
Root-Growth-Related MaTCP Transcription Factors Responsive to Drought Stress in Mulberry
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Conditions
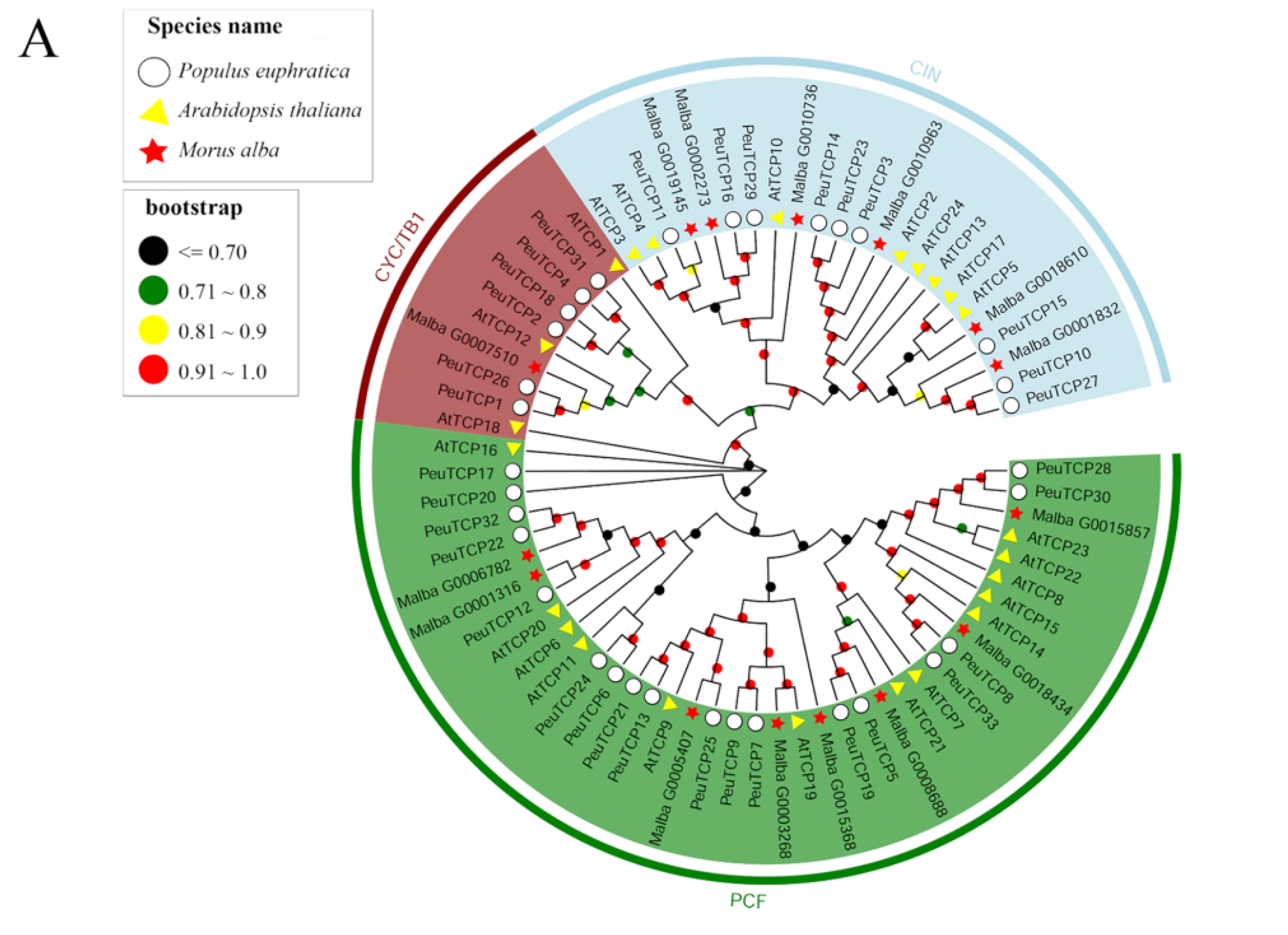
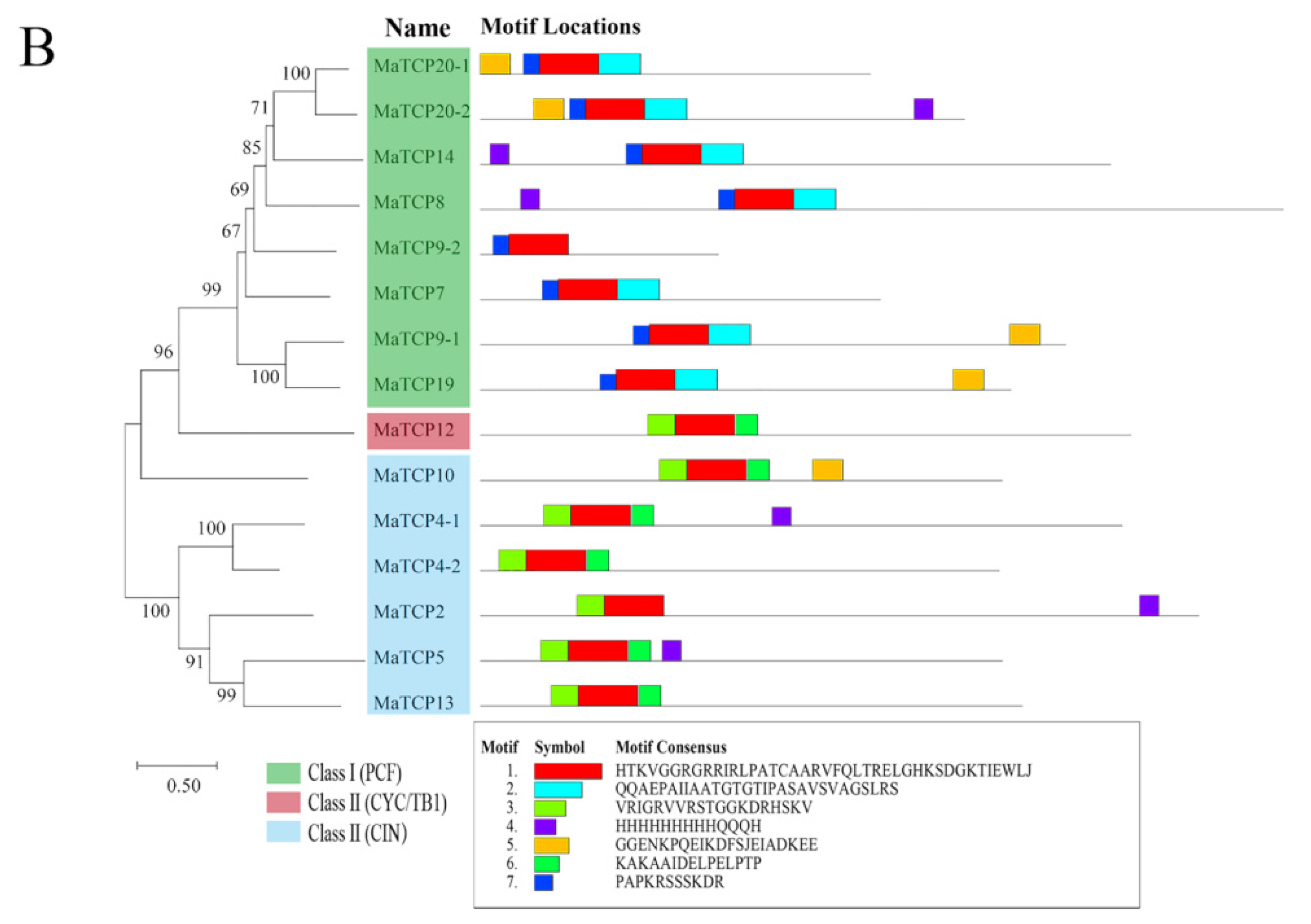
2.2. Phylogenetic and Comparative Sequence Analyses of the TCP Family
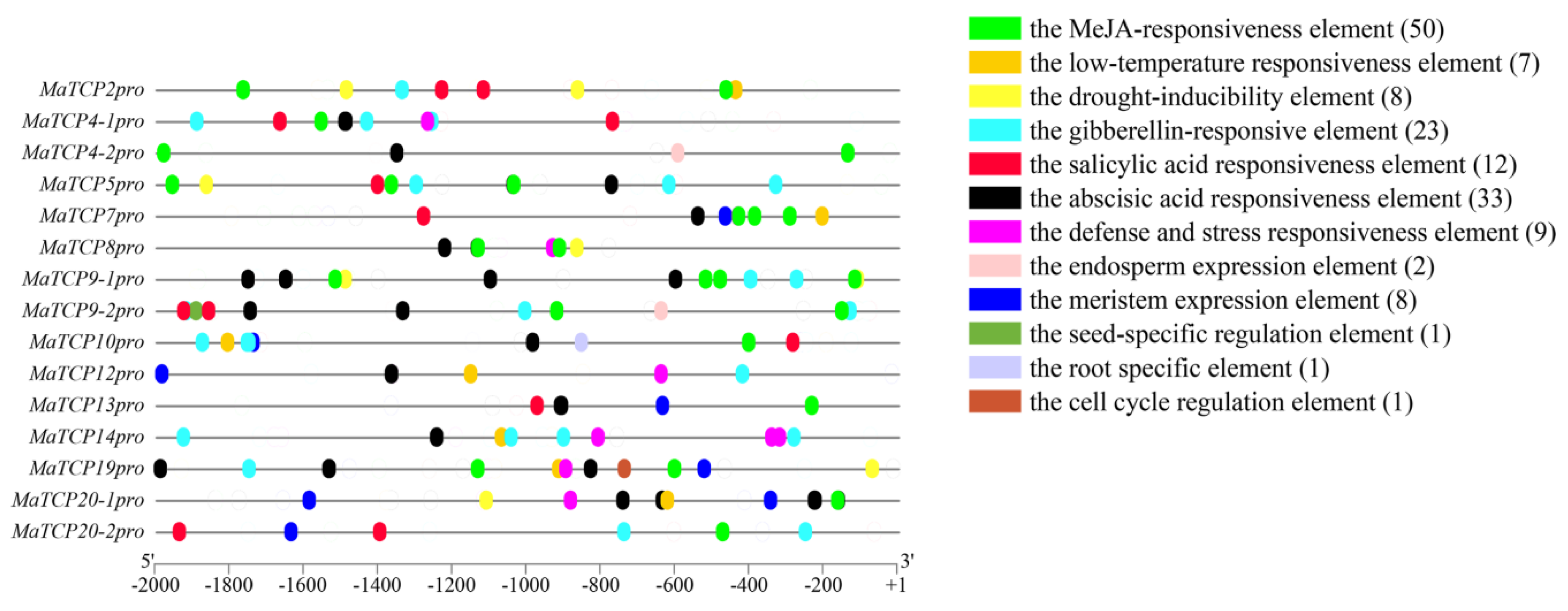
2.3. Promoter Element Analyses
2.4. RNA Extraction, Gene Cloning, and Quantitative Real-Time PCR (RT-qPCR) Analysis
2.5. Subcellular Localization
2.6. Stress Treatment
2.7. Data Processing and Statistical Analysis
3. Results
3.1. Characteristics of MaTCP Transcription Factors in Mulberry
3.2. The Promoter Analyses of MaTCP Genes
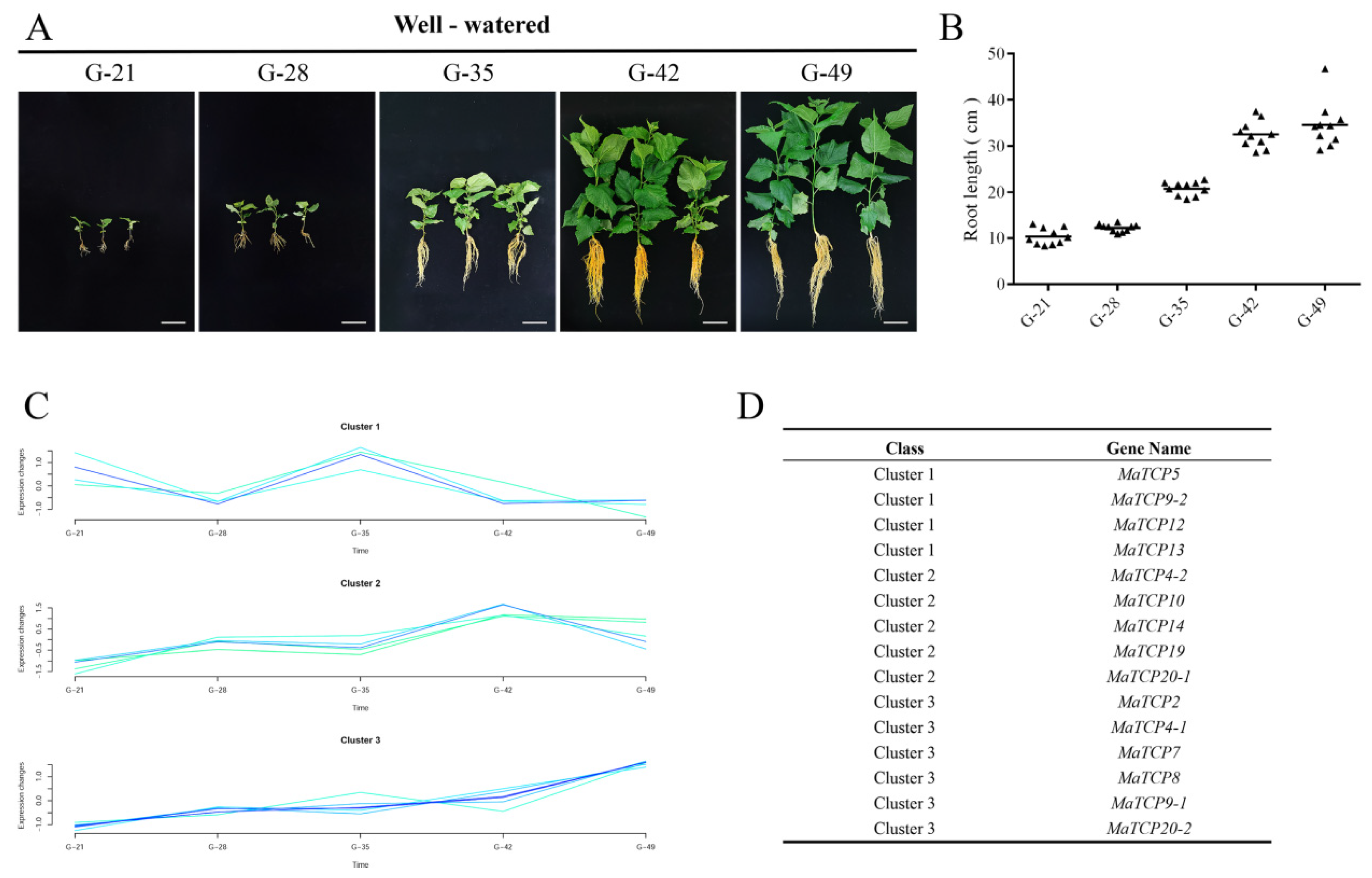
3.3. Expression Patterns of MaTCP Genes during Mulberry Seedling Growth
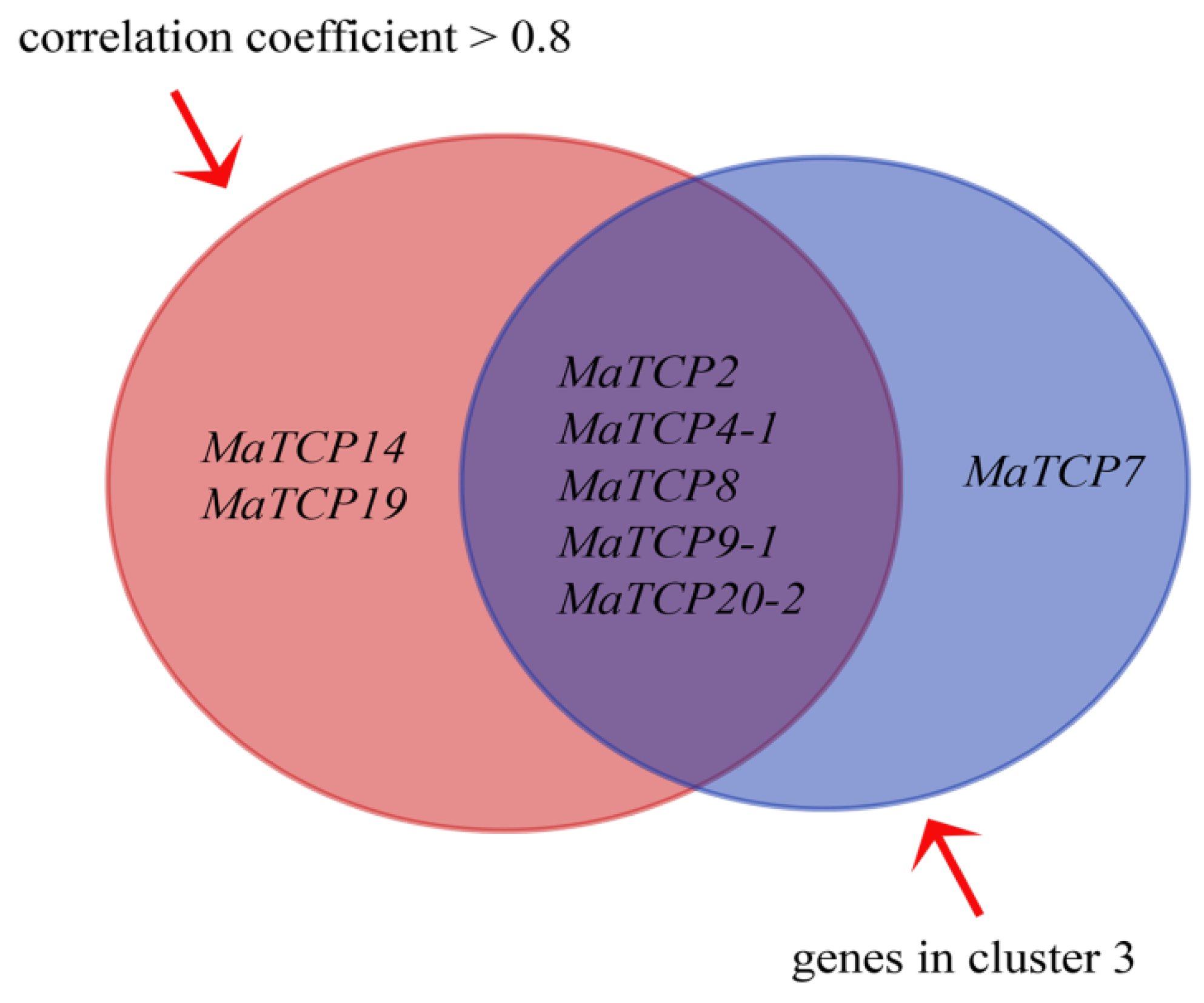
3.4. Association of Five MaTCP Genes with Root Development
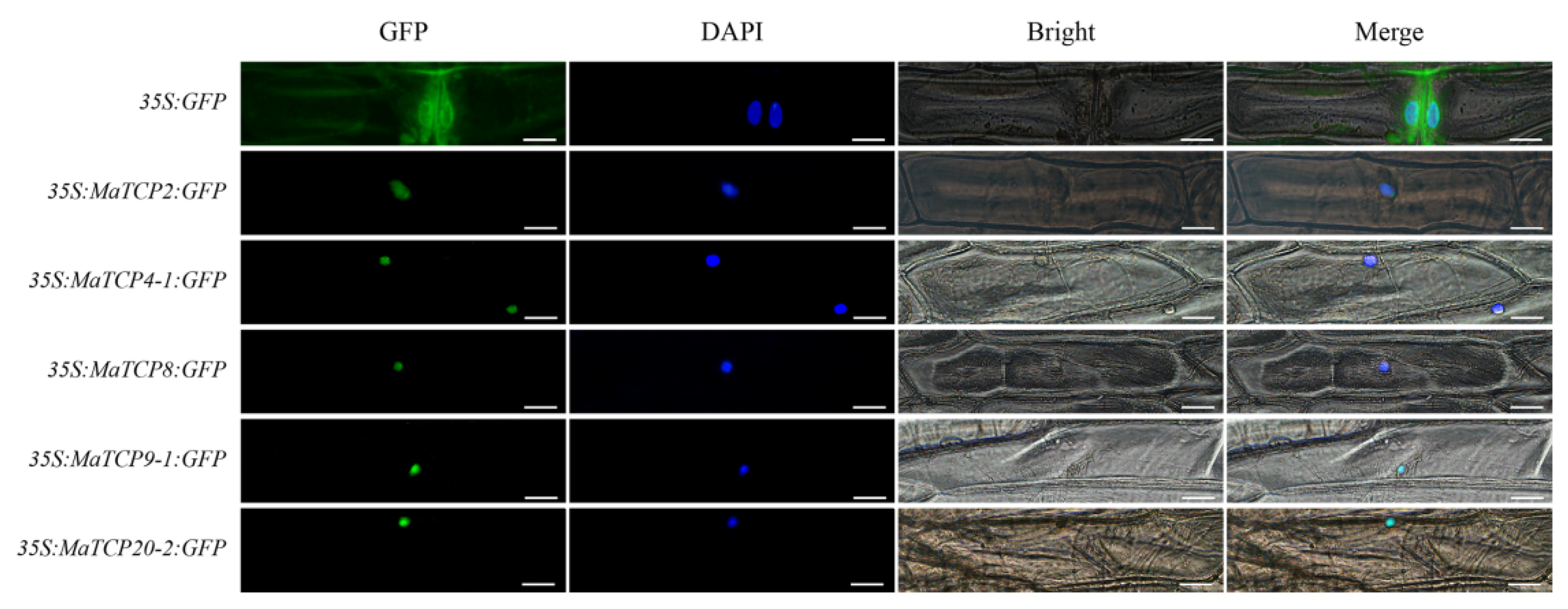
3.5. Subcellular Localization Analyses of Five MaTCP Proteins
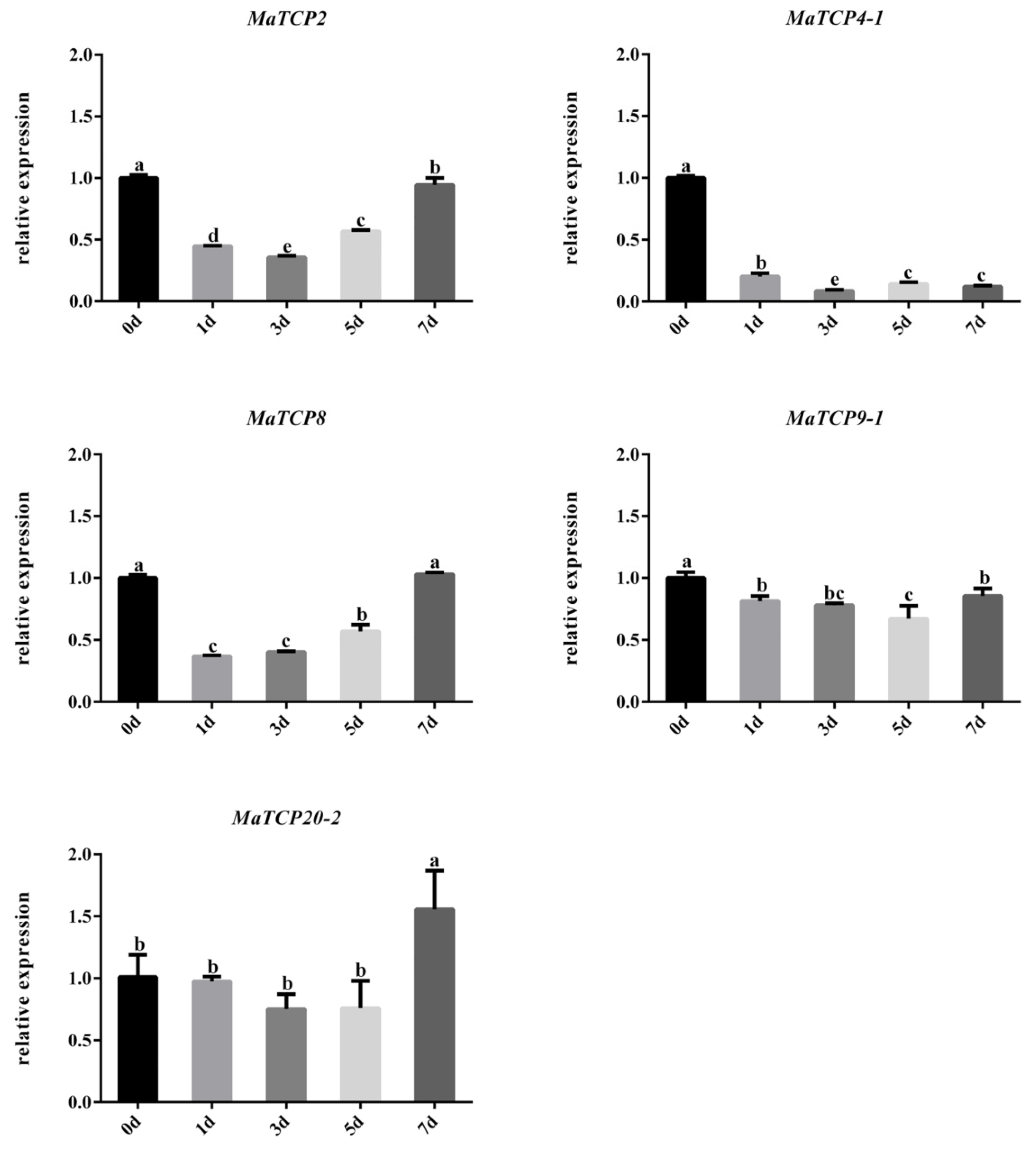
3.6. Expression of Five MaTCP Genes under Drought Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mega, R.; Abe, F.; Kim, J.S.; Tsuboi, Y.; Tanaka, K.; Kobayashi, H.; Sakata, Y.; Hanada, K.; Tsujimoto, H.; Kikuchi, J.; et al. Tuning water-use efficiency and drought tolerance in wheat using abscisic acid receptors. Nat. Plants 2019, 5, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.; Xue, W.Y.; Xiong, L.Z.; Yu, X.Q.; Luo, L.J.; Cui, K.H.; Jin, D.M.; Xing, Y.Z.; Zhang, Q.F. Genetic basis of drought resistance at reproductive stage in rice: Separation of drought tolerance from drought avoidance. Genetics 2006, 172, 1213–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.J.; Xiong, L.Z. