
Effects of Extreme Drought and Heat Events on Leaf Metabolome of Black Alder (Alnus glutinosa L.) Growing at Neighboring Sites with Different Water Availability

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
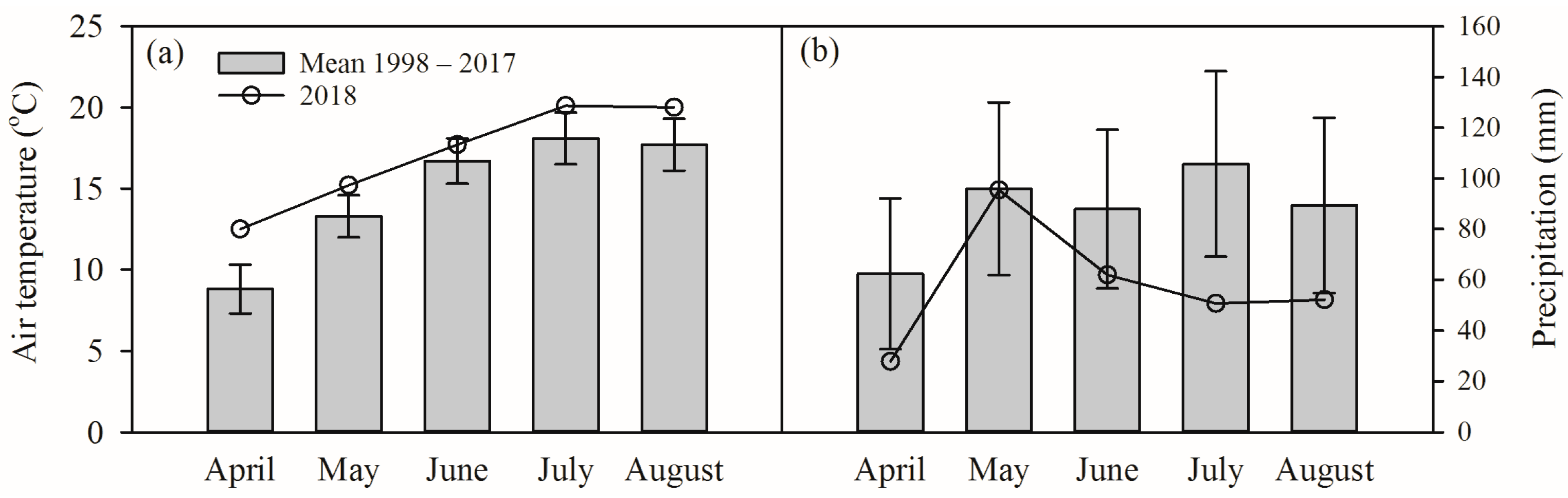
2.1. Experimental Conditions and Plant Material
2.2. Leaf Hydration Determination
2.3. Determination of Hydrogen Peroxide (H2O2), Malondialdehyde (MDA) Contents, and In Vitro Activities of Glutathione Reductase (GR) and Dehydroascorbate Reductase (DHAR)
2.4. Thiols and Ascorbate Measurement
2.5. Soluble Protein and Sugar Determination
2.6. Determination of Anions
2.7. Low Molecular Weight Soluble Metabolites Analyzed by Gas Chromatography-Mass Spectrometry (GC-MS)
2.8. Statistical Analysis
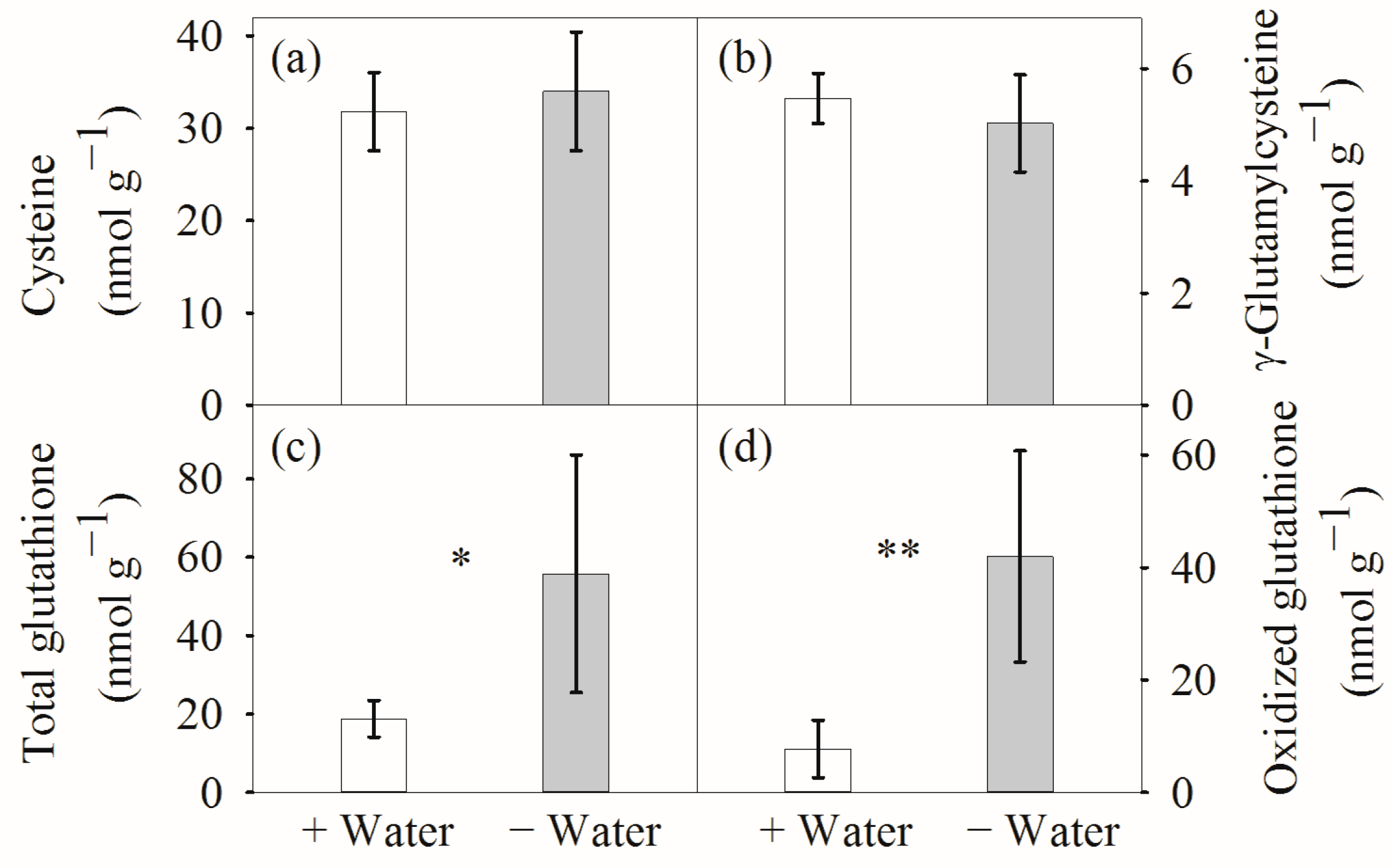
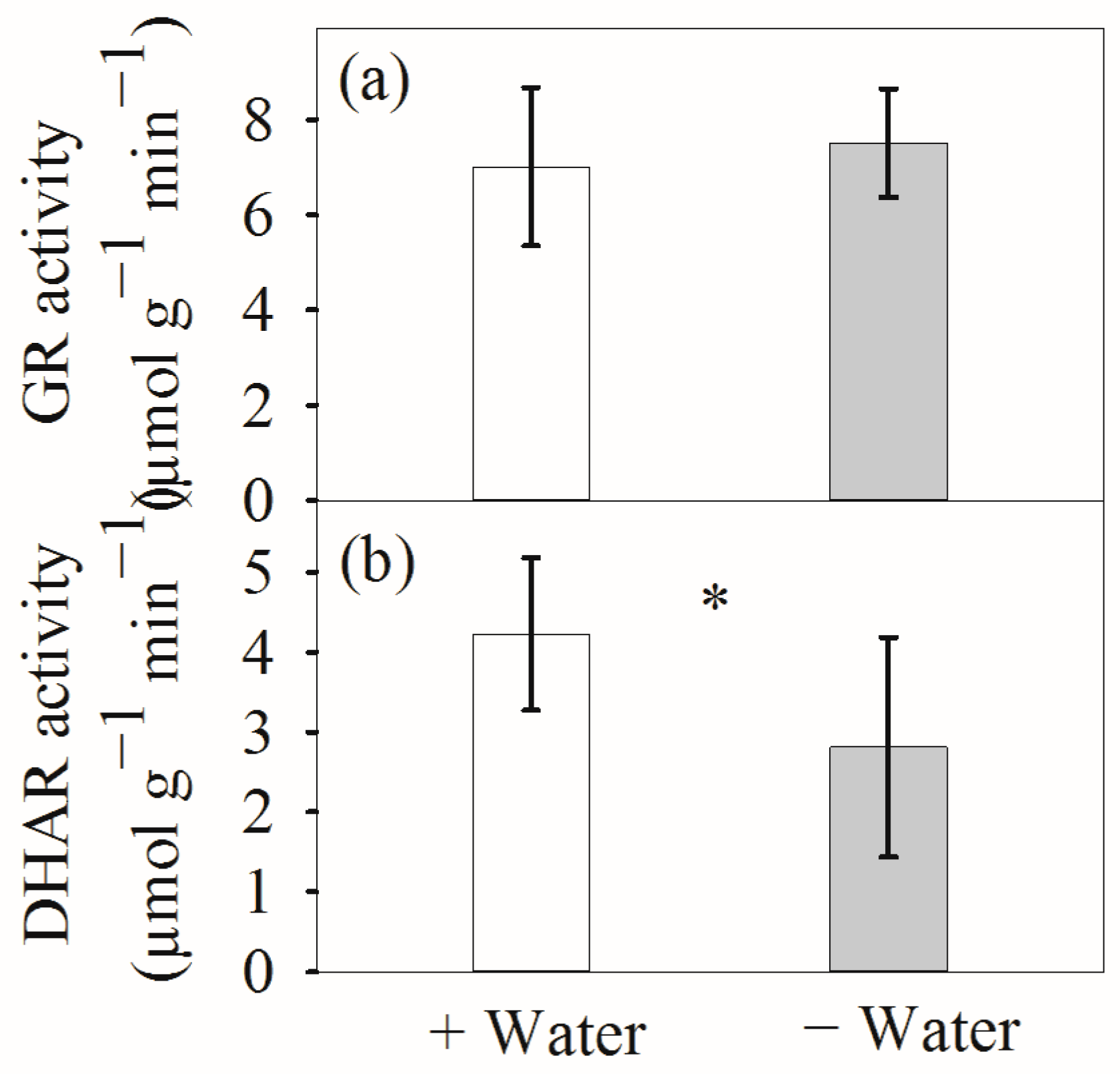
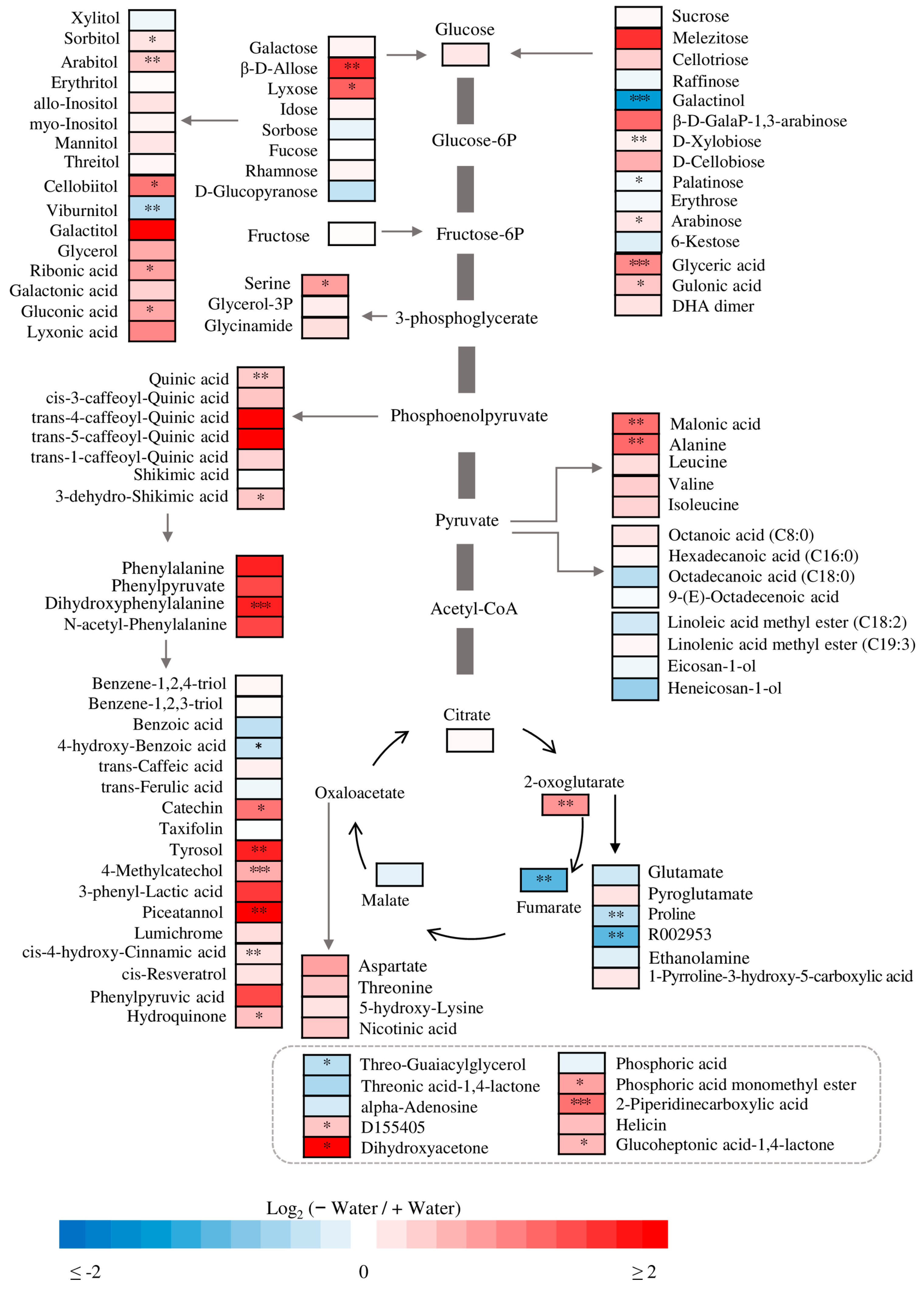
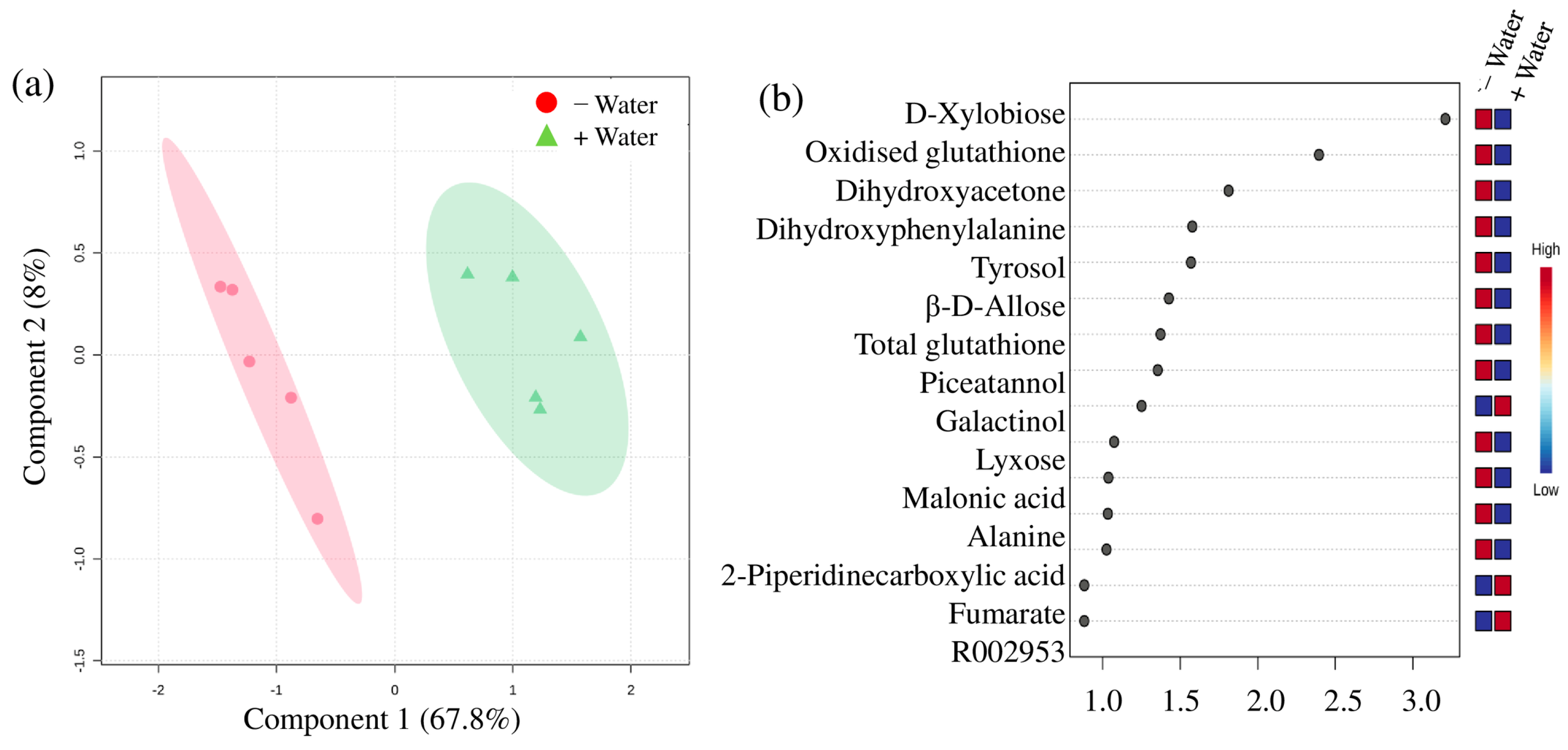
3. Results
4. Discussion
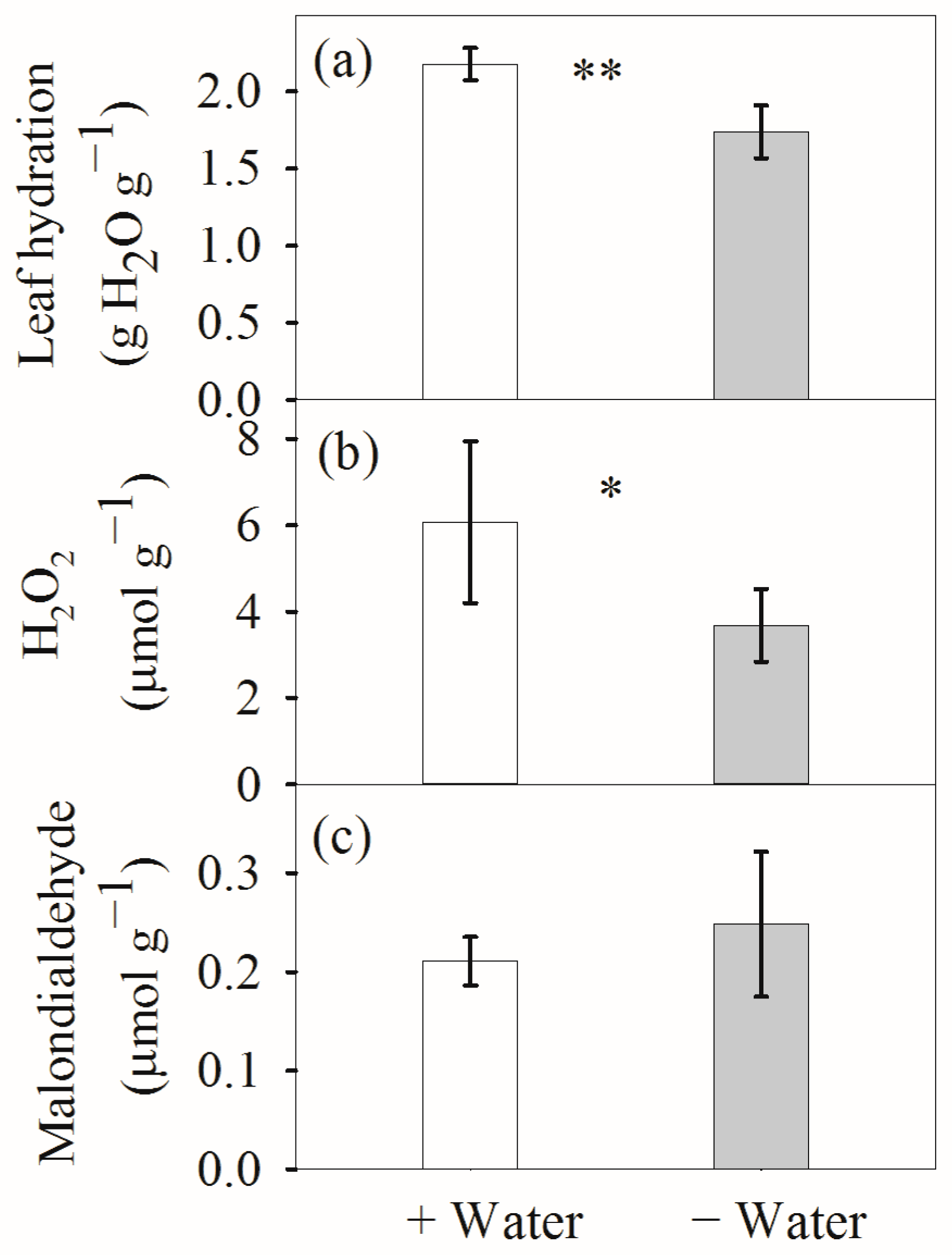
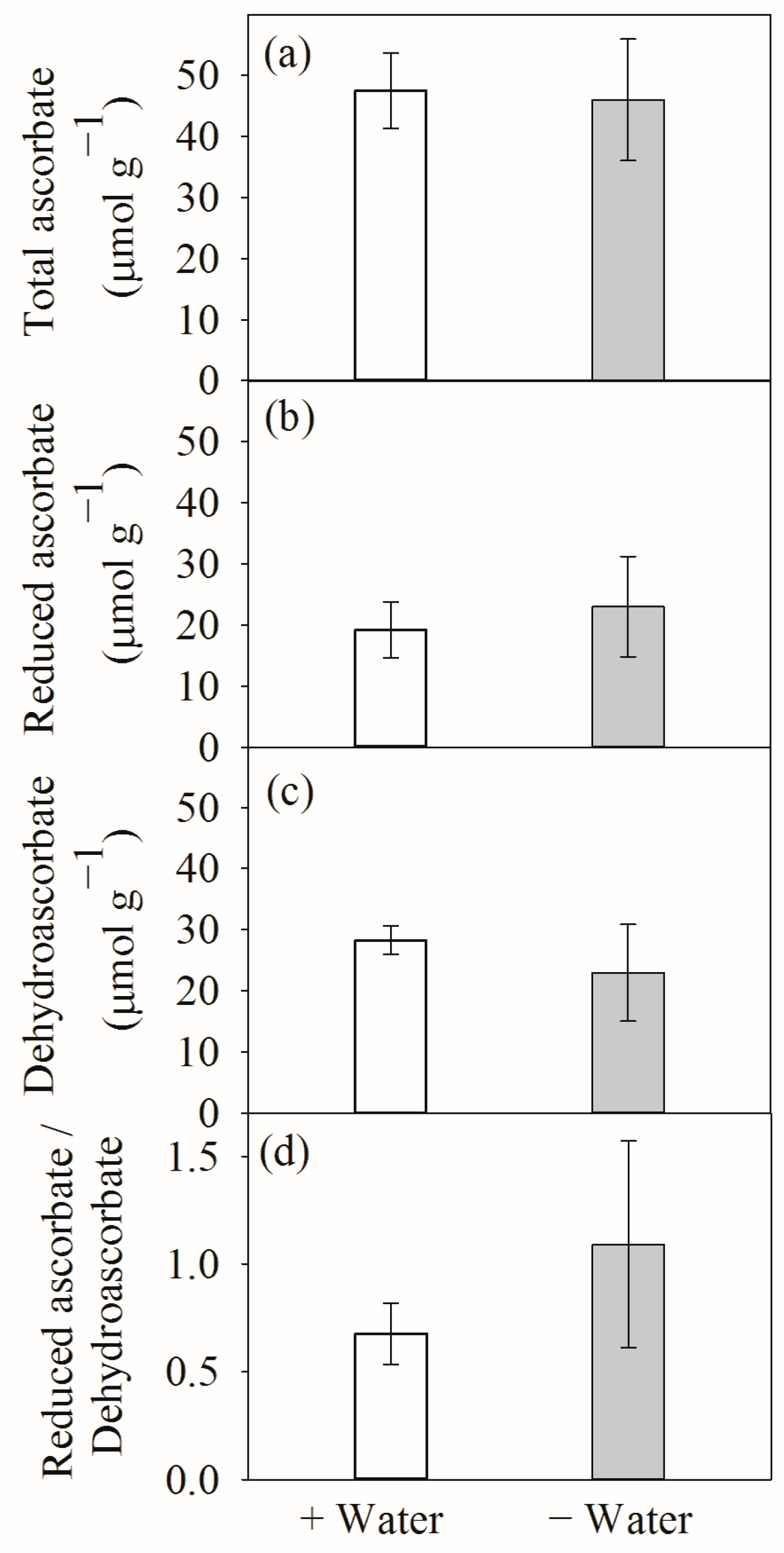
4.1. Leaf Hydration and Antioxidative Systems at Different Water Conditions
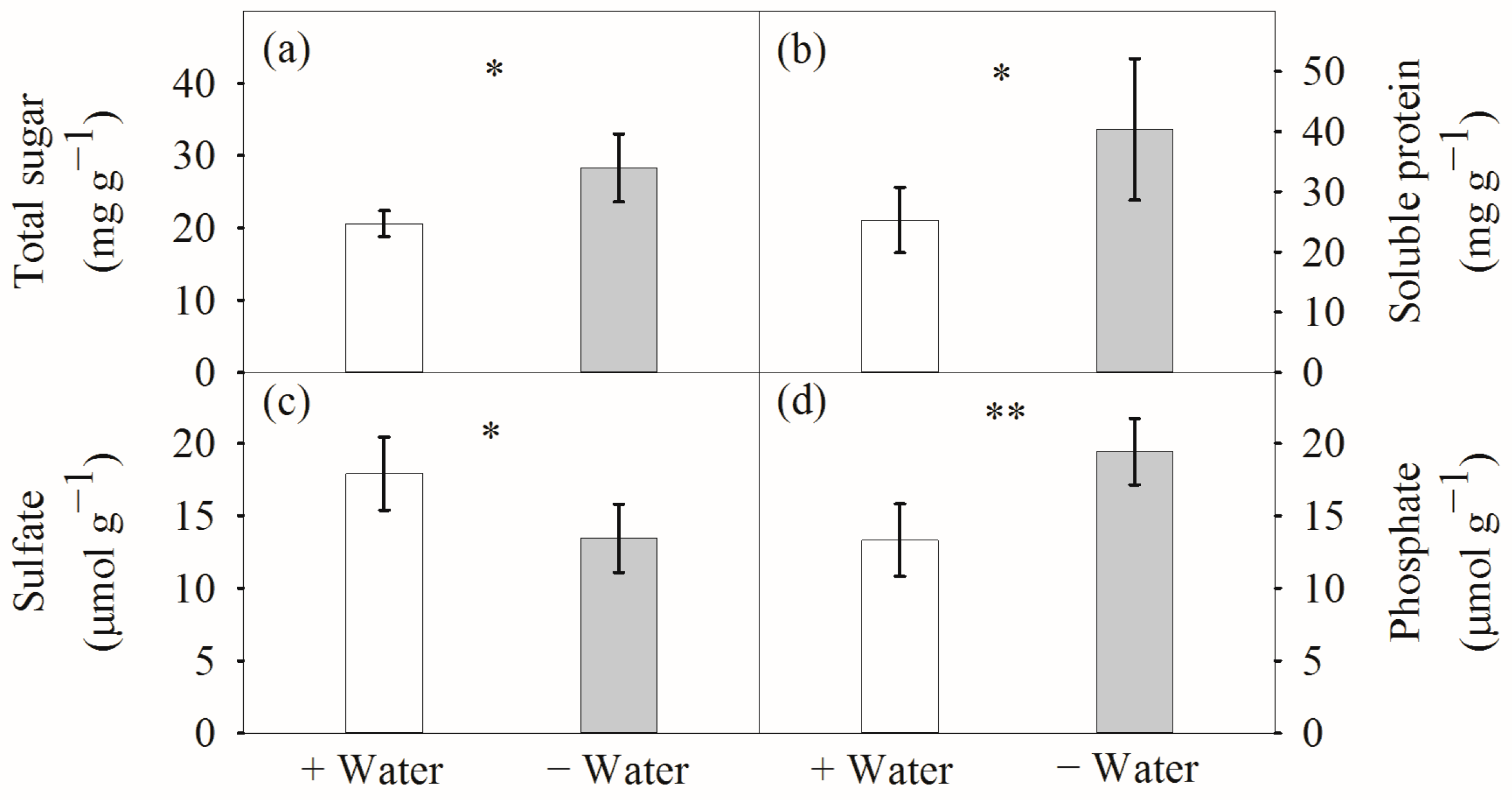
4.2. Compatible Solutes at Different Water Conditions
4.3. Responses of Anions to Water Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dybala, K.E.; Matzek, V.; Gardali, T.; Seavy, N.E. Carbon Sequestration in Riparian Forests: A Global Synthesis and Meta-analysis. Glob. Chang. Biol. 2019, 25, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlowski, T.T. Physiological-Ecological Impacts of Flooding on Riparian Forest Ecosystems. Wetlands 2002, 22, 550–561. [Google Scholar] [CrossRef]
- Rodríguez-González, P.M.; Colangelo, M.; Sánchez-Miranda, Á.; Sánchez-Salguero, R.; Campelo, F.; Rita, A.; Gomes Marques, I.; Albuquerque, A.; Ripullone, F.; Camarero, J.J. Climate, Drought and Hydrology Drive Narrow-Leaved Ash Growth Dynamics in Southern European Riparian Forests. For. Ecol. Manag. 2021, 490, 119128. [Google Scholar] [CrossRef]
- Hu, H.; Chen, H.; Hu, T.; Zhang, J. Adaptability Comparison between the Seedlings of Eucalyptus grandis and Alnus cremastogyne under the Condition of Continuous Drought Stress. J. Agric. Sci. 2012, 4, 75–86. [Google Scholar] [CrossRef]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Zhang, L.; Wu, X.; Chen, W.; Song, D. Phosphorous Fertilization Alleviates Drought Effects on Alnus Cremastogyne by Regulating Its Antioxidant and Osmotic Potential. Sci. Rep. 2018, 8, 5644. [Google Scholar] [CrossRef] [PubMed]
