
Estimation of the Population Dynamics of Taxus cuspidata by Using a Static Life Table for Its Conservation
Abstract
:1. Introduction
2. Materials and Methods
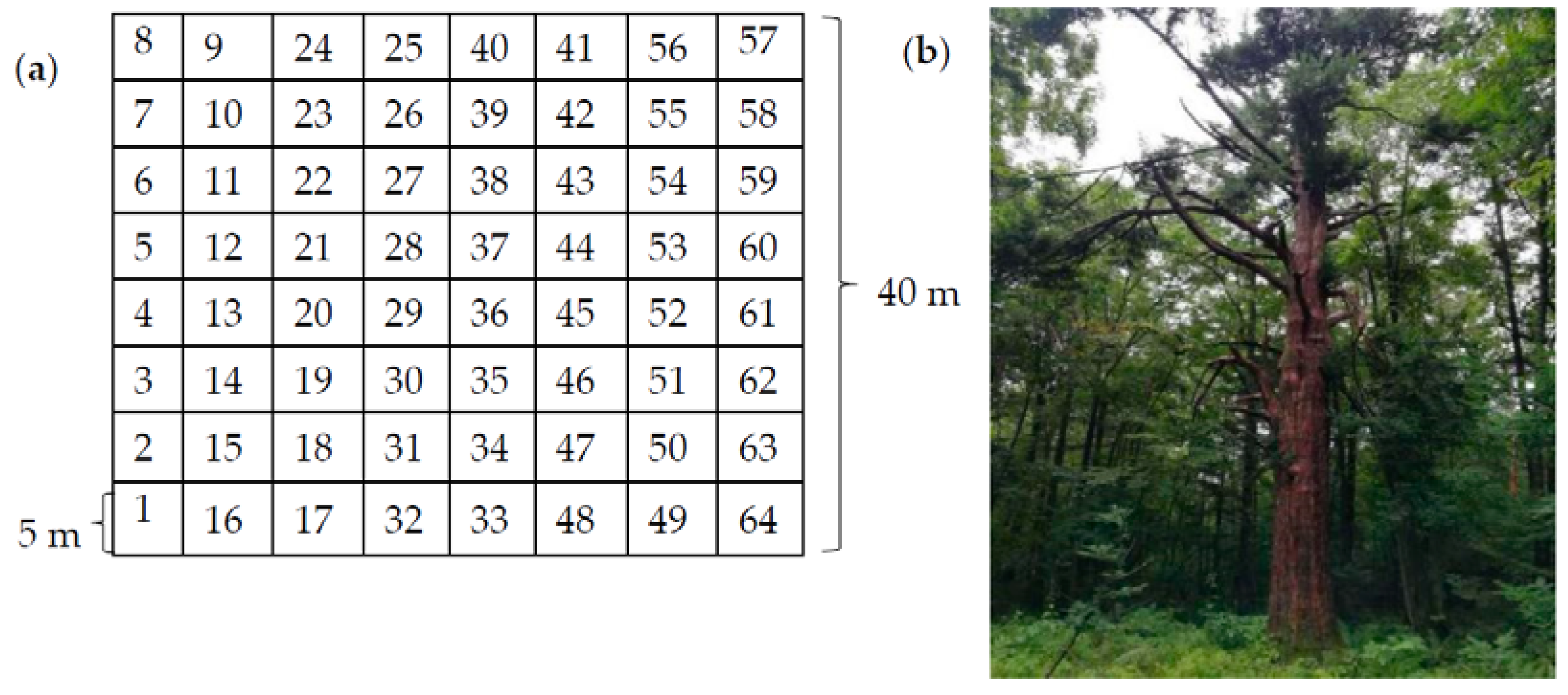
2.1. Sample Site Survey
2.2. Diameter Classification
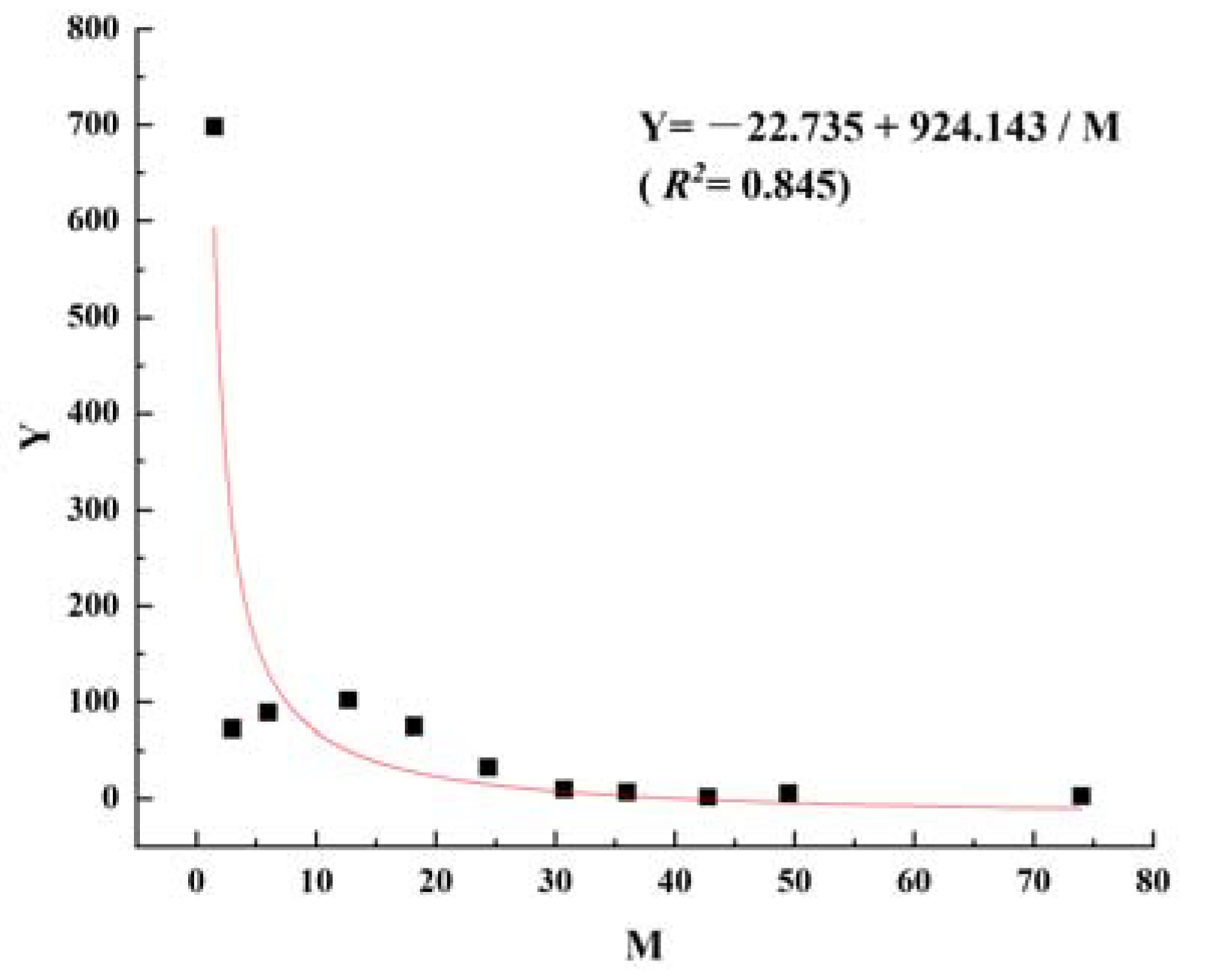
2.3. Smoothing Curve
2.4. Static Life Table Compilation
2.5. Survival Curve Drawing
2.6. Dynamic Analysis of Population Structure
2.7. Time Series Forecasting
3. Results
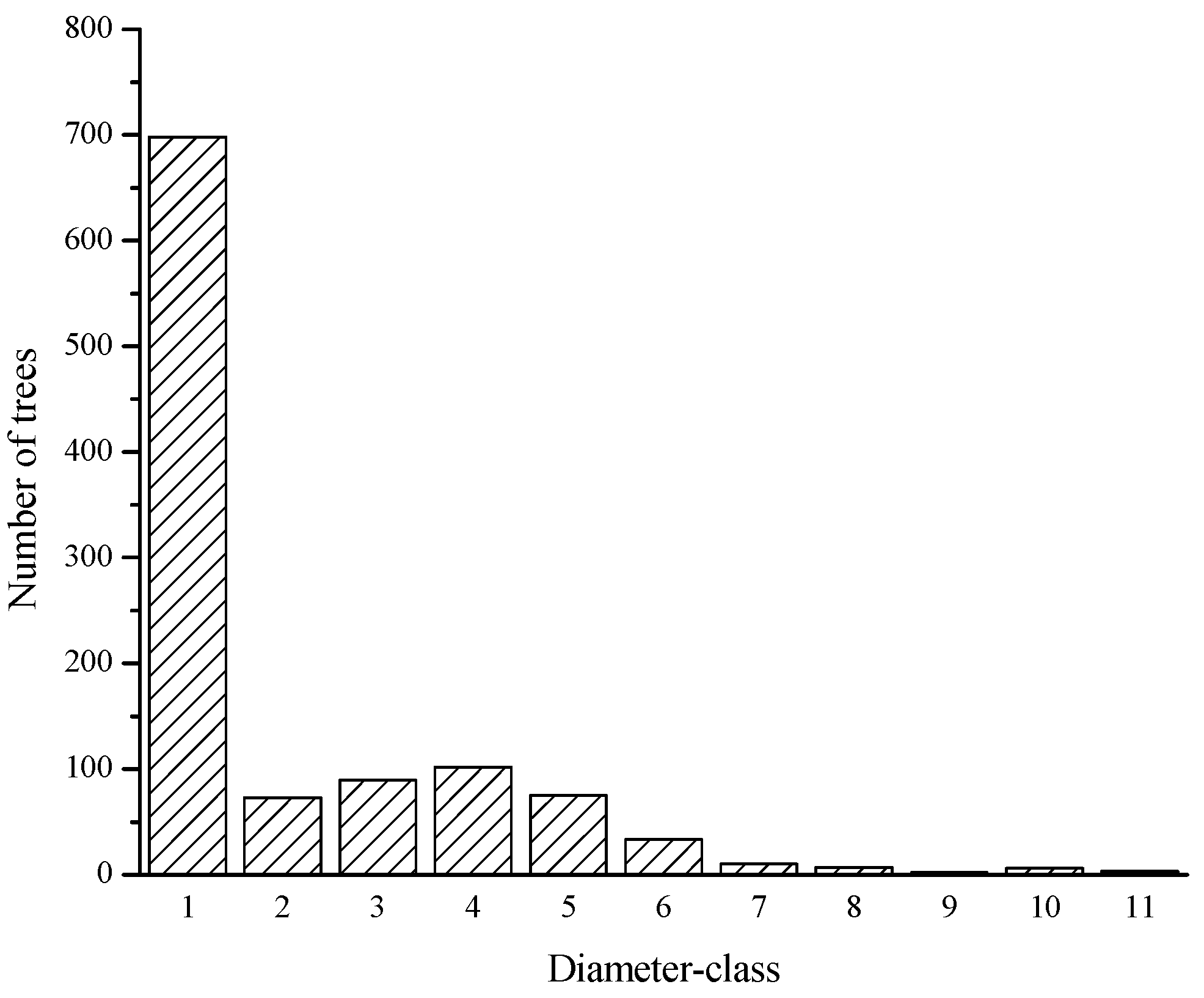
3.1. Population Size Structure of Taxus cuspidata
3.2. The Static Life Table of Taxus cuspidata
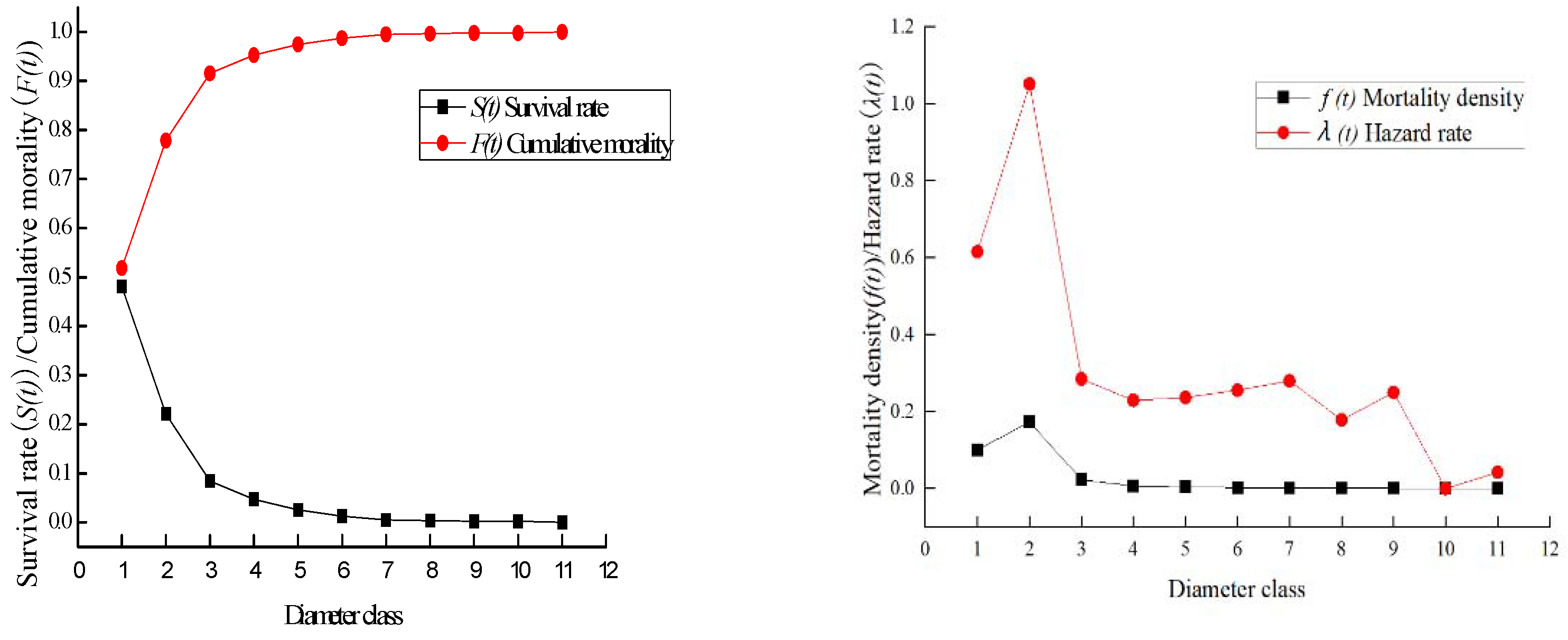
3.3. Survival Curve of Taxus cuspidata
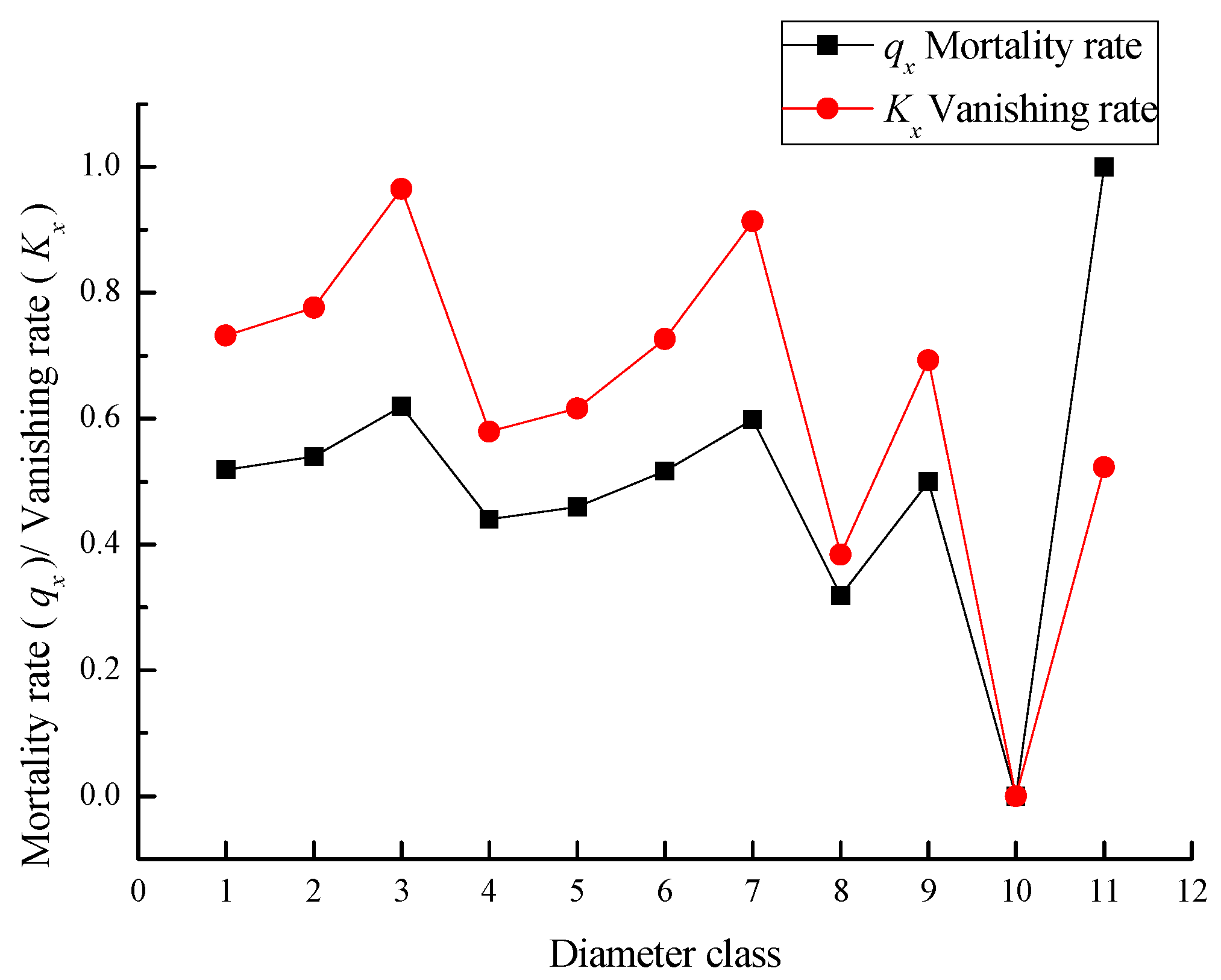
3.4. Population Survival Analysis of Taxus cuspidata
3.5. Dynamic Analysis of Population Structure of Taxus cuspidata
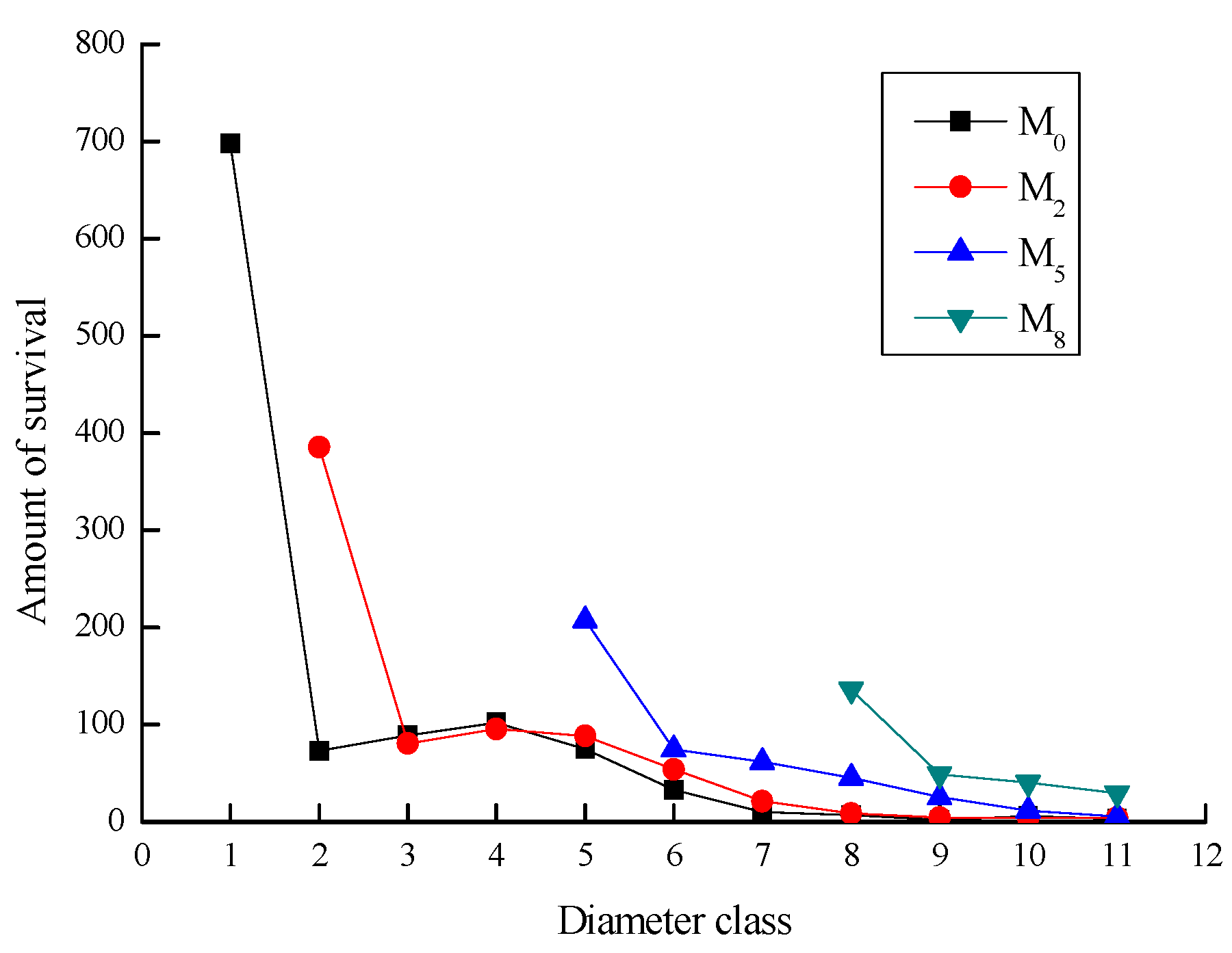
3.6. Dynamic Prediction of Taxus cuspidata Population
4. Discussion
4.1. Population Structure Analysis
4.2. Population Dynamics Analysis and Conservation Recommendations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zang, R.G.; Cheng, K.W.; Li, J.Q.; Zhang, W.Y.; Chen, X.F.; Tao, J.P. Natural Forest Biodiversity Conservation and Restoration; Science and Technology Press: Beijing, China, 2005; pp. 464–480. (In Chinese) [Google Scholar]
- Liu, T. Population Ecology of Natural Japanese Yew; Northeast Forestry University: Harebin, China, 2007; pp. 1–2, (In Chinese with English abstract). [Google Scholar]
- Chen, J.; Long, T.; Yang, L.; Wang, Y.; Xu, C.; Li, J.W. Habitat suitability assessment of Taxus cuspidata. J. Beijing For. Univ. 2019, 41, 51–59, (In Chinese with English abstract). [Google Scholar]
- Menges, E.S. Population viability analyses in plants: Challenges and opportunities. Trends Ecol. Evol. 2000, 15, 51–56. [Google Scholar] [CrossRef]
- Dang, H.; Zhang, Y.; Zhang, K.; Jiang, M.; Zhang, Q. Age structure and regeneration of subalpine fir (Abies fargesii) forests across an altitudinal range in the Qinling Mountains, China. For. Ecol. Manag. 2010, 259, 547–554. [Google Scholar] [CrossRef]
- Dolanc, C.R.; Thorne, J.H.; Safford, H.D. Widespread shifts in the demographic structure of subalpine forests in the Sierra Nevada, California, 1934 to 2007. Glob. Ecol. Biogeogr. 2013, 22, 264–276. [Google Scholar] [CrossRef]