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; de Groot, S.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, Y.H.; Chen, Q.J.; Bai, G.H.; Gao, W.W.; Qu, Y.Y.; Ni, Z.Y. Heterologous expression of GbTCP4, a class II TCP transcription factor, regulates trichome formation and root hair development in Arabidopsis. Genes 2019, 10, 726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yu, Y.H.; Wang, J.D.; Chen, Q.J.; Ni, Z.Y. Heterologous overexpression of the GbTCP5 gene increased root hair length, root hair and stem trichome density, and lignin content in transgenic Arabidopsis. Gene 2020, 758, 144954. [Google Scholar] [CrossRef]
- Resentini, F.; Felipo-Benavent, A.; Colombo, L.; Blazquez, M.A.; Alabadi, D.; Masiero, S. TCP14 and TCP15 mediate the promotion of seed germination by gibberellins in Arabidopsis thaliana. Mol. Plant. 2015, 8, 482–485. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.A.; Sun, Y.; Blair, P.B.; Mukhtar, M.S. TCP three-way handshake: Linking developmental processes with plant immunity. Trends Plant Sci. 2015, 20, 238–245. [Google Scholar] [CrossRef]
- Danisman, S. TCP transcription factors at the interface between environmental challenges and the plant’s growth responses. Front. Plant Sci. 2016, 7, 1930. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.J.; Zheng, Y.Q.; Lu, W.; Li, J.; Duan, Y.J.; Zhang, S.; Wang, Y.P. Roles of miR319-regulated TCPs in plant development and response to abiotic stress. Crop J. 2021, 9, 17–28. [Google Scholar] [CrossRef]