- Hultine, K.R.; Bush, S.E.; West, A.G.; Ehleringer, J.R. Population Structure, Physiology and Ecohydrological Impacts of Dioecious Riparian Tree Species of Western North America. Oecologia 2007, 154, 85–93. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. IPCC Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; p. 41. [Google Scholar]
- Kominoski, J.S.; Shah, J.J.F.; Canhoto, C.; Fischer, D.G.; Giling, D.P.; González, E.; Griffiths, N.A.; Larrañaga, A.; LeRoy, C.J.; Mineau, M.M.; et al. Forecasting Functional Implications of Global Changes in Riparian Plant Communities. Fron. Ecol. Environ. 2013, 11, 423–432. [Google Scholar] [CrossRef]
- Marques, I.G.; Campelo, F.; Rivaes, R.; Albuquerque, A.; Ferreira, M.T.; Rodríguez-González, P.M. Tree Rings Reveal Long-Term Changes in Growth Resilience in Southern European Riparian Forests. Dendrochronologia 2018, 52, 167–176. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Hinckley, T.M. Water Relations Characteristics of Alnus rubra and Populus trichocarpa: Responses to Field Drought. Can. J. Forest Res. 1988, 18, 1159–1166. [Google Scholar] [CrossRef]
- Hacke, U.; Sauter, J.J. Drought-Induced Xylem Dysfunction in Petioles, Branches, and Roots of Populus Balsamifera L. and Alnus glutinosa (L.) Gaertn. Plant Physiol. 1996, 111, 413–417. [Google Scholar] [CrossRef]
- Rennenberg, H.; Loreto, F.; Polle, A.; Brilli, F.; Fares, S.; Beniwal, R.S.; Gessler, A. Physiological Responses of Forest Trees to Heat and Drought. Plant Biol. 2006, 8, 556–571. [Google Scholar] [CrossRef]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The Interdependence of Mechanisms Underlying Climate-Driven Vegetation Mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How Tree Roots Respond to Drought. Front. Plant Sci. 2015, 6, 547. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Duke, N.C.; Kovacs, J.M.; Griffiths, A.D.; Preece, L.; Hill, D.; Oosterzee, P.V.; Mackenzie, J.; Morning, H.S.; Burrows, D. Large-Scale Dieback of Mangroves in Australia’s Gulf of Carpentaria: A Severe Ecosystem Response, Coincidental with an Unusually Extreme Weather Event. Mar. Freshw. Res. 2017, 68, 1816–1829. [Google Scholar] [CrossRef]
- Restaino, C.; Young, D.J.N.; Estes, B.; Gross, S.; Wuenschel, A.; Meyer, M.; Safford, H. Forest Structure and Climate Mediate Drought-Induced Tree Mortality in Forests of the Sierra Nevada, USA. Ecol. Appl. 2019, 29, e01902. [Google Scholar] [CrossRef]
- Stella, J.C.; Riddle, J.; Piégay, H.; Gagnage, M.; Trémélo, M.-L. Climate and Local Geomorphic Interactions Drive Patterns of Riparian Forest Decline along a Mediterranean Basin River. Geomorphology 2013, 202, 101–114. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Rennenberg, H. Molecular and Physiological Responses of Trees to Waterlogging Stress. Plant Cell Environ. 2014, 37, 2245–2259. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Zhou, H.; Hao, X.; Zhu, C.; Fu, A.; Yang, Y.; Li, W. Research Advances in Plant Physiology and Ecology of Desert Riparian Forests under Drought Stress. Forests 2022, 13, 619. [Google Scholar] [CrossRef]
