
Effects of Distance and Neighbor Size on Abies hickelii: The Asymmetric Competition Is Aggravated in an Endangered Species
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
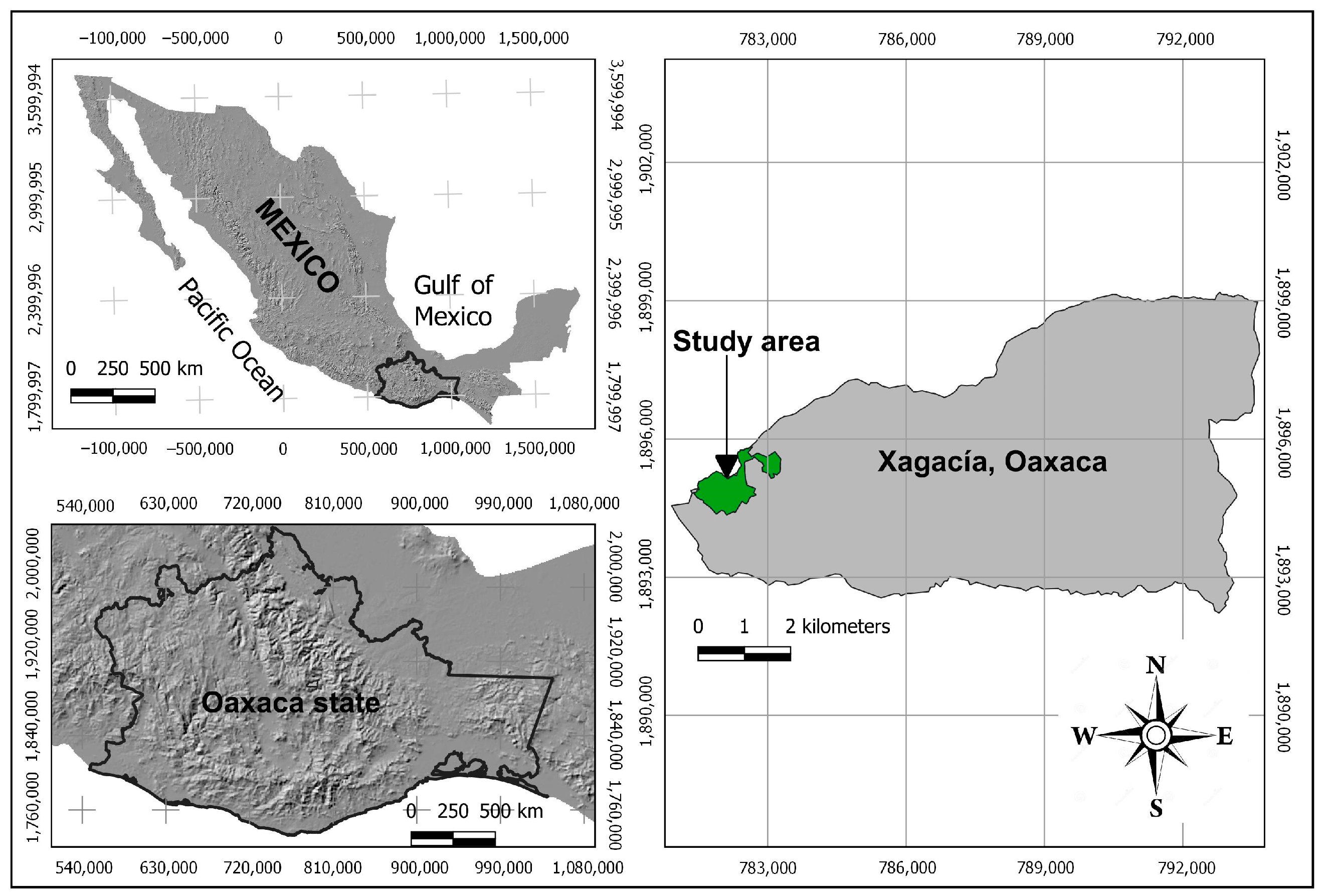
2.1. Sampling Area and Data Collection
2.2. Sampling and Data Collection
2.3. Data Analysis
