
Early Diagnosis of Pine Wilt Disease in Pinus thunbergii Based on Chlorophyll Fluorescence Parameters
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. PWN Inoculation Experiment and Disease Observation
2.3. Measurement of Chlorophyll Fluorescence Kinetic Parameters in P. thunbergii Needles
2.4. Data Analysis
3. Results
3.1. Statistics on the Disease Incidence and Disease Index of P. thunbergii
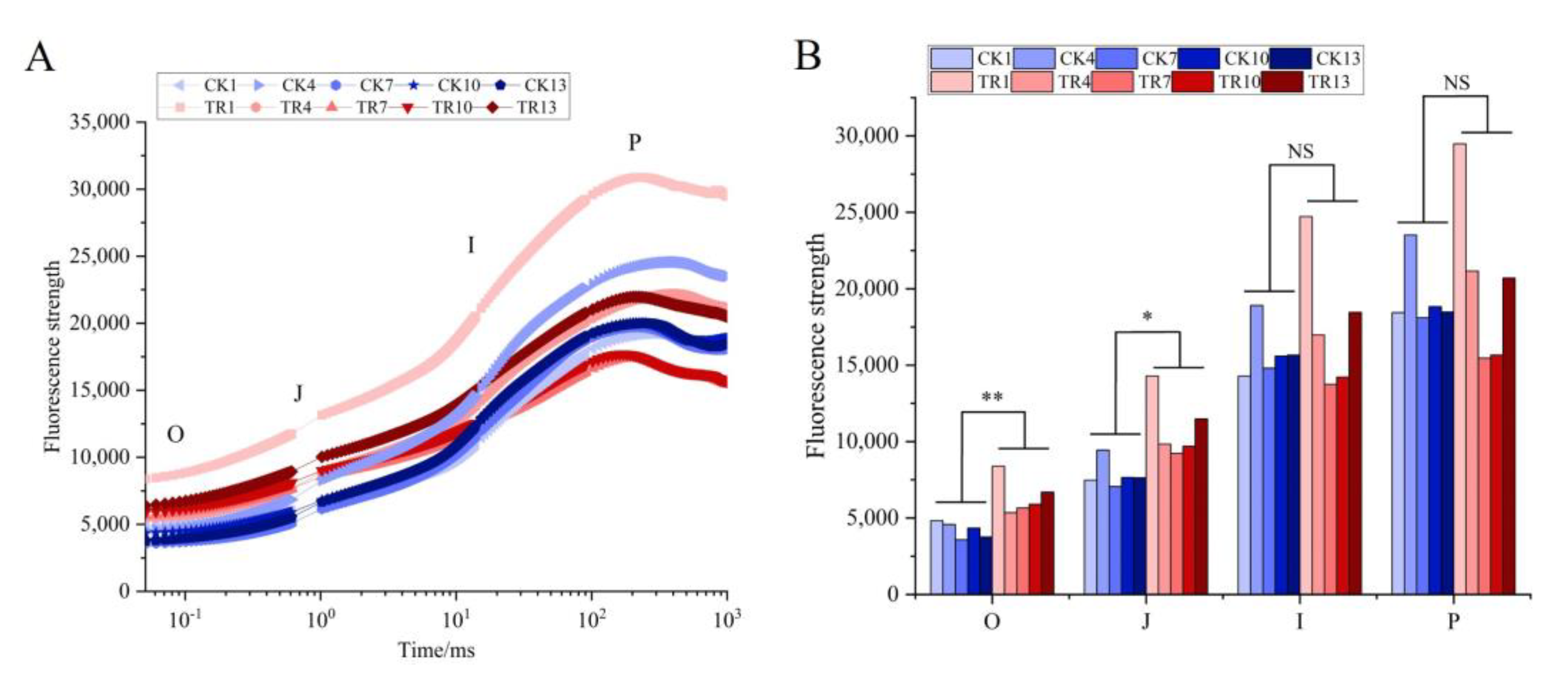
3.2. Effect of PWN on the Rapid Chlorophyll Fluorescence Induction Kinetic Curves of P. thunbergii
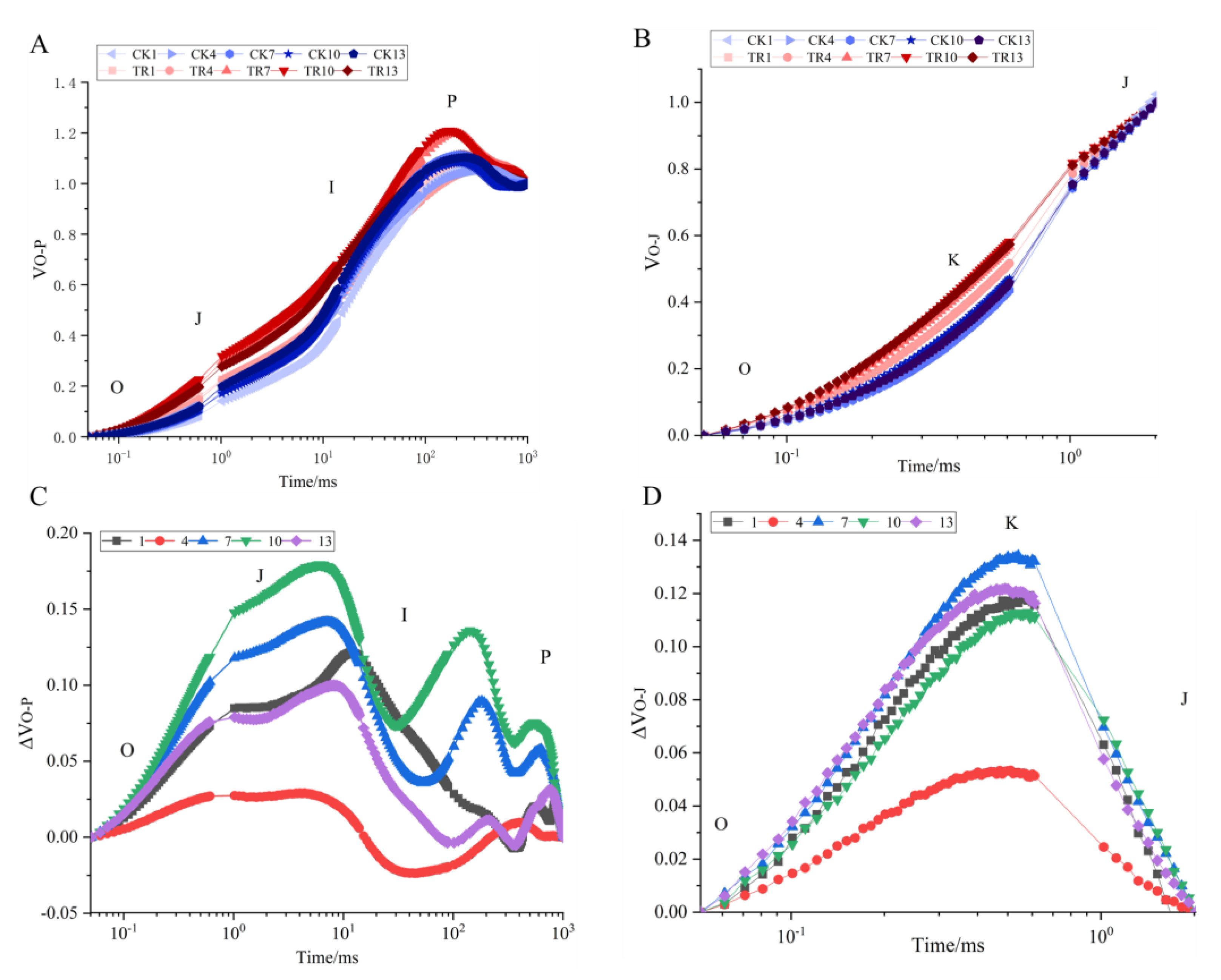
3.3. Effect of PWN on the Electron Acceptor Side and Electron Donor Side Transfer Capacity of PSII in P. thunbergii
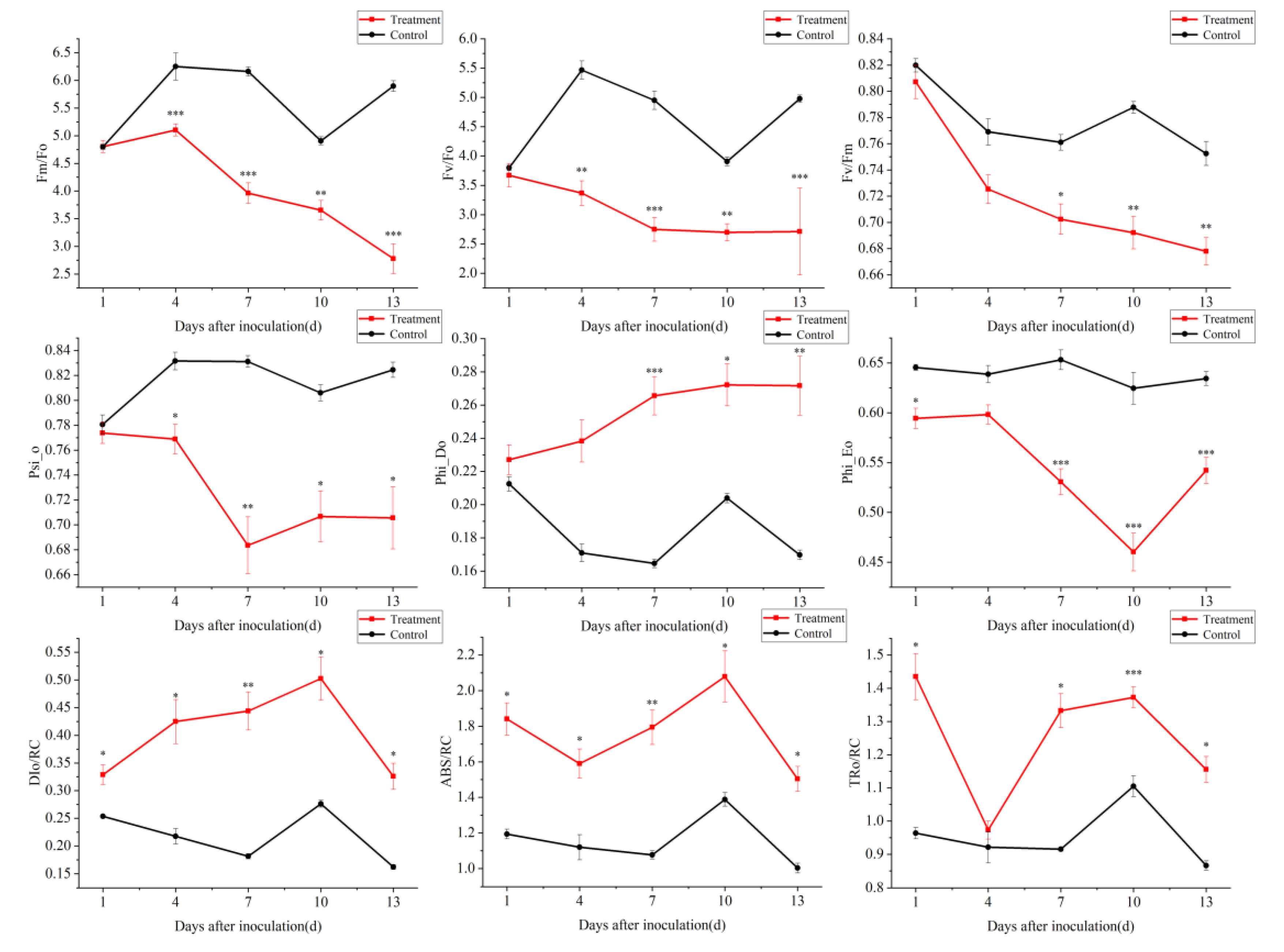
3.4. Effect of PWN on Chlorophyll Fluorescence Parameters of P. thunbergii Needles
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.A.; et al. Genomic Insights into the Origin of Parasitism in the Emerging Plant Pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futai, T. Field Diagnosis of the Asymptomatic Carrier of Pinewood Nematode. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Springer: Dordrecht, The Netherlands, 2008; pp. 279–290. [Google Scholar]
- Jiang, M.; Huang, B.; Xu, Y.U.; Zheng, W.T.; Jin, Y.L.; Liao, M.N.; Jian, N.I. Distribution, Damage and Control of Pine Wilt Disease. J. Zhejiang For. Sci. Technol. 2018, 38, 83–91. [Google Scholar]
- Shin, S.C.; Moon, I.S.; Han, H. Current research and management on pine wilt disease in Korea. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2010, 34, 167–168. [Google Scholar]