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP domain: A motif found in proteins regulating plant growth and development. Plant J. 1999, 18, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Martin-Trillo, M.; Cubas, P. TCP genes: A family snapshot ten years later. Trends Plant Sci. 2010, 15, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Manassero, N.G.U.; Viola, I.L.; Welchen, E.; Gonzalez, D.H. TCP transcription factors: Architectures of plant form. Biomol. Concepts 2013, 4, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, P.; Das Gupta, M.; Joseph, A.P.; Chatterjee, N.; Srinivasan, N.; Nath, U. Identification of specific DNA binding residues in the TCP family of transcription factors in Arabidopsis. Plant Cell 2010, 22, 1174–1189. [Google Scholar] [CrossRef] [Green Version]
- Horn, S.; Pabon-Mora, N.; Theuss, V.S.; Busch, A.; Zachgo, S. Analysis of the CYC/TB1 class of TCP transcription factors in basal angiosperms and magnoliids. Plant J. 2015, 81, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, M.; Cubas, P. TCP factors: New kids on the signaling block. Curr. Opin. Plant Biol. 2016, 33, 33–41. [Google Scholar] [CrossRef]
- Kieffer, M.; Master, V.; Waites, R.; Davies, B. TCP14 and TCP15 affect internode length and leaf shape in Arabidopsis. Plant J. 2011, 68, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.C.; Chen, L.L.; Lu, Y.R.; Wu, Y.B.; Dumenil, J.; Zhu, Z.G.; Bevan, M.W.; Li, Y.H. The ubiquitin receptors DA1, DAR1, and DAR2 redundantly regulate endoreduplication by modulating the stability of TCP14/15 in Arabidopsis. Plant Cell 2015, 27, 649–662. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Tu, L.L.; Hu, H.Y.; Tan, J.F.; Deng, F.L.; Tang, W.X.; Nie, Y.C.; Zhang, X.L. GbTCP, a cotton TCP transcription factor, confers fibre elongation and root hair development by a complex regulating system. J. Exp. Bot. 2012, 63, 6267–6281. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, P.; Tyagi, A.K. OsTCP19 influences developmental and abiotic stress signaling by modulating ABI4-mediated pathways. Sci. Rep. 2015, 5, 9998. [Google Scholar] [CrossRef]
- Ori, N.; Cohen, A.R.; Etzioni, A.; Brand, A.; Yanai, O.; Shleizer, S.; Menda, N.; Amsellem, Z.; Efroni, I.; Pekker, I.; et al. Regulation of LANCEOLATE by miR319 is required for compound-leaf development in tomato. Nat. Genet. 2007, 39, 787–791. [Google Scholar] [CrossRef]
- Nag, A.; King, S.; Jack, T. miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 22534–22539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Li, D.Y.; Li, Z.G.; Hu, Q.; Yang, C.H.; Zhu, L.H.; Luo, H. Constitutive expression of a miR319 gene alters plant development and enhances salt and drought tolerance in transgenic creeping bentgrass. Plant Physiol. 2013, 161, 1375–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, N.J.; Zhang, C.; Qi, X.W.; Zhao, S.C.; Tao, Y.; Yang, G.J.; Lee, T.H.; Wang, X.Y.; Cai, Q.L.; Li, D.; et al. Draft genome sequence of the mulberry tree Morus notabilis. Nat. Commun. 2013, 4, 2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.-P.; Ou, T.-T.; Wang, C.-J. Mulberry (sang shen zǐ) and its bioactive compounds, the chemoprevention effects and molecular mechanisms in vitro and in vivo. J. Tradit. Complement. Med. 2013, 3, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Rohela, G.K.; Shukla, P.; Muttanna; Kumar, R.; Chowdhury, S.R. Mulberry (Morus spp.): An ideal plant for sustainable development. Trees For. People 2020, 2, 100011. [Google Scholar] [CrossRef]