- Bastos, A.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Pongratz, J.; Fan, L.; Wigneron, J.-P.; Weber, U.; Reichstein, M.; Fu, Z. Direct and Seasonal Legacy Effects of the 2018 Heat Wave and Drought on European Ecosystem Productivity. Sci. Adv. 2020, 6, eaba2724. [Google Scholar] [CrossRef]
- Zscheischler, J.; Fischer, E.M. The Record-Breaking Compound Hot and Dry 2018 Growing Season in Germany. Weather Clim. Extrem. 2020, 29, 100270. [Google Scholar] [CrossRef]
- Buras, A.; Rammig, A.; Zang, C.S. Quantifying Impacts of the Drought 2018 on European Ecosystems in Comparison to 2003. Biogeosci. Discuss. 2019, 17, 1655–1672. [Google Scholar] [CrossRef] [Green Version]
- Peters, W.; Bastos, A.; Ciais, P.; Vermeulen, A. A Historical, Geographical and Ecological Perspective on the 2018 European Summer Drought. Philos. Trans. R. Soc. Lond. Ser. B 2020, 375, 20190505. [Google Scholar] [CrossRef]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.; Hauck, M.; Hajek, P. A First Assessment of the Impact of the Extreme 2018 Summer Drought on Central European Forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A Review of the Characteristics of Black Alder (Alnus glutinosa (L.) Gaertn.) and Their Implications for Silvicultural Practices. Forestry 2010, 83, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Deptuła, M.; Piernik, A.; Nienartowicz, A.; Hulisz, P.; Kamiński, D. Alnus glutinosa L. Gaertn. as Potential Tree for Brackish and Saline Habitats. Glob. Ecol. Conserv. 2020, 22, e00977. [Google Scholar] [CrossRef]
- Pogoda, P.; Ochał, W.; Orzeł, S. Modeling Diameter Distribution of Black Alder (Alnus glutinosa (L.) Gaertn.) Stands in Poland. Forests 2019, 10, 412. [Google Scholar] [CrossRef] [Green Version]
- Diagne, N.; Arumugam, K.; Ngom, M.; Nambiar-Veetil, M.; Franche, C.; Narayanan, K.K.; Laplaze, L. Use of Frankia and Actinorhizal Plants for Degraded Lands Reclamation. Biomed Res. Int. 2013, 2013, 948258. [Google Scholar] [CrossRef] [Green Version]
- Valor, T.; Camprodon, J.; Buscarini, S.; Casals, P. Drought-Induced Dieback of Riparian Black Alder as Revealed by Tree Rings and Oxygen Isotopes. For. Ecol. Manag. 2020, 478, 118500. [Google Scholar] [CrossRef]
- Granier, A.; Breda, N. Modelling Canopy Conductance and Stand Transpiration of an Oak Forest from Sap Flow Measurements. Ann. Sci. For. 1996, 53, 537–546. [Google Scholar] [CrossRef]