- Diao, Y.F.; Jin, G.Z.; Tian, S.Y.; Liu, Y.K.; Liu, Y.L.; Han, L.D.; Li, Y.H. Species composition and community structure of a Taxus cuspidata forest in Muling nature reserve of Heilongjiang province, China. Sci. Silvae Sin. 2016, 52, 26–36, (In Chinese with English abstract). [Google Scholar]
- Easterling, M.R.; Ellner, S.P.; Dixon, P. Size specific sensitivity: A new structured population model. Ecology 2000, 81, 694–708. [Google Scholar] [CrossRef]
- Ramula, S.; Kerr, N.Z.; Crone, E.E. Using statistics to design and estimate vital rates inmatrix population models for a perennial herb. Popul. Ecol. 2019, 62, 53–63. [Google Scholar] [CrossRef]
- Werner, P.A.; Peacock, S.J. Savanna canopy trees under fire: Long-term persistence and transient dynamics from a stage-based matrix population mode. Ecosphere 2019, 10, e02706. [Google Scholar] [CrossRef]
- Gao, W.Q.; Ni, Y.Y.; Xue, Z.M.; Wand, X.F.; Kang, F.F.; Hu, J.; Gao, Z.H.; Jiang, Z.P.; Liu, J.F. Population structure and regeneration dynamics of Quercus variabilis along latitudinal and longitudinal gradients. Ecosphere 2017, 8, 1–15, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Jin, H.; Zhao, Y.; Yin, H.; Qin, L.W.; Liu, L.J.; Wang, C.; Jia, X.; Li, B.Y. Quantitative characteristics and dynamic analysis of the endangered species Rhododendron chrysanthum population in Changbai Mountain. Chin. J. Ecol. 2017, 36, 3123–3130, (In Chinese with English abstract). [Google Scholar]
- Kroon, H.D.; Groenendael, J.V.; Ehrlen, J. Elasticities: A review of methods and model limitations. Ecology 2000, 81, 607–618. [Google Scholar] [CrossRef]
- Wang, D.D.; Zhang, Y.W. Identification and genetic diversity analysis of Taxus cuspidata hybrid. J. Northeast Norm. Univ. 2019, 51, 113–118, (In Chinese with English abstract). [Google Scholar]
- Wu, S.X.; Liu, Y.H.; Zhang, L.M.; Shang, F.Q.; Tan, C.Q. Growth stability analysis of ex situ conservation of Taxus cuspidata seedling from different sources. J. Northeast Norm. Univ. 2018, 40, 27–37, (In Chinese with English abstract). [Google Scholar]
- Wang, T. Study on Taxus cuspidata var Seeding Raising Technology; Northeast Normal University: Harebin, China, 2017; pp. 11–22, (In Chinese with English abstract). [Google Scholar]