3. Results
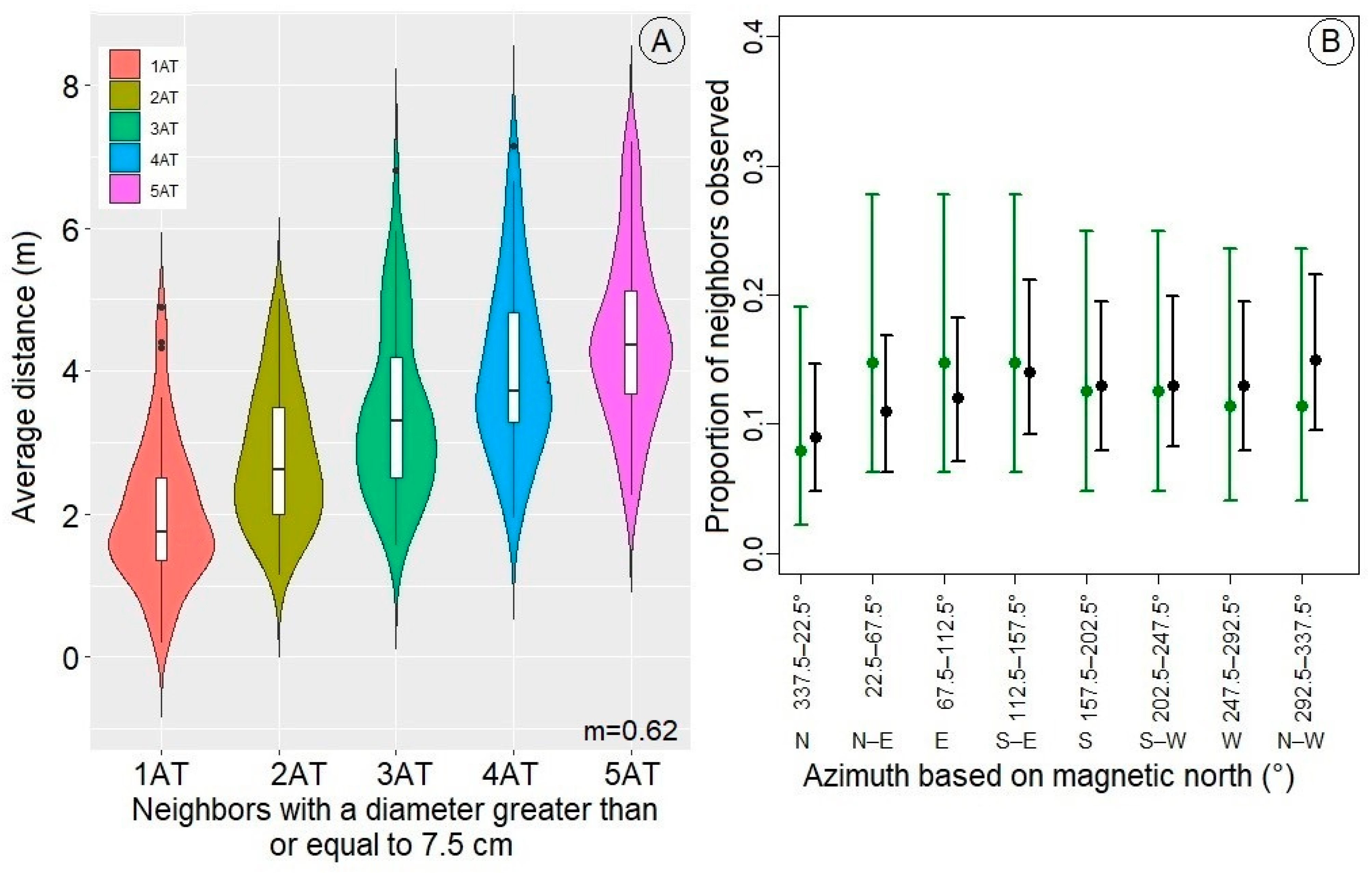
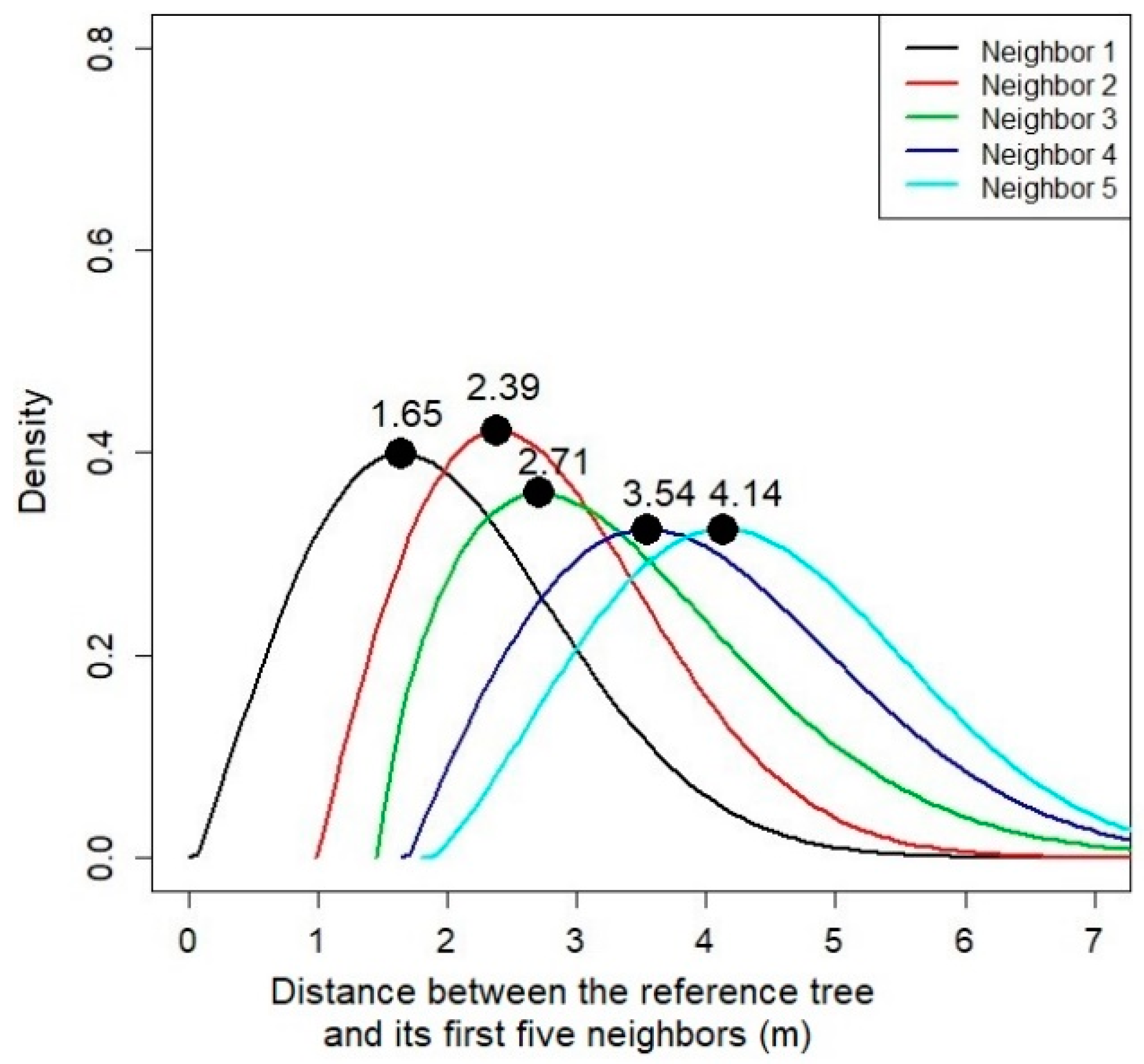
3.1. Distribution of the Neighbors and the Effect of Distance from the Neighbors
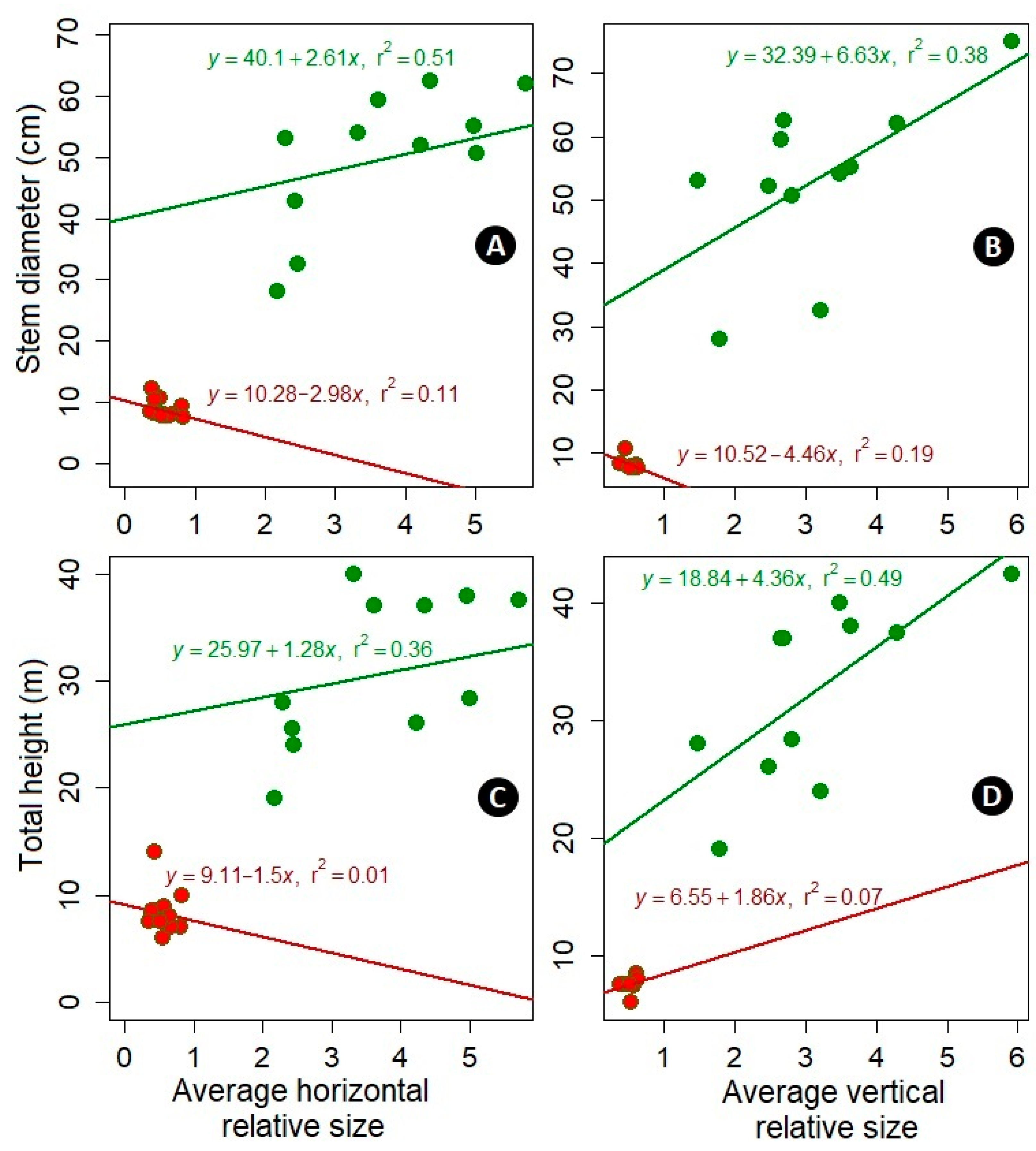
3.2. Effect of Neighbors Due to Their Sizes
3.3. Neighbor Effect Due to Size, Distance, and Species
4. Discussion
4.1. Evidence of Asymmetric Competition
4.2. Vertical Relative Size Has a Greater Effect Than the Horizontal Relative Size
4.3. The Roles of Species, Distance, and Size
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rehder, A. The firs of Mexico and Guatemala. J. Arnold Arbor. 1939, 20, 281–287. Available online: https://www.biodiversitylibrary.org/partpdf/21103 (accessed on 25 May 2023). [CrossRef]
- IUCN. The International Union for Conservation of Nature’s Red List of Threatened Species. 2023. Available online: https://www.iucnredlist.org/en (accessed on 27 June 2023).