- Liu, Y.; Willison, J.H.M. Prospects for cultivating white mulberry (Morus alba) in the drawdown zone of the Three Gorges Reservoir, China. Environ. Sci. Pollut. Res. 2013, 20, 7142–7151. [Google Scholar] [CrossRef]
- Ma, X.D.; Ma, J.C.; Fan, D.; Li, C.F.; Jiang, Y.Z.; Luo, K.M. Genome-wide identification of TCP family transcription factors from Populus euphratica and their involvement in leaf shape regulation. Sci. Rep. 2016, 6, 32795. [Google Scholar] [CrossRef] [Green Version]
- Jiao, F.; Luo, R.S.; Dai, X.L.; Liu, H.; Yu, G.; Han, S.H.; Lu, X.; Su, C.; Chen, Q.; Song, Q.X.; et al. Chromosome-level reference genome and population genomic analysis provide insights into the evolution and improvement of domesticated mulberry (Morus alba). Mol. Plant. 2020, 13, 1001–1012. [Google Scholar] [CrossRef]
- Mulder, N.J.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Binns, D.; Bradley, P.; Bork, P.; Bucher, P.; Cerutti, L.; et al. InterPro, progress and status in 2005. Nucleic Acids Res. 2005, 33, D201–D205. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.W.; Shuai, Q.; Chen, H.; Fan, L.; Zeng, Q.W.; He, N.J. Cloning and expression analyses of the anthocyanin biosynthetic genes in mulberry plants. Mol. Genet. Genom. 2014, 289, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Huang, R.Z.; Jiang, Y.B.; Jia, C.H.; Jiang, S.M.; Yan, X.P. Subcellular distribution and chemical forms of cadmium in Morus alba L. Int. J. Phytoremediat. 2018, 20, 448–453. [Google Scholar] [CrossRef]