- Herbst, M.; Eschenbach, C.; Kappen, L. Water Use in Neighbouring Stands of Beech (Fagus Sylvatica L.) and Black Alder (Alnus glutinosa (L.) Gaertn.). Ann. For. Sci. 1999, 56, 107–120. [Google Scholar] [CrossRef]
- Eschenbach, C.; Kappen, L. Leaf Water Relations of Black Alder [Alnus glutinosa (L.) Gaertn.] Growing at Neighbouring Sites with Different Water Regimes. Trees 1999, 14, 28–38. [Google Scholar] [CrossRef]
- Chucholl, C.; Pfeiffer, M. First evidence for an established Marmorkrebs (Decapoda, Astacida, Cambaridae) population in Southwestern Germany, in syntopic occurrence with Orconectes limosus (Rafinesque, 1817). Aquat. Invasions 2010, 5, 405–412. [Google Scholar] [CrossRef]
- Gasson, P.E.; Cutler, D.F. Tree Root Plate Morphology. Arboricul. Assoc. J. 1990, 14, 193–264. [Google Scholar] [CrossRef]
- Jakubisová, M.; Jakubis, M.; Lukáčik, I. Simulation of Bank Protective Effect of Black Alder Root System (Alnus glutinosa (L.) Gaertner) According to RipRoot Model. Thaiszia J. Bot. 2014, 24, 111–124. [Google Scholar]
- Du, B.; Kreuzwieser, J.; Winkler, J.B.; Ghirardo, A.; Schnitzler, J.-P.; Ache, P.; Alfarraj, S.; Hedrich, R.; White, P.; Rennenberg, H. Physiological Responses of Date Palm (Phoenix dactylifera) Seedlings to Acute Ozone Exposure at High Temperature. Environ. Pollut. 2018, 242, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Arab, L.; Kreuzwieser, J.; Kruse, J.; Zimmer, I.; Ache, P.; Alfarraj, S.; Al-Rasheid, K.A.; Schnitzler, J.-P.; Hedrich, R.; Rennenberg, H. Acclimation to Heat and Drought—Lessons to Learn from the Date Palm (Phoenix dactylifera). Environ. Exp. Bot. 2016, 125, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Polle, A.; Chakrabarti, K.; Schürmann, W.; Rennenberg, H. Composition and Properties of Hydrogen Peroxide Decomposing Systems in Extracellular and Total Extracts from Needles of Norway Spruce (Picea abies L., Karst.). Plant Physiol. 1990, 94, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Schupp, R.; Rennenberg, H. Diurnal Changes in the Glutathione Content of Spruce Needles (Picea abies L.). Plant Sci. 1988, 57, 113–117. [Google Scholar] [CrossRef]
- Strohm, M.; Jouanin, L.; Kunert, K.J.; Pruvost, C.; Polle, A.; Foyer, C.H.; Rennenberg, H. Regulation of Glutathione Synthesis in Leaves of Transgenic Poplar (Populus tremula × P. alba) Overexpressing Glutathione Synthetase. Plant J. 1995, 7, 141–145. [Google Scholar] [CrossRef]
- Samuilov, S.; Lang, F.; Djukic, M.; Djunisijevic-Bojovic, D.; Rennenberg, H. Lead Uptake Increases Drought Tolerance of Wild Type and Transgenic Poplar (Populus tremula × P. alba) Overexpressing Gsh 1. Environ. Pollut. 2016, 216, 773–785. [Google Scholar] [CrossRef]