- Farjon, A. Abies hickelii. The IUCN Red List of Threatened Species 2013, e.T42286A2969866. 2013. Available online: https://www.iucnredlist.org/species/42286/2969866 (accessed on 26 May 2023).
- Charlesworth, D.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Gu, L.; Zhang, D. Low genetic diversity and high genetic divergence caused by inbreeding and geographical isolation in the populations of endangered species Loropetalum subcordatum (Hamamelidaceae) endemic to China. Conserv. Genet. 2010, 11, 2281–2288. [Google Scholar] [CrossRef]
- Bello, C.H.Á.; Mata, L.L. Distribución y análisis estructural de Abies hickelii (flous & gaussen) en México. Interciencia 2001, 26, 244–251. [Google Scholar]
- Cogoni, D.; Sulis, E.; Bacchetta, G.; Fenu, G. The unpredictable fate of the single population of a threatened narrow endemic Mediterranean plant. Biodivers. Conserv. 2019, 28, 1799–1813. [Google Scholar] [CrossRef]
- Clark, P.J.; Evans, F.C. Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology 1954, 35, 445–453. [Google Scholar] [CrossRef]
- Weiner, J. Neighbourhood interference amongst Pinus rigida individuals. J. Ecol. 1984, 72, 183–195. [Google Scholar] [CrossRef]
- Goldberg, D.E. Neighborhood competition in an old-field plant community. Ecology 1987, 68, 1211–1223. [Google Scholar] [CrossRef]
- Stoll, P.; Weiner, J. A Neighborhood View of Interactions among Individual Plants. 2000. Available online: https://edoc.unibas.ch/15365/1/20180110105505_5a55e2f91b17e.pdf (accessed on 27 June 2023).
- Balandier, P.; Collet, C.; Miller, J.H.; Reynolds, P.E.; Zedaker, S.M. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 2006, 79, 3–27. [Google Scholar] [CrossRef]
- Mack, R.N.; Harper, J.L. Interference in dune annuals: Spatial pattern and neighbourhood effects. J. Ecol. 1977, 65, 345–363. [Google Scholar] [CrossRef]
- Biging, G.S.; Dobbertin, M.A. comparison of distance-dependent competition measures for height and basal area growth of individual conifer trees. For. Sci. 1992, 38, 695–720. [Google Scholar]
- Prodan, M. Punktstichprobe fur die forsteinrichtung [Point sampling for forest inventory]. Forst. Holzwirt. 1968, 23, 225–226. [Google Scholar]
- Ammann, P.; Waldbau, F. Erfolg der Jungwaldpflege im Schweizer Mittelland? Analyse und Folgerungen (Essay) [Is young growth tending successful in the Swiss plateau region? Analysis and implications (essay)]. Schweiz Z Forstwes 1969, 164, 262–270. Available online: https://ethz.ch/content/dam/ethz/special-interest/usys/ites/waldmgmt-waldbau-dam/documents/unterrichtshilfen/Management/Jungwaldpflege-Ammann-3013 (accessed on 25 July 2023). [CrossRef]
- Lynch, T.B.; Rusydi, R. Distance sampling for forest inventory in Indonesian teak plantations. For. Ecol. Manag. 1999, 113, 215–221. [Google Scholar] [CrossRef]
- Caeiro, F.; Mateus, A. Package ‘Randtests’. 2022. Available online: https://cran.r-project.org/web/packages/randtests/randtests.pdf (accessed on 27 June 2023).
- Mangiafico, S.S. An R Companion for the Handbook of Biological Statistics. 2015. Available online: https://rcompanion.org/documents/RCompanionBioStatistics.pdf (accessed on 27 April 2023).