- Liu, T.; Xu, H.Y.; Jin, H.Y.; Zhang, Y.H.; Zhou, Z.Q. Diurnal changes of chlorphyll fluorescence characteristics of Japanese Yew seedlings under different light conditions. Bull. Bot. Res. 2015, 35, 848–853, (In Chinese with English abstract). [Google Scholar]
- Liu, T.; Jin, H.Y.; Xu, H.Y.; Zhou, Z.Q.; Zhang, Y.H. Diurnal changes od photosynthetic characteristics of Taxus cuspidata seedlings under different light conditions. J. Northeast Norm. Univ. 2015, 37, 67–72, (In Chinese with English abstract). [Google Scholar]
- Li, Y.L. Interspecific Relationship of the Taxus cuspidata; Northeast Forestry University: Harebin, China, 2008; pp. 17–35, (In Chinese with English abstract). [Google Scholar]
- Diao, Y.F.; Liu, Y.K.; Liu, Y.L.; Tian, S.Y. Population structure characteristics of Taxus cuspidata. Sci. Sin. Vitae 2020, 50, 391–397, (In Chinese with English abstract). [Google Scholar]
- Caswell, H. Matrix Population Models: Construction, Analysis and Interpretation; Sinauer: Sunderland, MA, USA, 1989. [Google Scholar]
- Caswell, H. Life cycle models for plants. Lect. Math. Life Sci. 1986, 18, 171–233. [Google Scholar]
- Vandermeer, J. Choosing category size in a stage projection matrix. Oecologia 1978, 32, 79–84. [Google Scholar] [CrossRef]
- Caswell, H. Prospective and retrospective perturbation analyses: Their roles in conservation biology. Ecology 2000, 81, 619–627. [Google Scholar] [CrossRef]
- Dong, M.Z.; Wang, L.T.; Lv, M.J.; Meng, D.; Yang, C.Y.; Zhao, C.J.; Fu, Y.J.; Yang, Q. Communities and protection strategy of endangered species wild Taxus cuspidata. Bull. Bot. Res. 2020, 40, 416–423, (In Chinese with English abstract). [Google Scholar]
- Zhang, X.S. Vegetation Map of China and Its Geographic Pattern: Illustration of the Vegetation Map of the People ’s Republic of China (1:1 000 000); Geological Publishing House: Beijing, China, 2007. (In Chinese) [Google Scholar]
- Bridie, C.; Fortin, H.M.J. Development of a Populus Balsamifera Clone in subarctic Quebec reconstructed from spatial analyses. J. Ecol. 1995, 83, 309–320. [Google Scholar] [CrossRef]