- Mota, M.; Bonifácio, L.; Bravo, M.A.; Naves, P.; Penas, A.C.; Pires, J.; Sousa, E.; Vieira, P. Discovery of PWN in Portugal and in Europe. PWN 2003, 1, 1–5. [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus Xylophilus, causal agent of pine wilt disease on in northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef]
- Hunt, D. Pine wilt disease: A worldwide threat to forest ecosystems. Nematology 2009, 11, 315–316. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Lv, Q.; Yu, C.-M.; Li, Q.; Liu, H.-X.; Zhang, X.-Y. A review of the early diagnosis technology of pine wood nematode disease. J. Shandong Agric. Univ. (Nat. Sci. Ed.) 2014, 45, 158–160+176+153. [Google Scholar]
- Ning, T.; Fan, J.-T.; Fang, Y.-L.; Sun, J.-H. Changes in contents of host volatile terpenes under different damaged states and electroantennogram response of Monochamus alternatus Hope to these volatiles. J. Entomol. 2006, 49, 179–188. [Google Scholar]
- Vellosillo, T.; Vicente, J.; Kulasekaran, S.; Hamberg, M.; Castresana, C. Emerging complexity in reactive oxygen species production and signaling during the response of plants to pathogens. Plant Physiol. 2010, 158, 444–448. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-H.; Ye, J.-R.; Wei, C.-J.; Yang, X.-M. Effects of Pine Wood Nematode Infection on Metabolism of Active Oxygen in Japanese Black Pine and Slash Pine Seedlings. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2002, 26, 19–22. [Google Scholar]
- Oku, H. Role of phytotoxins in pine wilt diseases. J. Nematol. 1988, 20, 245–251. [Google Scholar] [PubMed]
