
Mountain Taiga in a Warming Climate: Contrast of Siberian Pine Growth along an Elevation Gradient
 , , , and
, , , and
Abstract
:1. Introduction
- How does SP growth respond to changes in thermal and moisture regimes along the elevation gradient?
- Does climate warming switch the limitation of trees’ growth by temperature to the limitation by moisture?
- Does the survival of SP differ within different elevation belts?
2. Materials and Methods
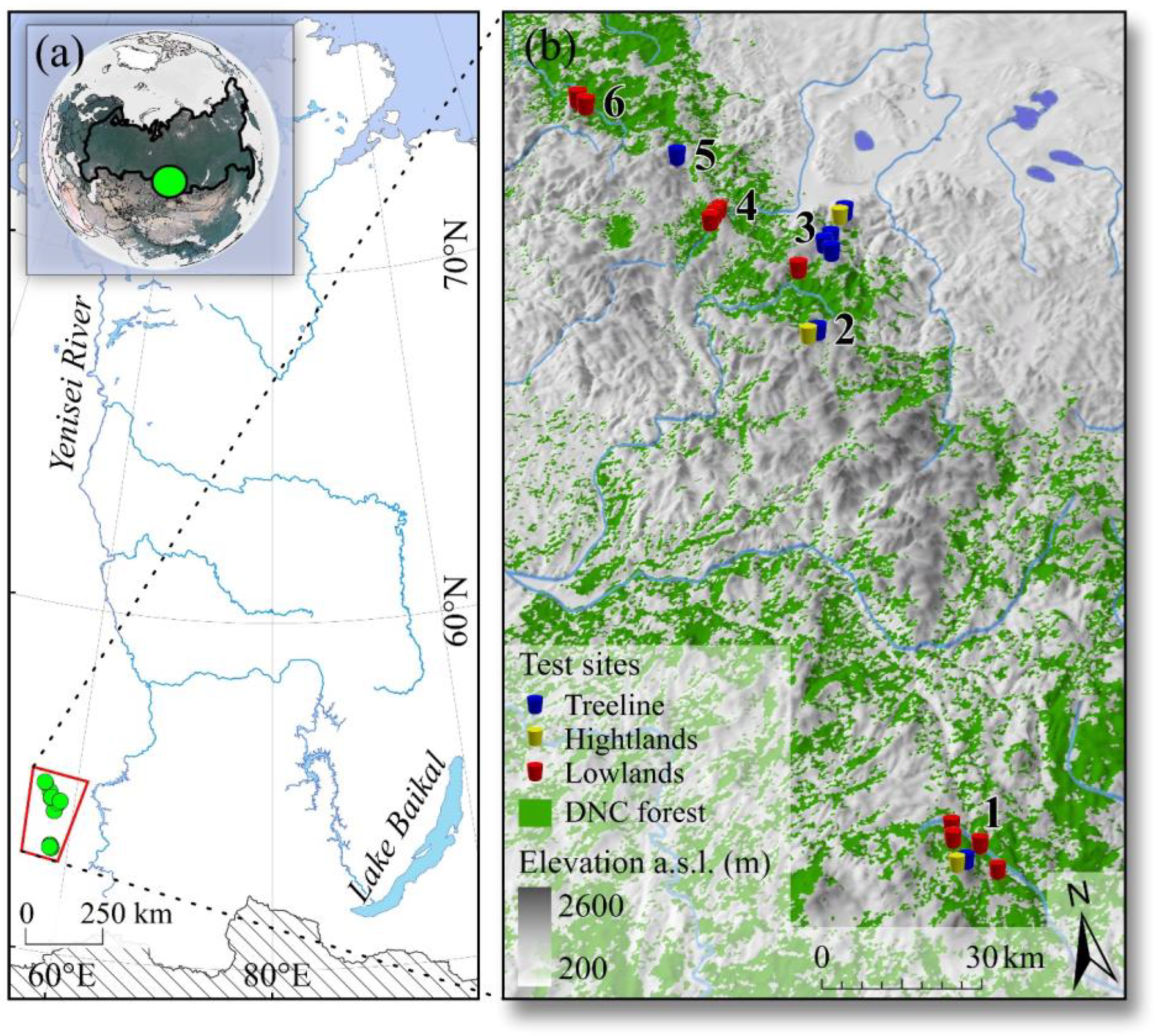