- Johnson, J.B. Stand structure and vegetation dynamics of a subalpine treed fen in Rocky Mountain National Park, Colorado. J. Veg. Sci. 1997, 8, 337–342. [Google Scholar] [CrossRef]
- Rebertus, A.J.; Veblen, T.T. Structure and tree fall gap dynamics of old-growth Nothofagus forests in Tierra del Fuego, Argentina. J. Veg. Sci. 1993, 4, 641–654. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Frost, I.; Rydin, H. Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce- dominated forests. Ecoscience 2000, 7, 38–44. [Google Scholar] [CrossRef]
- Harcombe, P.A.; Marks, P.L. Tree diameter distributions and replacement processes and replacement processes in southeast Texas forests. For. Sci. 1978, 24, 153–166. [Google Scholar]
- Lorimer, C.G. Age structure and disturbance history of a southern Appalachian virgin forest. Ecology 1980, 61, 1169–1184. [Google Scholar] [CrossRef]
- Gao, H.Z.; Huang, X.; Su, H.; Qiao, P.F.; Jiang, Z.M.; Shen, Y.R.; Cai, J. Structure and dynamic characteristics of Betula albo-sinensis populations in teo regions in the Qinling Mountains of northwestern China. J. Beijing For. Univ. 2022, 44, 12–20, (In Chinese with English abstract). [Google Scholar]
- Zhang, J.; Pan, Z.L.; Tian, Y.H.; Hou, J.L.; Peng, M.C.; Duan, H.; Li, Y.F.; Li, Y.P. Age structure and dynamics of Pinus yunnanensis population in Yunlong Tianchi Nature Reserve. Acta Ecol. Sin. 2022, 42, 9091–9099, (In Chinese with English abstract). [Google Scholar]
- LI, F.R. Study on diameter distribution and models prediction yields for natural Dahurian larch stands. J. North-East For. Univ. 1987, 15, 8–16, (In Chinese with English abstract). [Google Scholar]
- Ding, G.M.; Liu, X.M.; Ni, Z.Y.; Peng, J.Y. A preliminary study of population dynamics of Picea crassifolia in Qilian Mountains Gansu Province. J. Plant Resour. Environ. 2005, 14, 36–41, (In Chinese with English abstract). [Google Scholar]
- Begon, M.; Mortimer, M.; Thompson, D.J. Population Ecology: A Unified Study of Animal and Plants; Blackwell Science Ltd.: London, UK, 1981; pp. 1–1088. [Google Scholar]
- Jiang, H. Population Ecology of Spruce; Chinese Forestry Press: Beijing, China, 1992; pp. 1–26. (In Chinese) [Google Scholar]
- Liu, J.H.; Hong, W. A study on forecast of population dynamics of Castanopsis kawakamii. Chin J. Appl. Environ. Biol. 1999, 5, 247–253, (In Chinese with English abstract). [Google Scholar]