- Gao, R.-H.; Luo, Y.-Q.; Shi, J. Effect of Pine Wilt Disease Infection on Leaf Photosynthetic Characteristics of Masson Pine. For. Res. 2019, 32, 65–73. [Google Scholar]
- Zhang, H.; Huang, X.-F.; Xu, H.-C. Changes of photosynthesis and antioxidant enzyme activities at different stage naturally infected by Bursaphelenchus xylophilus. J. Environ. Entomol. 2014, 36, 139–144. [Google Scholar]
- Linn, A.I.; Zeller, A.K.; Pfündel, E.E.; Gerhards, R. Features and applications of a field imaging chlorophyll fluorometer to measure stress in agricultural plants. Precis. Agric. 2020, 22, 947–963. [Google Scholar] [CrossRef]
- Recchia, I.; Sparla, F.; Pupillo, P. Photosynthetic properties of spring geophytes assessed by chlorophyll fluorescence analysis. Plant Physiol. Biochem. 2017, 118, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Smillie, R.M.; Nott, R. Salt tolerance in crop plants monitored by chlorophyll fluorescence in vivo. Plant Physiol. 1982, 70, 1049–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méthy, M.; Olioso, A.; Trabaud, L. Chlorophyll fluorescence as a tool for management of plant resources. Remote Sens. Environ. 1994, 47, 2–9. [Google Scholar] [CrossRef]
- Chávez-Arias, C.C.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological, Biochemical and Chlorophyll Fluorescence Parameters of Physalis peruviana L. Seedlings Exposed to Different Short-Term Waterlogging Periods and Fusarium Wilt Infection. Agronomy 2019, 9, 213. [Google Scholar] [CrossRef]
- Banks, J.M. Chlorophyll Fluorescence as a Tool to Identify Drought Stress in Acer genotypes. Environ. Exp. Bot. 2018, 155, 118–127. [Google Scholar] [CrossRef]