- Feng, D.L.; Huang, X.H.; Liu, Y.; Willison, J.H.M. Growth and changes of endogenous hormones of mulberry roots in a simulated rocky desertification area. Environ. Sci. Pollut. Res. 2016, 23, 11171–11180. [Google Scholar] [CrossRef]
- Huang, X.H.; Liu, Y.; Li, J.X.; Xiong, X.Z.; Chen, Y.H.; Yin, X.H.; Feng, D.L. The response of mulberry trees after seedling hardening to summer drought in the hydro-fluctuation belt of Three Gorges Reservoir Areas. Environ. Sci. Pollut. Res. 2013, 20, 7103–7111. [Google Scholar] [CrossRef]
- Checker, V.G.; Saeed, B.; Khurana, P. Analysis of expressed sequence tags from mulberry (Morus indica) roots and implications for comparative transcriptomics and marker identification. Tree Genet. Genomes 2012, 8, 1437–1450. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TFID | Common Name | Type | Conserved Domain (aa) | Chromosome Location | CDS Length (bp) | Protein Length (aa) | Predicted Protein Localization |
|---|---|---|---|---|---|---|---|
| M.alba_G0010963 | MaTCP2 | CIN | 78–236 | Chr04: 10504440..10506373 | 1491 | 496 | Nucleus |
| M.alba_G0019145 | MaTCP4-1 | CIN | 55–170 | Chr09: 2738157..2740500 | 1332 | 443 | Nucleus |
| M.alba_G0002273 | MaTCP4-2 | CIN | 24–117 | Chr10: 10539871..10541516 | 1077 | 358 | Nucleus |
| M.alba_G0018610 | MaTCP5 | CIN | 54–147 | Chr08: 11690488..11691912 | 1083 | 360 | Nucleus |
| M.alba_G0008688 | MaTCP7 | PCF | 51–128 | Chr02: 11919887..11922133 | 831 | 276 | Nucleus |
| M.alba_G0015857 | MaTCP8 | PCF | 172–336 | Chr06: 21186236..21188526 | 1665 | 554 | Nucleus |
| M.alba_G0005407 | MaTCP9-1 | PCF | 113–190 | Chr12: 15460670..15462253 | 1215 | 404 | Nucleus |