- Wratten, S.D.; Fry, G.I.A. Field and Laboratory Exercises in Ecology; Edward Arnad: London, UK, 1996; pp. 98–103. [Google Scholar]
- Wang, M. The Study on Population Dynamics and Spatial Distribution Pattern of Main Plants in Xiaolingshan of Gansu Province; Northwest Normal University: Lanzhou, Gansu, 2012; pp. 7–10, (In Chinese with English abstract). [Google Scholar]
- Yang, F.X.; Wang, S.Y.; Xu, H.G.; Li, B.Q. The theory of survival analysis and its application to life table. Acta Ecol. Sin. 1991, 11, 153–158, (In Chinese with English abstract). [Google Scholar]
- Deevey, E.S. Life tables for natural populations of animals. Q. Rev. Biol. 1947, 22, 283–314. [Google Scholar] [CrossRef] [PubMed]
- Hett, J.M.; Loucks, O.L. Age structure models of balsam fir and eastern hemlock. J. Ecol. 1976, 64, 1029–1044. [Google Scholar] [CrossRef]
- Chen, X.D. A study on the method of quantitative analysis for plant population and community structural dynamics. Acta Ecol. Sin. 1998, 18, 104–107, (In Chinese with English abstract). [Google Scholar]
- Liu, G.J.; Zhang, X.M.; Zhu, J.T.; Lv, C.Y.; Lu, Y. Structure and dynamics of Haloxy ammodendron population along the southeastern margin of the Zhunger basin. Acta Bot. Boreal.-Occident. Sin. 2011, 31, 1250–1256, (In Chinese with English abstract). [Google Scholar]
- Yang, L.; Zhang, Z.L.; Yun, Y.; Yan, W.P.; Chen, X.; Zhang, L.; Zheng, D.J.; Chen, J.L. The population structure and dynamics of Dracaena cambodiana, en endangered tree on Hainan island. Acta Ecol. Sin. 2018, 38, 201–214, (In Chinese with English abstract). [Google Scholar]
- Colling, G.; Matthies, D. Effects of habitat deterioration on population dynamics and extinction risk of an endangered, long-lived perennial herb (Scorzonera humilis). J. Ecol. 2006, 94, 959–972. [Google Scholar] [CrossRef]
- Su, J.Y.; Yan, Y.; Li, C.; Li, D.; Du, F. Informing conservation strategies with genetic diversity in wild plant with extremely small populations: A review on gymnosperms. Biodivers. Sci. 2020, 28, 376–384, (In Chinese with English abstract). [Google Scholar]