- Clopper, C.; Pearson, E.S. “The use of confidence or fiducial limits illustrated in the case of the binomial”. Biometrika 1934, 26, 404–413. [Google Scholar] [CrossRef]
- Antúnez, P.; López Serrano, P.M.; González Adame, G.; Rubio Camacho, E.A.; Mendoza Díaz, M.M. Efecto de la altitud, pendiente y exposición geográfica en la distribución de helechos arborescentes. Acta Bot. Mex. 2022, 29, e1962. [Google Scholar] [CrossRef]
- Torchiano, M.; Torchiano, M.M. Package “Effsize”. 2020. Available online: https://cran.r-project.org/web/packages/effsize/effsize.pdf (accessed on 10 June 2023).
- Robinson, A.P.; Hamann, J.D. Forest Analytics with R: An Introduction; Springer Science & Business Media: New York, NY, USA, 2010. [Google Scholar]
- Prodan, M.; Peters, R.; Cox, F.; Real, P. Mensura Forestal; Agroamerica: San José, Costa Rica, 1997. [Google Scholar]
- Weiner, J. Asymmetric competition in plant populations. Trends Ecol. Evol. 1990, 5, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Connolly, J.; Wayne, P. Asymmetric Competition between Plant Species. Oecologia 1996, 108, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Freckleton, R.P.; Watkinson, A.R. Asymmetric competition between plant species. Funct. Ecol. 2001, 15, 615–623. [Google Scholar] [CrossRef]
- Drew, T.J.; Flewelling, J.W. Some recent Japanese theories of yield-density relationships and their application to Monterey pine plantations. For. Sci. 1977, 23, 517–534. [Google Scholar] [CrossRef]
- Trouvé, R.; Bontemps, J.D.; Seynave, I.; Collet, C.; Lebourgeois, F. Stand density, tree social status and water stress influence allocation in height and diameter growth of Quercus petraea (Liebl.). Tree Physiol. 2015, 35, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Duchesneau, R.; Lesage, I.; Messier, C.; Morin, H. Effects of light and intraspecific competition on growth and crown morphology of two size classes of understory balsam fir saplings. For. Ecol. Manag. 2001, 140, 215–225. [Google Scholar] [CrossRef]
- Pollastrini, M.; Holland, V.; Brüggemann, W.; Koricheva, J.; Jussila, I.; Scherer-Lorenzen, M.; Berger, S.; Bussotti, F. Interactions and competition processes among tree species in young experimental mixed forests, assessed with chlorophyll fluorescence and leaf morphology. Plant Biol. 2014, 16, 323–331. [Google Scholar] [CrossRef]
- Sevik, H.; Cetin, M.; Kapucu, O. Effect of light on young structures of Turkish fir (Abies nordmanniana subsp. bornmulleriana). Oxid. Commun. 2016, 39, 485–492. [Google Scholar]
- Drexhage, M.; Colin, F. Estimating root system biomass from breast-height diameters. Forestry 2001, 74, 491–497. [Google Scholar] [CrossRef]
- Zanetti, C.; Vennetier, M.; Mériaux, P.; Provansal, M. Plasticity of tree root system structure in contrasting soil materials and environmental conditions. Plant Soil 2015, 387, 21–35. [Google Scholar] [CrossRef]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Ann. Rev. Ecol. Syst. 1997, 28, 545–570. [Google Scholar] [CrossRef]
- Contreras, M.A.; Affleck, D.; Chung, W. Evaluating tree competition indices as predictors of basal area increment in western Montana forests. For. Ecol. Manag. 2011, 262, 1939–1949. [Google Scholar] [CrossRef]