2.1. The Study Area
2.2. Climate Variables
2.3. On-the-Ground Studies
2.4. Dendrochronological Analysis
3. Results
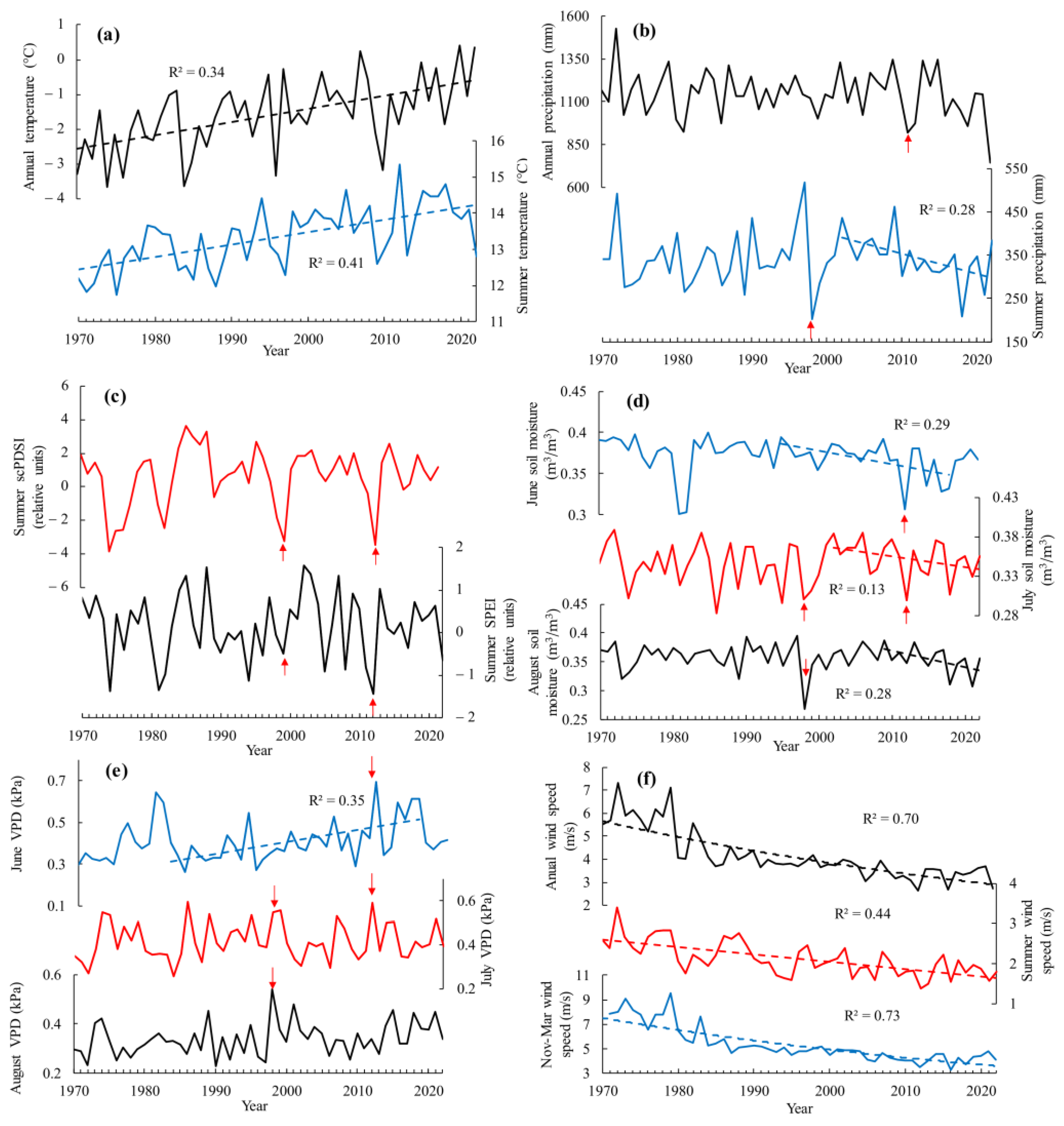
3.1. Climate Variables’ Dynamic
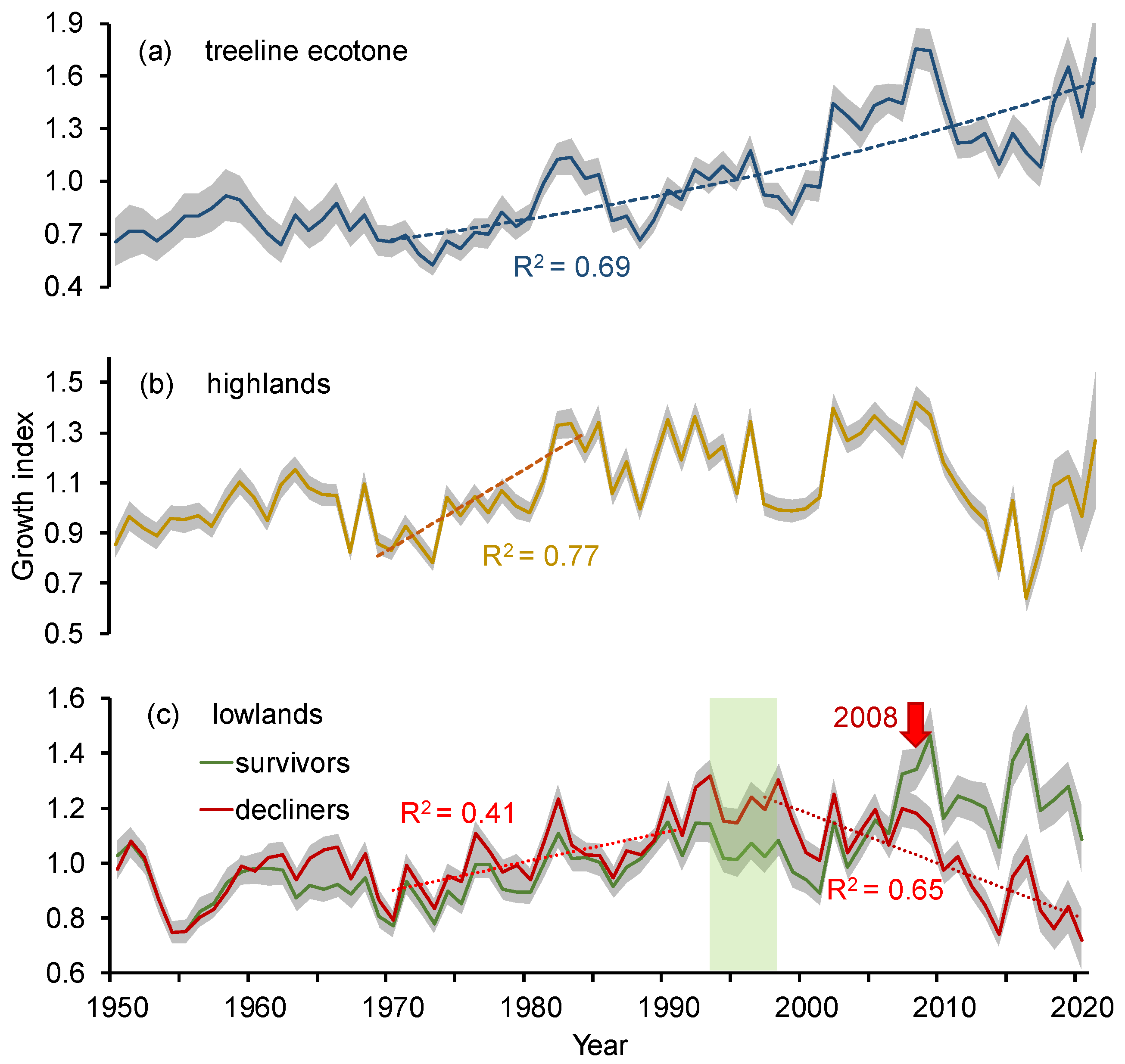
3.2. Siberian Pine Growth Dynamic along the Elevation Gradient
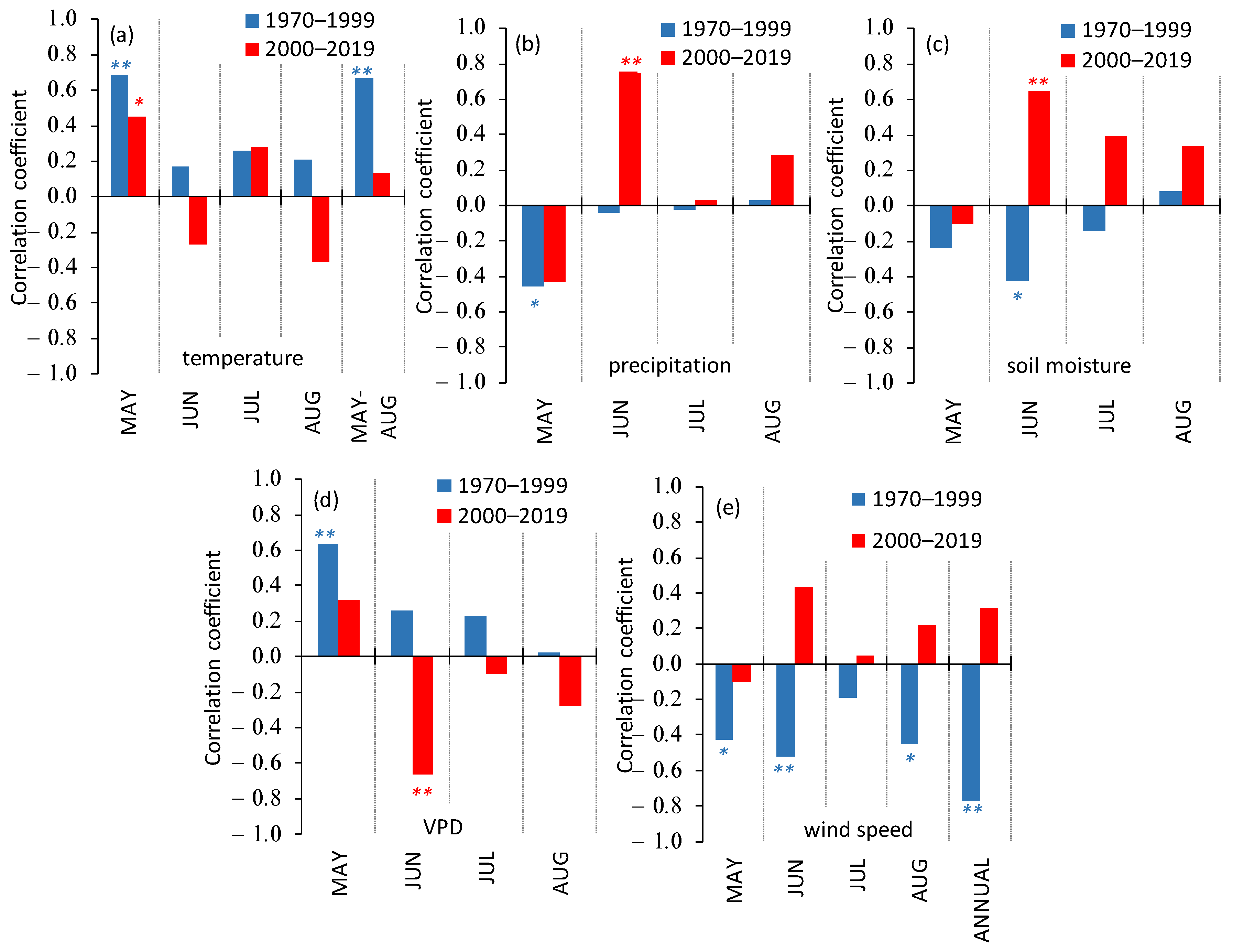
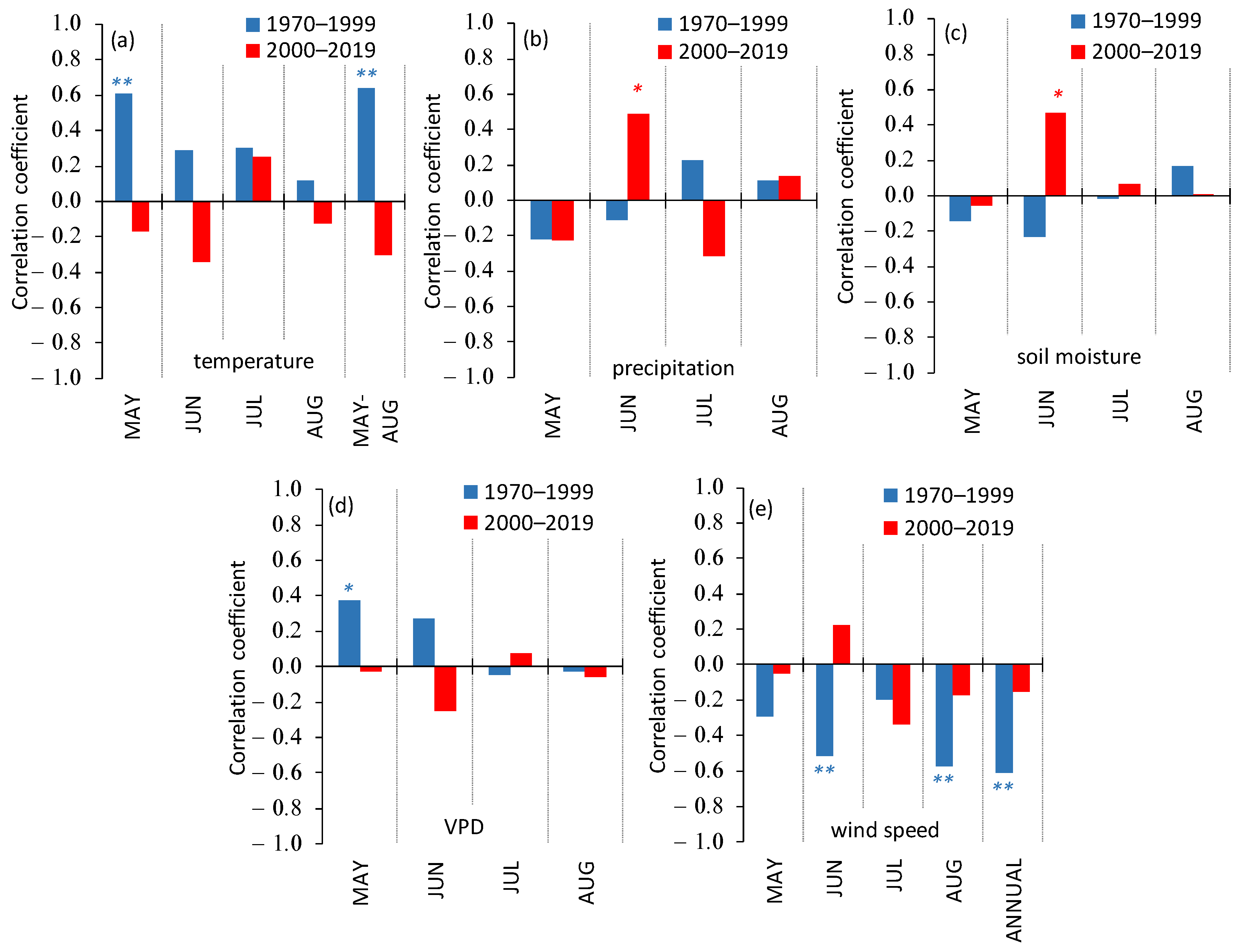
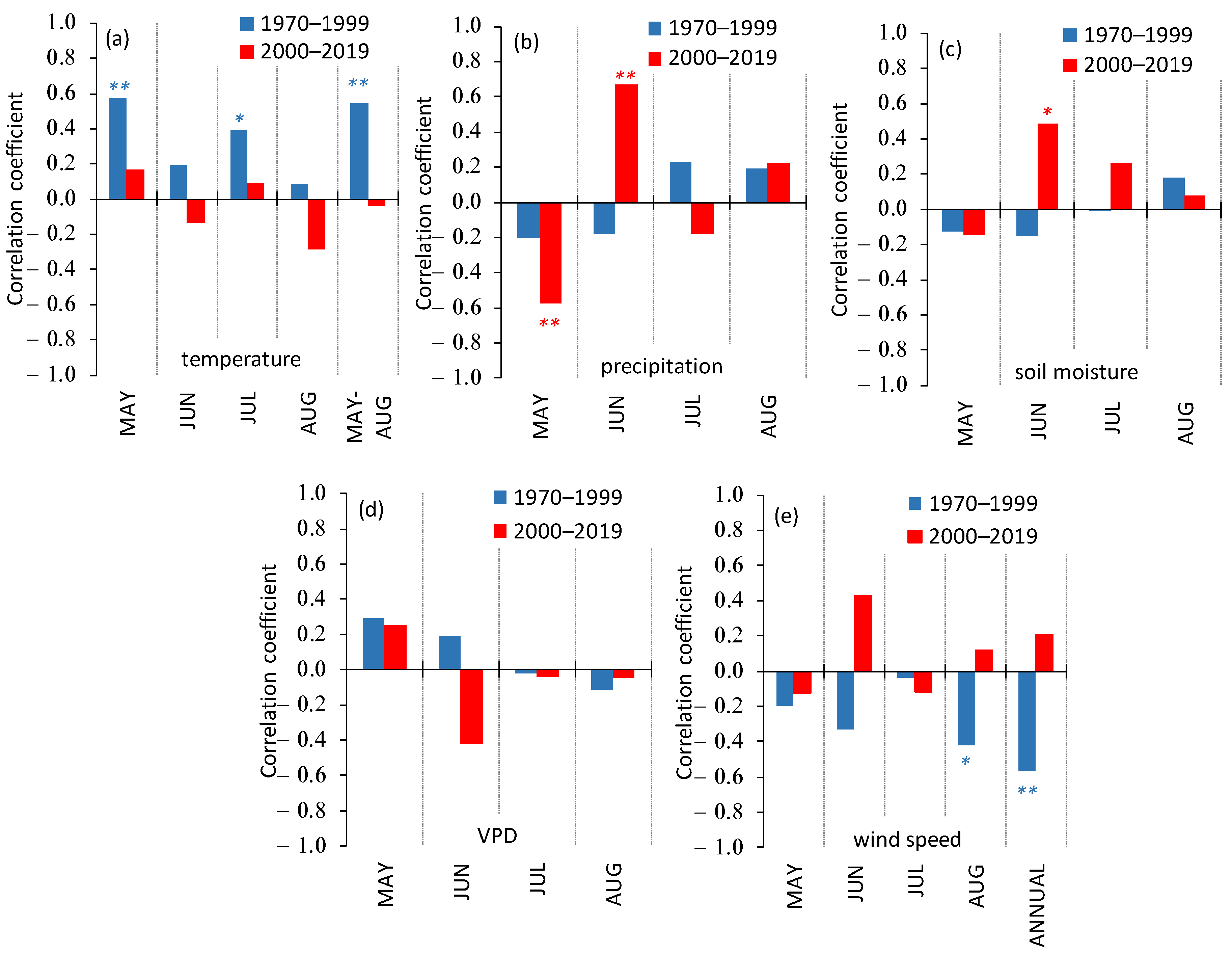
3.3. Siberian Pine Growth Dependence on Climate Variables