- Schansker, G.; Tóth, S.Z.; Strasser, R.J. Methylviologen and dibromothymoquinone treatments of pea leaves reveal the role of photosystem I in the Chl a fluorescence rise OJIP. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2005, 1706, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Melakeberhan, H.; Toivonen, P.M.A.; Vidaver, W.E.; Webster, J.M.; Dube, S.L. Effect of Bursaphelenchus xylophilus on the water potential and water-splitting complex of photosystem II of Pinus sylvestris seedlings. Physiol. Mol. Plant Pathol. 1991, 38, 83–91. [Google Scholar] [CrossRef]
- Xie, L.-Q.; Ju, Y.-W.; Yang, Z.-D.; Zhao, B.-G. Dynamics of densities of bacteria and nematode in the branches of Pinus thunbergii inoculated with Bursaphelenchus xylophilus. J. Zhejiang A F Univ. 2005, 22, 310–314. [Google Scholar]
- Tan, J.-J.; Wu, H.-P.; Chen, Y.; Yang, R.-Z. Effect of Bursaphelenchus xylophilus on the Free Proline Content in Leaves of Pinus thunbergii. J. South China Agric. Univ. 2000, 21, 92. [Google Scholar]
- Wang, G.; Bi, A.; Amombo, E.; Li, H.; Zhang, L.; Cheng, C.; Hu, T.; Fu, J. Exogenous Calcium Enhances the Photosystem II Photochemistry Response in Salt Stressed Tall Fescue. Front. Plant Sci. 2017, 8, 2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. Probing Photosynth. Mech. Regul. Adapt. 2000, 445–483. Available online: https://www.researchgate.net/publication/252250818_The_fluorescence_transient_as_a_tool_to_characterize_and_screen_photosynthetic_samples (accessed on 10 January 2023).
