50 northern hardwood stands. For. Sci. 2009, 55, 93–107. [Google Scholar]
- Buongiorno, J.; Peyron, J.L.; Houllier, F.; Bruciamacchie, M. A stand-growth model for even-aged stands of maritime pine (Pinus pinaster Ait.) in Mediterranean areas of France. For. Ecol. Manag. 2010, 259, 234–248. [Google Scholar]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Aubinet, M.; Chermanne, B.; Vandenhaute, M.; Longdoz, B.; Yernaux, M.; Laitat, E. Long term carbon dioxide exchange above a mixed forest in the Belgian Ardennes. Agric. For. Meteorol. 2001, 108, 293–315. [Google Scholar] [CrossRef]
- Vepakomma, U.; Pukkala, T. Optimal management of teak plantations in India: A spatial approach. For. Ecol. Manag. 2016, 360, 132–141. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Number | Slope Aspect | Slope/Degree | Tree Number | Mean DBH /cm | Mean Height /m | Mean East–West Crown Diameter/m | Mean North–South Crown Diameter/m |
|---|---|---|---|---|---|---|---|
| 1 | southwestern | 15 | 94 | 16.5 | 14.9 | 3.9 | 3.6 |
| 2 | south | 15 | 121 | 15.4 | 13.8 | 3.9 | 3.7 |
| 3 | south | 40 | 116 | 13.5 | 12.4 | 3.6 | 3.7 |
| 4 | southwestern | 20 | 125 | 12.9 | 10.1 | 2.1 | 2.6 |
| 5 | south | 14 | 98 | 13.5 | 11.8 | 2.5 | 2.8 |
| 6 | south | 20 | 126 | 9.6 | 8.5 | 3.1 | 3.3 |
| 7 | southwestern | 16 | 122 | 14.7 | 13.9 | 2.6 | 2.9 |
| 8 | southwestern | 15 | 95 | 10.7 | 9.1 | 2.5 | 2.4 |
| 9 | south | 20 | 131 | 13.1 | 12.7 | 3.3 | 3.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, J.; Cao, X.; Liu, Z.; Lv, Y. The Multivariate Distribution of Stand Spatial Structure and Tree Size Indices Using Neighborhood-Based Variables in Coniferous and Broad Mixed Forest. Forests 2023, 14, 2228. https://doi.org/10.3390/f14112228
Wang Y, Li J, Cao X, Liu Z, Lv Y. The Multivariate Distribution of Stand Spatial Structure and Tree Size Indices Using Neighborhood-Based Variables in Coniferous and Broad Mixed Forest. Forests. 2023; 14(11):2228. https://doi.org/10.3390/f14112228
Chicago/Turabian StyleWang, Yiru, Jiping Li, Xiaoyu Cao, Zhaohua Liu, and Yong Lv. 2023. "The Multivariate Distribution of Stand Spatial Structure and Tree Size Indices Using Neighborhood-Based Variables in Coniferous and Broad Mixed Forest" Forests 14, no. 11: 2228. https://doi.org/10.3390/f14112228