- Forrester, D.I.; Kohnle, U.; Albrecht, A.T.; Bauhus, J. Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manag. 2013, 304, 233–242. [Google Scholar] [CrossRef]
- Schmitt, J. Reaction norms of morphological and life-history traits to light availability in Impatiens capensis. Evolution 1993, 47, 1654–1668. [Google Scholar] [CrossRef]
- Schmitt, J.; Wulff, R.D. Light spectral quality, phytochrome and plant competition. Trends Ecol. Evol. 1993, 8, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Antúnez, P. Influence of physiography, soil and climate on Taxus globosa. Nord. J. Bot. 2021, 39. [Google Scholar] [CrossRef]
- Antúnez, P.; Wehenkel, C.; Hernández-Díaz, J.C.; Garza-López, M. Quantile regression as a complementary tool for modelling biological data with high variability. J. Trop. For. Sci. 2023, 35, 130–140. Available online: https://www.jstor.org/stable/48723350 (accessed on 7 August 2023). [CrossRef]
- Martin, G.L.; Ek, A.R. A comparison of competition measures and growth models for predicting plantation red pine diameter and height growth. For. Sci. 1984, 30, 731–743. [Google Scholar] [CrossRef]
- Johann, K. Der A-Wert, ein objektiver Parameter zur Bestimmung der Freistellungsstärke von Zentralbäumen. In Tagungsbericht, Deutscher Verband Forstlicher Versuchsanstalten—Sektion Ertragskunde; [The A-value, an objective criterion for estimating the release of central trees from competition]; DVFFVA: Weibersbrunn, Germany, 1982; pp. 146–158. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DBH-AT | TH-AT | DBH-RT | TH-RT | |
|---|---|---|---|---|
| Minimum | 1.66 | 4.10 | 7.50 | 4.45 |
| Maximum | 119.60 | 46.00 | 75.05 | 42.50 |
| Mean | 24.66 | 16.73 | 23.27 | 16.74 |
| Standard deviation | 20.20 | 9.92 | 19.41 | 10.62 |
| Asymmetry coefficient | 1.49 | 1.00 | 1.10 | 1.09 |
| Non-Standardized Coefficients Associated with the Neighbor’s Distance | ||||||
|---|---|---|---|---|---|---|
| Variable Assessed | Grouping Criteria | Coefficients | SE | T-Value | p-Value | R2 |
| DBH | Ungrouped | −4.82 | 6.95 | −0.694 | 0.492 | 0.011 |
| DBH | DBH of the RT > any of the neighbors | −5.460 | 8.758 | −0.623 | 0.547 | 0.037 |
| DBH | DBH of the RT is < any of its neighbors | 1.132 | 0.464 | 2.441 | 0.031 | 0.332 |
| TH | Ungrouped | −3.394 | 3.838 | −0.884 | 0.381 | 0.018 |
| TH | TH of the RT > any of the neighbors | 0.357 | 6.963 | 0.051 | 0.960 | 0.0003 |
| TH | TH of the RT is < any of its neighbors | −1.218 | 0.386 | −3.153 | 0.020 | 0.624 |
| Standardized coefficients associated with the neighbor’s distance | ||||||
| DBH | Ungrouped | −0.200 | 0.288 | −0.694 | 0.492 | 0.011 |
| DBH | DBH of the RT > any of the neighbors | −0.227 | 0.364 | −0.623 | 0.547 | 0.037 |
| DBH | DBH of the RT is < any of its neighbors | 0.047 | 0.019 | 2.441 | 0.033 | 0.332 |
| TH | Ungrouped | −0.254 | 0.287 | −0.884 | 0.381 | 0.018 |
| TH | TH of the RT > any of the neighbors | 0.026 | 0.513 | 0.051 | 0.960 | 0.0003 |
| TH | TH of the RT is < any of its neighbors | −0.090 | 0.028 | −3.153 | 0.020 | 0.624 |
| Variable Assessed | Grouping Criteria | Coefficients | SE | T-Value | p-Value | R2 |
|---|---|---|---|---|---|---|