- Kan, X.; Ren, J.; Chen, T.; Cui, M.; Li, C.; Zhou, R.; Zhang, Y.; Liu, H.; Deng, D.; Yin, Z. Effects of salinity on photosynthesis in maize probed by prompt fluorescence, delayed fluorescence and P700 signals. Environ. Exp. Bot. 2017, 140, 50–64. [Google Scholar] [CrossRef]
- Ajigboye, O.O.; Bousquet, L.; Murchie, E.H.; Ray, R.V. Chlorophyll fluorescence parameters allow the rapid detection and differentiation of plant responses in three different wheat pathosystems. Funct. Plant Biol. 2016, 43, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Swarbrick, P.J.; Schulze-Lefert, P.; Scholes, J.D. Metabolic consequences of susceptibility and resistance (race-specific and broad-spectrum) in barley leaves challenged with powdery mildew. Plant Cell Environ. 2006, 29, 1061–1076. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.; Liu, Y.; Captoline, I.; Li, X.; Ye, D.; Wu, R. Citrus Huanglongbing detection based on polyphasic chlorophyll a fluorescence coupled with machine learning and model transfer in two citrus cultivars. Comput. Electron. Agric. 2021, 187, 106289. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, J.; Xi, Y.; Zhang, L.; Gao, Q.; Wang-Pruski, G. Effects of Autotoxicity on Seed Germination, Gas Exchange Attributes and Chlorophyll Fluorescence in Melon Seedlings. J. Plant Growth Regul. 2021, 41, 993–1003. [Google Scholar] [CrossRef]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Govindjee, N.; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2010, 1797, 1428–1438. [Google Scholar] [CrossRef] [PubMed]
- Fei, L.; Chu, J.; Zhang, X.; Dong, S.; Dai, X.; He, M. Physiological and Proteomic Analyses Indicate Delayed Sowing Improves Photosynthetic Capacity in Wheat Flag Leaves Under Heat Stress. Front. Plant Sci. 2022, 13, 848464. [Google Scholar] [CrossRef] [PubMed]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2011, 1817, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.-B.; Rui, L.; Feng, Y.-Q.; Wu, X.-Q. Autophagy contributes to resistance to the oxidative stress induced by pine reactive oxygen species metabolism, promoting infection by Bursaphelenchus xylophilus. Pest Manag. Sci. 2020, 76, 2755–2767. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, S.; Parihar, P.; Mishra, R.K.; Tripathi, D.K.; Singh, V.P.; Chauhan, D.K.; Prasad, S.M. Reactive Oxygen Species (ROS): Beneficial Companions of Plants' Developmental Processes. Front. Plant Sci. 2016, 7, 1299. [Google Scholar] [CrossRef] [Green Version]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhong, S.; Zhang, M.; Liang, Y.; Gong, G.; Chang, X.; Tan, F.; Yang, H.; Qiu, X.; Luo, L.; et al. Potential Role of Photosynthesis in the Regulation of Reactive Oxygen Species and Defence Responses to Blumeria graminis f. sp. tritici in Wheat. Int. J. Mol. Sci. 2020, 21, 5767. [Google Scholar] [CrossRef]
- Sperdouli, I.; Mellidou, I.; Moustakas, M. Harnessing Chlorophyll Fluorescence for Phenotyping Analysis of Wild and Cultivated Tomato for High Photochemical Efficiency under Water Deficit for Climate Change Resilience. Climate 2021, 9, 154. [Google Scholar] [CrossRef]