- Du, B.; Jansen, K.; Junker, L.V.; Eiblmeier, M.; Kreuzwieser, J.; Gessler, A.; Ensminger, I.; Rennenberg, H. Elevated Temperature Differently Affects Foliar Nitrogen Partitioning in Seedlings of Diverse Douglas Fir Provenances. Tree Physiol. 2014, 34, 1090–1101. [Google Scholar] [CrossRef]
- Carroll, N.V.; Longley, R.W.; Roe, J.H. The determination of glycogen in liverand muscle by use of anthrone reagent. J. Biol. Chem. 1956, 220, 583–593. [Google Scholar] [CrossRef]
- Peuke, A.D.; Gessler, A.; Rennenberg, H. The Effect of Drought on C and N Stable Isotopes in Different Fractions of Leaves, Stems and Roots of Sensitive and Tolerant Beech Ecotypes. Plant Cell Environ. 2006, 29, 823–835. [Google Scholar] [CrossRef]
- Hummel, J.; Strehmel, N.; Selbig, J.; Walther, D.; Kopka, J. Decision Tree Supported Substructure Prediction of Metabolites from GC-MS Profiles. Metabolomics 2010, 6, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards More Transparent and Integrative Metabolomics Analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, Superoxide Dismutase and Ascorbate-Glutathione Cycle Enzymes Confer Drought Tolerance of Amaranthus Tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [Green Version]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Ascorbate and Glutathione: The Heart of the Redox Hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Smirnof, N. The Role of Active Oxygen in the Response of Plants to Water Deficit and Desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Tausz, M.; Wonisch, A.; Peters, J.; Jiménez, M.S.; Morales, D.; Grill, D. Short-Term Changes in Free Radical Scavengers and Chloroplast Pigments in Pinus canariensis Needles as Affected by Mild Drought Stress. J. Plant Physiol. 2001, 158, 213–219. [Google Scholar] [CrossRef]
- Tausz, M.; Šircelj, H.; Grill, D. The glutathione system as a stress marker in plant ecophysiology: Is a stress-response concept valid? J. Exp. Bot. 2004, 55, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- García-Plazaola, J.I.; Becerril, J.M. Effects of drought on photoprotective mechanisms in European beech (Fagus sylvatica L.) seedlings from different provenances. Trees 2000, 14, 485–490. [Google Scholar] [CrossRef]
- Dusart, N.; Gérard, J.; Le Thiec, D.; Collignon, C.; Jolivet, Y.; Vaultier, M.N. Integrated analysis of the detoxification responses of two Euramerican poplar genotypes exposed to ozone and water deficit: Focus on the ascorbate-glutathione cycle. Sci. Total Environ. 2019, 651, 2365–2379. [Google Scholar] [CrossRef] [PubMed]