- Aguilar, R.; Quesada, M.; Ashworth, L.; Herrera’s-Diego, Y.; Lobo, J. Genetic consequences of habitat fragmentation in plant populations: Susceptible signals in plant traits and met-hodological approaches. Mol. Ecol. 2008, 17, 5177–5188. [Google Scholar] [CrossRef]
- Vranckx, G.U.Y.; Jacquemyn, H.; Muys, B.; Honnay, O. Meta-analysis of susceptibility of woody plants to loss of genetic diversity through habitat fragmentation. Conserv. Biol. 2012, 26, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.T.; Ison, J.L.; Ashley, M.V.; Howe, H.F. The paradox of forest fragmentation genetics. Conserv. Biol. 2008, 22, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Ellstrand, N.C.; Elam, D.R. Population genetic conseque-nces of small population size: Implications for plant conservation. Annu. Rev. Ecol. Syst. 1993, 24, 217–242. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Shen, W.H.; Tan, C.Q.; Xiang, W.H.; Peng, Y.H.; Pan, G.B. The population structure and dynamics analysis and protection strategy of Vatica guangxiensis extremely small population. Ecol. Sci. 2016, 35, 67–72. [Google Scholar]
- Cogoni, D.; Sulis, E.; Bacchetta, G.; Fenu, G. The unpredictable fate of the single population of a threatened narrow endemic Mediterranean plant. Biodivers. Conserv. 2019, 28, 1799–1813. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Elevation (m) | Inclination | Location | Latitude and Longitude | Dominant Tree | Population Density (/ha) | |
|---|---|---|---|---|---|---|---|
| N | E | ||||||
| 1 | 866 | 26° | Huanggou, Helong | 42°24′23″ | 128°38′4″ | Abies nephrolepis, Picea jezoensis, Tilia amurensis, Pinus koraiensis | 2.5 |
| 2 | 813 | 30° | Huanggou, Helong | 42°24′24.9″ | 128°40′5.9″ | Tilia amurensis, Acer pseudosieboldianum, Abies nephrolepis, Acer ukurunduense | 1.94 |
| 3 | 1021 | 30° | Huanggou, Helong | 42°24′27.9″ | 128°40′7.8″ | Acer ukurunduense, Abies nephrolepis, Acer tegmentosum, Tilia amurensis | 1.25 |
| 4 | 998 | 29° | Huanggou, Helong | 42°24′28.5″ | 128°40′10.8″ | Acer ukurunduense, Abies nephrolepis, Pinus koraiensis, Tilia amurensis, Taxus cuspidata | 1.69 |
| 5 | 979 | 32° | Huanggou, Helong | 42°24′06.7″ | 128°39′54.0″ | Tilia amurensis, Acer pseudosieboldianum, Abies nephrolepis, Acer ukurunduense | 3.56 |