4. Discussion
5. Conclusions
- The consequences of climate warming differed within different elevation belts. In the treeline ecotone, the tree’s growth has continuously increased since the warming onset in the 1970s, whereas in the lowlands, the initial growth increase switched to a growth drop since the beginning of the 2000s with consequent partial mortality of Siberian pine forest. In the highlands, the tree’s growth was increasing until the end of the 1980s with the following growth fluctuations.
- The control of the Siberian pine growth by air temperature and moisture variables included two phases. Firstly, (from the warming onset in the 1970s), the trees’ GI was positively correlated with elevated temperature, whereas the GI dependence on precipitation and soil moisture was negative. During the second phase (since the warming increase after 2000), negative correlations between GI and moisture variables switched to positive ones, whereas the GI correlations with air temperature switched from positive to insignificant (except for the beginning of the growth period, i.e., during the seasonal water stress).
- The wind influence on the GI dramatically changed due to a continuous decrease in wind speed throughout the study period. Thus, the significant negative influence of winds on the trees’ growth within all elevation belts turned insignificant after 2000.
- Within the lowlands, warming-driven water stress, in combination with bark borers’ attacks, led to tree mortality, which suggests the retraction of the Siberian pine range in the lowlands of the Siberian Mountains. The projected drought increase will likely lead to the substitution of Siberian pine with drought-tolerant species.
- Afforestation within the areas of Siberian pine mortality should not be based on the planting of Siberian pine but on planting drought-tolerant species such as larch (Larix sibirica) and Scots pine (Pinus sylvestris).