- Morell, S.; Follmann, H.; De Tullio, M.; Häberlein, I. Dehydroascorbate and Dehydroascorbate Reductase Are Phantom Indicators of Oxidative Stress in Plants. FEBS Lett. 1997, 414, 567–570. [Google Scholar] [CrossRef] [Green Version]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Morishima, I.; Shibahara, T.; Inanaga, S.; Tanaka, K. Enhanced Tolerance to Ozone and Drought Stresses in Transgenic Tobacco Overexpressing Dehydroascorbate Reductase in Cytosol. Physiol. Plant. 2006, 127, 57–65. [Google Scholar] [CrossRef]
- Sorkheh, K.; Shiran, B.; Rouhi, V.; Khodambashi, M.; Sofo, A. Regulation of the Ascorbate-Glutathione Cycle in Wild Almond during Drought Stress. Russ. J. Plant Physiol. 2011, 58, 76–84. [Google Scholar] [CrossRef]
- Wang, W.; Peng, C.; Kneeshaw, D.D.; Larocque, G.R.; Luo, Z. Drought-Induced Tree Mortality: Ecological Consequences, Causes, and Modeling. Environ. Rev. 2012, 20, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Gao, Q.; Xu, H.-Y.; Ma, F.; Zhao, C.-M.; Liu, J.-Q. Physiological Responses to Gradual Drought Stress in the Diploid Hybrid Pinus densata and Its Two Parental Species. Trees 2009, 23, 717–728. [Google Scholar] [CrossRef]
- Horemans, N.; Foyer, C.H.; Asard, H. Transport and Action of Ascorbate at the Plant Plasma Membrane. Trends Plant Sci. 2000, 5, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. The Water–Water Cycle as Alternative Photon and Electron Sinks. Philos. Trans. R. Soc. Lond. Ser. B 2000, 355, 1419–1431. [Google Scholar] [CrossRef] [Green Version]
- Terai, Y.; Ueno, H.; Ogawa, T.; Sawa, Y.; Miyagi, A.; Kawai-Yamada, M.; Ishikawa, T.; Maruta, T. Dehydroascorbate Reductases and Glutathione Set a Threshold for High-Light–Induced Ascorbate Accumulation. Plant Physiol. 2020, 183, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of Osmoprotectants in Improving Salinity and Drought Tolerance in Plants: A Review. Rev. Environ. Sci. Bio/Technol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Stagos, D. Antioxidant Activity of Polyphenolic Plant Extracts. Antioxidants 2020, 9, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, B.; Jansen, K.; Kleiber, A.; Eiblmeier, M.; Kammerer, B.; Ensminger, I.; Gessler, A.; Rennenberg, H.; Kreuzwieser, J. A Coastal and an Interior Douglas Fir Provenance Exhibit Different Metabolic Strategies to Deal with Drought Stress. Tree Physiol. 2016, 36, 148–163. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Kruse, J.; Winkler, J.B.; Alfarraj, S.; Albasher, G.; Schnitzler, J.-P.; Ache, P.; Hedrich, R.; Rennenberg, H. Metabolic Responses of Date Palm (Phoenix dactylifera L.) Leaves to Drought Differ in Summer and Winter Climate. Tree Physiol. 2021, 41, 1685–1700. [Google Scholar] [CrossRef] [PubMed]