| M.alba_G0015368 | MaTCP9-2 | PCF | 17–144 | Chr06: 17004501..17004995 | 495 | 164 | Nucleus |
| M.alba_G0010736 | MaTCP10 | CIN | 135–219 | Chr04: 6280140..6281898 | 1083 | 360 | Nucleus |
| M.alba_G0007510 | MaTCP12 | CYC/TB1 | 126–257 | Chr14: 7692279..7693628 | 1350 | 449 | Nucleus |
| M.alba_G0001832 | MaTCP13 | CIN | 61–207 | Chr10: 4961787..4963406 | 1125 | 374 | Nucleus |
| M.alba_G0018434 | MaTCP14 | PCF | 107–282 | Chr08: 7338007..7339840 | 1308 | 435 | Nucleus |
| M.alba_G0003268 | MaTCP19 | PCF | 91–157 | Chr11: 3914056..3915332 | 1102 | 366 | Nucleus |
| M.alba_G0006782 | MaTCP20-1 | PCF | 37–125 | Chr14: 728387..729849 | 810 | 269 | Nucleus |
| M.alba_G0001316 | MaTCP20-2 | PCF | 69–156 | Chr10: 593920..595287 | 1005 | 334 | Nucleus |
| Root Length | MaTCP2 | MaTCP4-1 | MaTCP4-2 | MaTCP5 | MaTCP7 | MaTCP8 | MaTCP9-1 | MaTCP9-2 | MaTCP10 | MaTCP12 | MaTCP13 | MaTCP14 | MaTCP19 | MaTCP20-1 | MaTCP20-2 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root Length | 1.0000 | |||||||||||||||
| MaTCP2 | 0.8743 | 1.0000 | ||||||||||||||
| MaTCP4-1 | 0.8234 | 0.9902 | 1.0000 | |||||||||||||
| MaTCP4-2 | 0.6699 | 0.3515 | 0.2348 | 1.0000 | ||||||||||||
| MaTCP5 | −0.4257 | −0.4922 | −0.3966 | −0.6194 | 1.0000 | |||||||||||
| MaTCP7 | 0.6955 | 0.8982 | 0.9492 | 0.0031 | −0.1111 | 1.0000 | ||||||||||
| MaTCP8 | 0.8761 | 0.9808 | 0.9472 | 0.4683 | −0.6480 | 0.7989 | 1.0000 | |||||||||
| MaTCP9-1 | 0.8481 | 0.9968 | 0.9875 | 0.3488 | −0.5323 | 0.8911 | 0.9839 | 1.0000 | ||||||||
| MaTCP9-2 | −0.2840 | −0.3748 | −0.2780 | −0.4582 | 0.9480 | 0.0143 | −0.5367 | −0.4068 | 1.0000 | |||||||