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hammond, W.M.; Williams, A.P.; Abatzoglou, J.T.; Adams, H.D.; Klein, T.; López, R.; Sáenz-Romero, C.; Hartmann, H.; Breshears, D.D.; Allen, C.D. Global field observations of tree die-off reveal hotter-drought fingerprint for Earth’s forests. Nat. Commun. 2022, 13, 1761. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.P.; Davi, N.K.; Magney, T.S.; Andreu-Hayles, L.; Nachin, B.; Suran, B.; Varuolo-Clarke, A.M.; Cook, B.I.; D’Arrigo, R.D.; Pederson, N.; et al. Approaching a thermal tipping point in the Eurasian boreal forest at its southern margin. Commun. Earth Environ. 2023, 4, 247. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Im, S.T.; Petrov, I.A. Alpine ecotone in the Siberian Mountains: Vegetation response to warming. J. Mt. Sci. 2021, 18, 3099–3108. [Google Scholar] [CrossRef]
- Maher, C.; Hewitt, R.E.; Sullivan, P.F. Sufficient conditions for rapid range expansion of a boreal conifer. Nature 2022, 608, 546–551. [Google Scholar] [CrossRef]
- Watts, J.; Farina, M.; Kimball, J.; Schiferl, L.; Liu, Z.; Arndt, K.; Zona, D.; Ballantyne, A.; Euskirchen, E.S.; Parmentier, F.W.; et al. Carbon uptake in Eurasian boreal forests dominates the high-latitude net ecosystem carbon budget. Glob. Chan. Biol. 2023, 29, 1870–1889. [Google Scholar] [CrossRef] [PubMed]
- Kharuk, V.I.; Im, S.T.; Petrov, I.A.; Dvinskaya, M.L.; Shushpanov, A.S.; Golyukov, A.S. Climate-driven conifer mortality in Siberia. Global Ecol. Biogeogr. 2021, 30, 543–556. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Wu, C.; Acil, N.; Carvalhais, N.; Pugh, T.A.M.; Sadler, J.P.; Seidl, R. A climate risk analysis of Earth’s forests in the 21st century. Science 2022, 377, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Guarín, A.; Taylor, A.H. Drought triggered tree mortality in mixed conifer forests in Yosemite National Park, California, USA. For. Ecol. Manag. 2005, 218, 229–244. [Google Scholar] [CrossRef]
- Redmond, M.D.; Barger, N.N. Tree regeneration following drought- and insect-induced mortality in piñon–juniper woodlands. New Phytol. 2013, 200, 402–412. [Google Scholar] [CrossRef]
- Boyd, M.A.; Berner, L.T.; Doak, P.; Goetz, S.J.; Rogers, B.M.; Wagner, D.; Walker, X.J.; Mack, M.C. Impacts of climate and insect herbivory on productivity and physiology of trembling aspen (Populus tremuloides) in Alaskan boreal forests. Environ. Res. Lett. 2019, 14, 085010. [Google Scholar] [CrossRef]
- Yousefpour, R.; Hanewinkel, M.; Le Moguédec, G. Evaluating the suitability of management strategies of pure Norway spruce forests in the black forest area of Southwest Germany for adaptation to or mitigation of climate change. Environ. Manag. 2010, 45, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vilalta, J.; Lloret, F.; Breshears, D.D. Drought-induced forest decline: Causes, scope and implications. Biol. Lett. 2012, 8, 689–691. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Mues, V.; Moreno, A.; Hasenauer, H.; Seidl, R. Climate variability drives recent tree mortality in Europe. Glob. Chang. Biol. 2017, 23, 4788–4797. [Google Scholar] [CrossRef] [PubMed]
- Hlásny, T.; Barka, I.; Kulla, L.; Bucha, T.; Sedmák, R.; Trombik, J. Sustainable forest management in a mountain region in the Central Western Carpathians, northeastern Slovakia: The role of climate change. Reg. Environ. Chang. 2017, 17, 65–77. [Google Scholar] [CrossRef]
- Zamolodchikov, D.G. Evaluation of climate-induced changes in diversity of tree species according to forest fund data records. Biol. Bull. Rev. 2011, 131, 382–392. (In Russian) [Google Scholar]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Goulden, M.L.; Bales, R.C. California forest die-off linked to multi-year deep soil drying in 2012–2015 drought. Nat. Geosci. 2019, 12, 632–637. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]