| Non-standardized coefficients associated with the horizontal relative size | ||||||
| DBH | Ungrouped | 5.909 | 0.648 | 8.906 | <0.001 | 0.648 |
| DBH | DBH of the RT > any of the neighbors | 2.611 | 0.808 | 3.233 | 0.009 | 0.511 |
| DBH | DBH of the RT is < any of its neighbors | −2.980 | 2.489 | −1.197 | 0.254 | 0.107 |
| TH | Ungrouped | 3.140 | 0.394 | 7.973 | <0.001 | 0.596 |
| TH | DBH of the RT > any of the neighbors | 1.275 | 0.540 | 2.360 | 0.040 | 0.358 |
| TH | DBH of the RT is < any of its neighbors | −1.497 | 3.503 | −0.427 | 0.676 | 0.015 |
| Non-standardized coefficients associated with the vertical relative size | ||||||
| DBH | Ungrouped | 14.131 | 1.074 | 13.157 | <0.001 | 0.801 |
| DBH | TH of the RT > any of the neighbors | 6.626 | 2.844 | 2.330 | 0.045 | 0.376 |
| DBH | TH of the RT is < any of its neighbors | −4.465 | 3.751 | −1.190 | 0.279 | 0.191 |
| TH | TH ungrouped | 8.002 | 0.539 | 14.836 | <0.001 | 0.837 |
| TH | TH of the RT > any of the neighbors | 4.362 | 1.490 | 2.928 | 0.017 | 0.488 |
| TH | TH of the RT < any of the neighbors | 1.859 | 2.816 | 0.660 | 0.534 | 0.068 |
| Standardized coefficients associated with the horizontal relative size | ||||||
| DBH | Ungrouped | 0.805 | 0.090 | 8.906 | <0.001 | 0.648 |
| DBH | DBH of the RT > any of the neighbors | 0.356 | 0.110 | 3.233 | 0.009 | 0.511 |
| DBH | DBH of the RT is < any of its neighbors | −0.406 | 0.339 | −1.197 | 0.254 | 0.107 |
| TH | Ungrouped | 0.773 | 0.097 | 7.973 | <0.001 | 0.596 |
| TH | DBH of the RT > any of the neighbors | 0.313 | 0.133 | 2.360 | 0.039 | 0.358 |
| TH | DBH of the RT is < any of its neighbors | −0.368 | 0.862 | −0.427 | 0.677 | 0.015 |
| Standardized coefficients associated with the vertical relative size | ||||||
| DBH | Ungrouped | 0.895 | 0.068 | 13.160 | <0.001 | 0.801 |
| DBH | TH of the RT > any of the neighbors | 6.626 | 2.844 | 2.330 | 0.044 | 0.376 |
| DBH | TH of the RT is < any of its neighbors | −4.465 | 3.751 | −1.190 | 0.278 | 0.191 |
| TH | TH ungrouped | 0.915 | 0.062 | 14.840 | <0.001 | 0.837 |
| TH | TH of the RT > any of the neighbors | 4.362 | 1.490 | 2.928 | 0.017 | 0.488 |
| TH | TH of the RT < any of the neighbors | 1.859 | 2.816 | 0.660 | 0.534 | 0.067 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antúnez, P.; Hernández-Cruz, I.; Ibrahim-Abdulsalam, F.; Clark-Tapia, R.; Ruiz-Aquino, F.; Valenzuela-Encinas, C. Effects of Distance and Neighbor Size on Abies hickelii: The Asymmetric Competition Is Aggravated in an Endangered Species. Forests 2023, 14, 1654. https://doi.org/10.3390/f14081654
Antúnez P, Hernández-Cruz I, Ibrahim-Abdulsalam F, Clark-Tapia R, Ruiz-Aquino F, Valenzuela-Encinas C. Effects of Distance and Neighbor Size on Abies hickelii: The Asymmetric Competition Is Aggravated in an Endangered Species. Forests. 2023; 14(8):1654. https://doi.org/10.3390/f14081654
Chicago/Turabian StyleAntúnez, Pablo, Iván Hernández-Cruz, Fatima Ibrahim-Abdulsalam, Ricardo Clark-Tapia, Faustino Ruiz-Aquino, and César Valenzuela-Encinas. 2023. "Effects of Distance and Neighbor Size on Abies hickelii: The Asymmetric Competition Is Aggravated in an Endangered Species" Forests 14, no. 8: 1654. https://doi.org/10.3390/f14081654