| 6 | 1130 | 12° | Huanggou, Helong | 42°23′58.8″ | 128°40′0.9″ | Pinus koraiensis, Acer ukurunduense, Abies nephrolepis, Tilia amurensis, Abies holophylla | 1.81 |
| 7 | 1030 | 23° | Huanggou, Helong | 42°23′45″ | 128°39′56″ | Acer ukurunduense, Pinus koraiensis, Abies nephrolepis, Acer pseudosieboldianum, Tilia amurensis | 3.38 |
| 8 | 873 | 13° | Jingouling, Wangqing | 43°22′29″ | 130°9′33.9″ | Abies nephrolepis, Taxus cuspidata, Tilia amurensis, Pinus koraiensis, Betula costata | 17.56 |
| 9 | 694 | 17° | Jingouling, Wangqing | 43°21′37″ | 130°10′12.8″ | Abies nephrolepis, Acer pseudosieboldianum, Taxus cuspidata,, Acer mono, Pinus koraiensis | 6.38 |
| 10 | 910 | 14° | Duhuangzi, Wangqing | 43°12′24″ | 130°36′35.3″ | Abies nephrolepis, Acer ukurunduense, Tilia amurensis, Pinus koraiensis, Acer tegmentosum | 2.06 |
| 11 | 907 | 13° | Duhuangzi, Wangqing | 43°12′22.7″ | 130°36′35.7″ | Abies nephrolepis, Tilia amurensis, Acer tegmentosum, Acer barbinerve, Acer ukurunduense | 1.5 |
| 12 | 856 | 21° | Duhuangzi, Wangqing | 43°12′27.7″ | 130°36′36.7″ | Abies nephrolepis, Acer barbinerve, Pinus koraiensis, Tilia amurensis, Abies holophylla | 2.38 |
| 13 | 808 | 18° | Duhuangzi, Wangqing | 43°12′23.0″ | 130°36′25.8″ | Abies nephrolepis, Acer barbinerve, Tilia amurensis, Acer tegmentosum, Acer ukurunduense | 0.81 |
| 14 | 818 | 18° | Duhuangzi, Wangqing | 43°12′28.6″ | 130°36′25.3″ | Abies nephrolepis, Acer barbinerve, Tilia amurensis, Acer ukurunduense, Acer tegmentosum | 1.19 |
| 15 | 936 | 24° | Duhuangzi, Wangqing | 43°11′51.3″ | 130°36′25.7″ | Abies nephrolepis, Tilia amurensis, Acer barbinerve, Acer ukurunduense, Acer tegmentosum | 1.75 |
| 16 | 900 | 3° | Huanggou, Wangqing | 43°18′45.8″ | 130°19′31″ | Abies nephrolepis, Acer barbinerve, Acer ukurunduense, Tilia amurensis, Taxus cuspidata | 4.94 |
| 17 | 700 | 12° | Lanjia, Wangqing | 43°26′19″ | 130°57′25″ | Acer barbinerve, Abies nephrolepis, Pinus koraiensis, Acer ukurunduense, Acer tegmentosum | 4 |
| 18 | 789 | 7° | Lanjia, Wangqing | 43°26′19.4″ | 130°57′15.8″ | Acer barbinerve, Tilia amurensis, Abies nephrolepis, Acer ukurunduense, Pinus koraiensis | 0.75 |
| 19 | 766 | 27° | Lanjia, Wangqing | 43°27′11.6″ | 130°56′12.4″ | Abies nephrolepis, Tilia amurensis, Acer tegmentosum, Acer barbinerve, Abies holophylla | 3.25 |