- Meravi, N.; Kumar Prajapati, S. Seasonal variation in chlorophyll a fluorescence of Butea monosperma. Biol. Rhythm Res. 2018, 51, 879–886. [Google Scholar] [CrossRef]
- Else, M.A.; Janowiak, F.; Atkinson, C.J.; Jackson, M.B. Root signals and stomatal closure in relation to photosynthesis, chlorophyll a fluorescence and adventitious rooting of flooded tomato plants. Ann. Bot. 2008, 103, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Liu, N.; Zhao, J.; Ge, F.; Xu, Y.; Chen, Y. Disturbance of photosystem II-oxygen evolution complex induced the oxidative damage in Chlorella vulgaris under the stress of cetyltrimethylammonium chloride. Chemosphere 2019, 223, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cendrero-Mateo, M.P.; Moran, M.S.; Papuga, S.A.; Thorp, K.R.; Alonso, L.; Moreno, J.; Ponce-Campos, G.; Rascher, U.; Wang, G. Plant chlorophyll fluorescence: Active and passive measurements at canopy and leaf scales with different nitrogen treatments. J. Exp. Bot. 2015, 67, 275–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, J.M.; Cordon, G.B.; Lagorio, M.G. Re-absorption and scattering of chlorophyll fluorescence in canopies: A revised approach. Remote Sens. Environ. 2020, 246, 111860. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Disease Incidence/% | Disease Index | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 4 | 7 | 10 | 13 | 16 | 30 | 1 | 4 | 7 | 10 | 13 | 16 | 30 | |
| AMA3 | 0 | 0 | 0 | 0 | 33.3 | 60 | 100 | 0 | 0 | 0 | 0 | 8.3 | 16.7 | 100 |
| CK | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Days after Inoculation | The Relative Rates of Change in Chlorophyll Fluorescence Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| ΔFm/Fo /% | ΔFv/Fo /% | ΔFv/Fm /% | ΔPsi_o /% | ΔPhi_Do /% | ΔPhi_Eo /% | ΔDIo/RC /% | ΔABS/RC /% | ΔTRo/RC /% | |
| 1 | 0.107 | −3.300 | −0.875 | −1.539 | 6.866 | −7.916 | 29.728 | 54.155 | 48.888 |
| 4 | −18.340 | −38.398 | −7.535 | −5.689 | 39.374 | −6.351 | 95.139 | 41.942 | 5.620 |
| 7 | −35.692 | −44.484 | −17.756 | −7.699 | 61.330 | −18.749 | 144.725 | 66.697 | 45.578 |
| 10 | −25.521 | −30.927 | −12.316 | −12.135 | 33.402 | −26.301 | 81.783 | 49.739 | 24.224 |
| 13 | −52.893 | −45.520 | −14.428 | −9.918 | 59.959 | −14.529 | 101.160 | 49.950 | 33.453 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Zhang, M.; Hu, J.; Pan, M.; Shen, L.; Ye, J.; Tan, J. Early Diagnosis of Pine Wilt Disease in Pinus thunbergii Based on Chlorophyll Fluorescence Parameters. Forests 2023, 14, 154. https://doi.org/10.3390/f14010154
Liu F, Zhang M, Hu J, Pan M, Shen L, Ye J, Tan J. Early Diagnosis of Pine Wilt Disease in Pinus thunbergii Based on Chlorophyll Fluorescence Parameters. Forests. 2023; 14(1):154. https://doi.org/10.3390/f14010154
Chicago/Turabian StyleLiu, Fei, Maojiao Zhang, Jiafeng Hu, Min Pan, Luyang Shen, Jianren Ye, and Jiajin Tan. 2023. "Early Diagnosis of Pine Wilt Disease in Pinus thunbergii Based on Chlorophyll Fluorescence Parameters" Forests 14, no. 1: 154. https://doi.org/10.3390/f14010154