| MaTCP10 | 0.5372 | 0.1679 | 0.0528 | 0.9789 | −0.5054 | −0.1547 | 0.2840 | 0.1647 | −0.3416 | 1.0000 | ||||||
| MaTCP12 | −0.6220 | −0.6888 | −0.6220 | −0.6790 | 0.8925 | −0.4070 | −0.7914 | −0.7299 | 0.7122 | −0.5611 | 1.0000 | |||||
| MaTCP13 | −0.3205 | −0.6062 | −0.5719 | −0.0533 | 0.7356 | −0.3836 | −0.6815 | −0.6290 | 0.8311 | 0.1258 | 0.5628 | 1.0000 | ||||
| MaTCP14 | 0.8765 | 0.7762 | 0.6946 | 0.8480 | −0.7511 | 0.4828 | 0.8541 | 0.7840 | −0.5664 | 0.7320 | −0.8993 | −0.4104 | 1.0000 | |||
| MaTCP19 | 0.9249 | 0.8014 | 0.7117 | 0.8032 | −0.7134 | 0.4834 | 0.8760 | 0.7908 | −0.6072 | 0.6673 | −0.7799 | −0.5066 | 0.9523 | 1.0000 | ||
| MaTCP20-1 | 0.7055 | 0.5086 | 0.4407 | 0.8764 | −0.5261 | 0.3010 | 0.5691 | 0.5223 | −0.2542 | 0.8447 | −0.7812 | 0.0261 | 0.8811 | 0.7290 | 1.0000 | |
| MaTCP20-2 | 0.9031 | 0.9684 | 0.9312 | 0.5514 | −0.6460 | 0.7850 | 0.9895 | 0.9734 | −0.4947 | 0.3818 | −0.8315 | −0.5890 | 0.9052 | 0.8945 | 0.6782 | 1.0000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, W.; He, J.; Luo, Y.; Yang, Z.; Xia, X.; Han, Y.; He, N. Root-Growth-Related MaTCP Transcription Factors Responsive to Drought Stress in Mulberry. Forests 2023, 14, 143. https://doi.org/10.3390/f14010143
Wei W, He J, Luo Y, Yang Z, Xia X, Han Y, He N. Root-Growth-Related MaTCP Transcription Factors Responsive to Drought Stress in Mulberry. Forests. 2023; 14(1):143. https://doi.org/10.3390/f14010143
Chicago/Turabian StyleWei, Wuqi, Jinzhi He, Yiwei Luo, Zhen Yang, Xiaoyu Xia, Yuanxiang Han, and Ningjia He. 2023. "Root-Growth-Related MaTCP Transcription Factors Responsive to Drought Stress in Mulberry" Forests 14, no. 1: 143. https://doi.org/10.3390/f14010143