- Kolb, T.E.; Fettig, C.J.; Ayres, M.P.; Bentz, B.J.; Hicke, J.A.; Mathiasen, R.; Stewart, J.E.; Weed, A.S. Observed and anticipated impacts of drought on forest insects and diseases in the United States. For. Ecol. Manag. 2016, 380, 321–324. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Ponomarev, E.I.; Ivanova, G.A.; Dvinskaya, M.L.; Coogan, S.C.P.; Flannigan, M.D. Wildfires in the Siberian taiga. Ambio 2021, 50, 1953–1974. [Google Scholar] [CrossRef] [PubMed]
- Rinn, F. TSAP V 3.6 Reference Manual: Computer Program for Tree-Ring Analysis and Presentation; Frank Rinn Distribution: Heidelberg, Germany, 1996. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 44, 69–75. [Google Scholar]
- Cook, E.R.; Holmes, R.L. Chronology Development, Statistical Analysis. Guide for Computer Program ARSTAN; Lab. of Tree Ring Res., the University of Arizona: Tucson, AZ, USA, 1986; pp. 50–65. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010. [Google Scholar]
- Ryan, S.E.; Porth, L.S. A Tutorial on the Piecewise Regression Approach Applied to Bedload Transport Data; Gen. Tech. Rep. RMRS-GTR-189; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2007; 41p. [Google Scholar]
- Cailleret, M.; Jansen, S.; Robert, E.M.R.; Desoto, L.; Aakala, T.; Antos, J.A.; Beikircher, B.; Bigler, C.; Bugmann, H.; Caccianiga, M.; et al. A synthesis of radial growth patterns preceding tree mortality. Glob. Chang. Biol. 2017, 23, 1675–1690. [Google Scholar] [CrossRef] [PubMed]
- Kharuk, V.I.; Ranson, K.J.; Petrov, I.A.; Dvinskaya, M.L.; Im, S.T.; Golyukov, A.S. Larch (Larix dahurica Turcz) growth response to climate change in the Siberian permafrost zone. Reg. Environ. Chang. 2019, 19, 233–243. [Google Scholar] [CrossRef]
- Stephenson, N.L.; Das, A.J.; Ampersee, N.J.; Bulaon, B.M.; Yee, J.L. Which trees die during drought? The key role of insect host-tree selection. J. Ecol. 2019, 107, 2383–2401. [Google Scholar] [CrossRef]
- Sangüesa-Barreda, G.; Linares, J.C.; Camarero, J.J. Reduced growth sensitivity to climate in bark-beetle infested Aleppo pines: Connecting climatic and biotic drivers of forest dieback. For. Ecol. Manag. 2015, 357, 126–137. [Google Scholar] [CrossRef]
- Davis, K.; Dobrowski, S.Z.; Holden, Z.A.; Higuera, P.E.; Abatzoglou, J.T. Microclimatic buffering in forests of the future: The role of local water balance. Ecography 2018, 42, 1–11. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on CLIMATE CHANGE.; Pachauri, R., Meyer, L., Eds.; IPCC: Geneva, Switzerland, 2014; Available online: https://epic.awi.de/id/eprint/37530/1/IPCC_AR5_SYR_Final.pdf (accessed on 23 November 2023).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation Belt (m) | Tree Height (m) | DBH (cm) | Species Composition | Siberian Pine Age (Years) | Number of Test Sites |
|---|---|---|---|---|---|
| Treeline ecotone 1400–1550 | 3.5 | 10 | 5Sp3L2F1B * | 65 | 10 |
| Highlands 1200–1400 | 8 | 20 | 5Sp3F1L1B | 140 | 14 |
| Lowlands 500–900 | 27 | 65 | 5Sp4F1S | 190 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kharuk, V.I.; Petrov, I.A.; Golyukov, A.S.; Im, S.T.; Shushpanov, A.S. Mountain Taiga in a Warming Climate: Contrast of Siberian Pine Growth along an Elevation Gradient. Forests 2024, 15, 50. https://doi.org/10.3390/f15010050
Kharuk VI, Petrov IA, Golyukov AS, Im ST, Shushpanov AS. Mountain Taiga in a Warming Climate: Contrast of Siberian Pine Growth along an Elevation Gradient. Forests. 2024; 15(1):50. https://doi.org/10.3390/f15010050
Chicago/Turabian StyleKharuk, Viacheslav I., Il’ya A. Petrov, Alexey S. Golyukov, Sergei T. Im, and Alexander S. Shushpanov. 2024. "Mountain Taiga in a Warming Climate: Contrast of Siberian Pine Growth along an Elevation Gradient" Forests 15, no. 1: 50. https://doi.org/10.3390/f15010050