- Merewitz, E. Chemical Priming-Induced Drought Stress Tolerance in Plants. In Drought Stress Tolerance in Plants; Springer International Publishing: Cham, Switzerland, 2016; pp. 77–103. [Google Scholar]
- Duan, S.; Kwon, S.J.; Lim, Y.J.; Gil, C.S.; Jin, C.; Eom, S.H. L-3, 4-Dihydroxyphenylalanine Accumulation in Faba Bean (Vicia faba L.) Tissues during Different Growth Stages. Agronomy 2021, 11, 502. [Google Scholar] [CrossRef]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and Raffinose Constitute a Novel Function to Protect Plants from Oxidative Damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [Green Version]
- Taji, T.; Ohsumi, C.; Iuchi, S.; Seki, M.; Kasuga, M.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Important roles of drought- and cold-inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana. Plant J. 2002, 29, 417–426. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Y.; Liu, Y.; Li, X.; Hao, G.; Han, Q.; Dirk, L.M.A.; Downie, A.B.; Ruan, Y.-L.; Wang, J.; et al. Raffinose synthase enhances drought tolerance through raffinose synthesis or galactinol hydrolysis in maize and Arabidopsis plants. J. Biol. Chem. 2020, 295, 8064–8077. [Google Scholar] [CrossRef] [PubMed]
- Riikonen, J.; Kontunen-Soppela, S.; Vapaavuori, E.; Tervahauta, A.; Tuomainen, M.; Oksanen, E. Carbohydrate concentrations and freezing stress resistance of silver birch buds grown under elevated temperature and ozone. Tree Physiol. 2013, 33, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valluru, R.; Van den Ende, W. Myo-Inositol and beyond–Emerging Networks under Stress. Plant Sci. 2011, 181, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Návarová, H.; Bernsdorff, F.; Döring, A.-C.; Zeier, J. Pipecolic Acid, an Endogenous Mediator of Defense Amplification and Priming, Is a Critical Regulator of Inducible Plant Immunity. Plant Cell 2012, 24, 5123–5141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranda, I.; Sánchez-Gómez, D.; de Miguel, M.; Mancha, J.A.; Guevara, M.A.; Cadahía, E.; de Simón, M.B.F. Fagus sylvatica L. provenances maintain different leaf metabolic profiles and functional response. Acta Oecol. 2017, 82, 1–9. [Google Scholar] [CrossRef]