| 20 | 745 | 20° | Madida, Wangqing | 43°9″35.5′ | 130°41′23.5″ | Abies nephrolepis, Acer tegmentosum, Tilia amurensis, Acer ukurunduense, Abies holophylla | 2.31 |
| Breast Diameter (cm) | 0–2.5 | 2.5–4 | 4–10 | 10–16 | 16–22 | 22–28 | 28–34 | 34–40 | 40–46 | 46–52 | 52+ |
| Diameter class | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| x | a0 | ax | M | lx | lnlx | dx | qx | Lx | Tx | ex | Kx |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 698 | 593 | 1.5 | 1000 | 6.9078 | 519 | 0.5189 | 741 | 741 | 0.74 | 0.7316 |
| 2 | 73 | 285 | 3 | 481 | 6.1761 | 260 | 0.5398 | 351 | 1092 | 2.27 | 0.7762 |
| 3 | 89 | 131 | 6 | 221 | 5.4000 | 137 | 0.6189 | 153 | 1245 | 5.62 | 0.9647 |
| 4 | 102 | 50 | 12.7 | 84 | 4.4352 | 37 | 0.4395 | 66 | 1311 | 15.53 | 0.5790 |
| 5 | 75 | 28 | 18.2 | 47 | 3.8563 | 22 | 0.4601 | 36 | 1347 | 28.48 | 0.6164 |
| 6 | 33 | 15 | 24.4 | 26 | 3.2399 | 13 | 0.5166 | 19 | 1366 | 53.50 | 0.7269 |
| 7 | 10 | 7 | 30.75 | 12 | 2.5130 | 7 | 0.5989 | 9 | 1375 | 111.38 | 0.9135 |
| 8 | 7 | 3 | 36 | 5 | 1.5995 | 2 | 0.3187 | 4 | 1379 | 278.50 | 0.3838 |
| 9 | 2 | 2 | 42.75 | 3 | 1.2157 | 2 | 0.5000 | 3 | 1381 | 409.53 | 0.6931 |
| 10 | 6 | 1 | 49.5 | 2 | 0.5226 | 0 | 0.0000 | 2 | 1383 | 820.07 | 0.0000 |
| 11 | 3 | 1 | 74 | 2 | 0.5226 | 2 | 1.0000 | 1 | 1384 | 820.57 | 0.5226 |
| Item | V1 | V2 | V3 | V4 | V5 | V6 | V7 | V8 | V9 | V10 | V11 | Vpi | V′pi |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dynamic index (%) | 0.9 | −0.18 | −0.13 | 0.26 | 0.56 | 0.7 | 0.3 | 0.71 | −0.67 | 0.5 | _ | 0.6395 | 0.0291 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Guo, Z.; Cui, X.; Fan, C. Estimation of the Population Dynamics of Taxus cuspidata by Using a Static Life Table for Its Conservation. Forests 2023, 14, 2194. https://doi.org/10.3390/f14112194
Liu D, Guo Z, Cui X, Fan C. Estimation of the Population Dynamics of Taxus cuspidata by Using a Static Life Table for Its Conservation. Forests. 2023; 14(11):2194. https://doi.org/10.3390/f14112194
Chicago/Turabian StyleLiu, Dan, Zhongling Guo, Xiaoyang Cui, and Chunnan Fan. 2023. "Estimation of the Population Dynamics of Taxus cuspidata by Using a Static Life Table for Its Conservation" Forests 14, no. 11: 2194. https://doi.org/10.3390/f14112194