- Sebastiana, M.; Duarte, B.; Monteiro, F.; Malhó, R.; Caçador, I.; Matos, A.R. The leaf lipid composition of ectomycorrhizal oak plants shows a drought-tolerance signature. Plant Physiol. Biochem. 2019, 144, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Kolb, R.M.; Rawyler, A.; Brändle, R. Parameters affecting the early seedling development of four neotropical trees under oxygen deprivation stress. Ann. Bot. 2002, 89, 551–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowakowska, J.A.; Stocki, M.; Stocka, N.; Ślusarski, S.; Tkaczyk, M.; Caetano, J.M.; Tulik, M.; Hsiang, T.; Oszako, T. Interactions between Phytophthora cactorum, Armillaria gallica and Betula pendula Roth. Seedlings Subjected to Defoliation. Forests 2020, 11, 1107. [Google Scholar] [CrossRef]
- Okazaki, Y.; Saito, K. Roles of Lipids as Signaling Molecules and Mitigators during Stress Response in Plants. Plant J. 2014, 79, 584–596. [Google Scholar] [CrossRef]
- Malcheska, F.; Ahmad, A.; Batool, S.; Müller, H.M.; Ludwigmüller, J.; Kreuzwieser, J.; Randewig, D.; Hänsch, R.; Mendel, R.R.; Hell, R. Drought Enhanced Xylem Sap Sulfate Closes Stomata by Affecting ALMT12 and Guard Cell ABA Synthesis. Plant Physiol. 2017, 174, 798. [Google Scholar] [CrossRef] [Green Version]
- Batool, S.; Uslu, V.V.; Rajab, H.; Ahmad, N.; Waadt, R.; Geiger, D.; Malagoli, M.; Xiang, C.-B.; Hedrich, R.; Rennenberg, H. Sulfate Is Incorporated into Cysteine to Trigger ABA Production and Stomatal Closure. Plant Cell 2018, 30, 2973–2987. [Google Scholar] [CrossRef] [PubMed]
- Ernst, L.; Goodger, J.Q.; Alvarez, S.; Marsh, E.L.; Berla, B.; Lockhart, E.; Jung, J.; Li, P.; Bohnert, H.J.; Schachtman, D.P. Sulphate as a Xylem-Borne Chemical Signal Precedes the Expression of ABA Biosynthetic Genes in Maize Roots. J. Exp. Bot. 2010, 61, 3395–3405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.X.; Wirtz, M.; Phua, S.Y.; Estavillo, G.M.; Pogson, B.J. Balancing Metabolites in Drought: The Sulfur Assimilation Conundrum. Trends Plant Sci. 2013, 18, 18–29. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.G. Mechanisms Linking Drought, Hydraulics, Carbon Metabolism, and Vegetation Mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Liao, Z.; Liu, L.; Du, B. Effects of Extreme Drought and Heat Events on Leaf Metabolome of Black Alder (Alnus glutinosa L.) Growing at Neighboring Sites with Different Water Availability. Forests 2023, 14, 151. https://doi.org/10.3390/f14010151
Zhu L, Liao Z, Liu L, Du B. Effects of Extreme Drought and Heat Events on Leaf Metabolome of Black Alder (Alnus glutinosa L.) Growing at Neighboring Sites with Different Water Availability. Forests. 2023; 14(1):151. https://doi.org/10.3390/f14010151
Chicago/Turabian StyleZhu, Lijun, Zhengqiao Liao, Lei Liu, and Baoguo Du. 2023. "Effects of Extreme Drought and Heat Events on Leaf Metabolome of Black Alder (Alnus glutinosa L.) Growing at Neighboring Sites with Different Water Availability" Forests 14, no. 1: 151. https://doi.org/10.3390/f14010151