
Moderate Nitrogen Deposition Alleviates Drought Stress of Bretschneidera sinensis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Fertilizer
2.2. Experimental Treatments
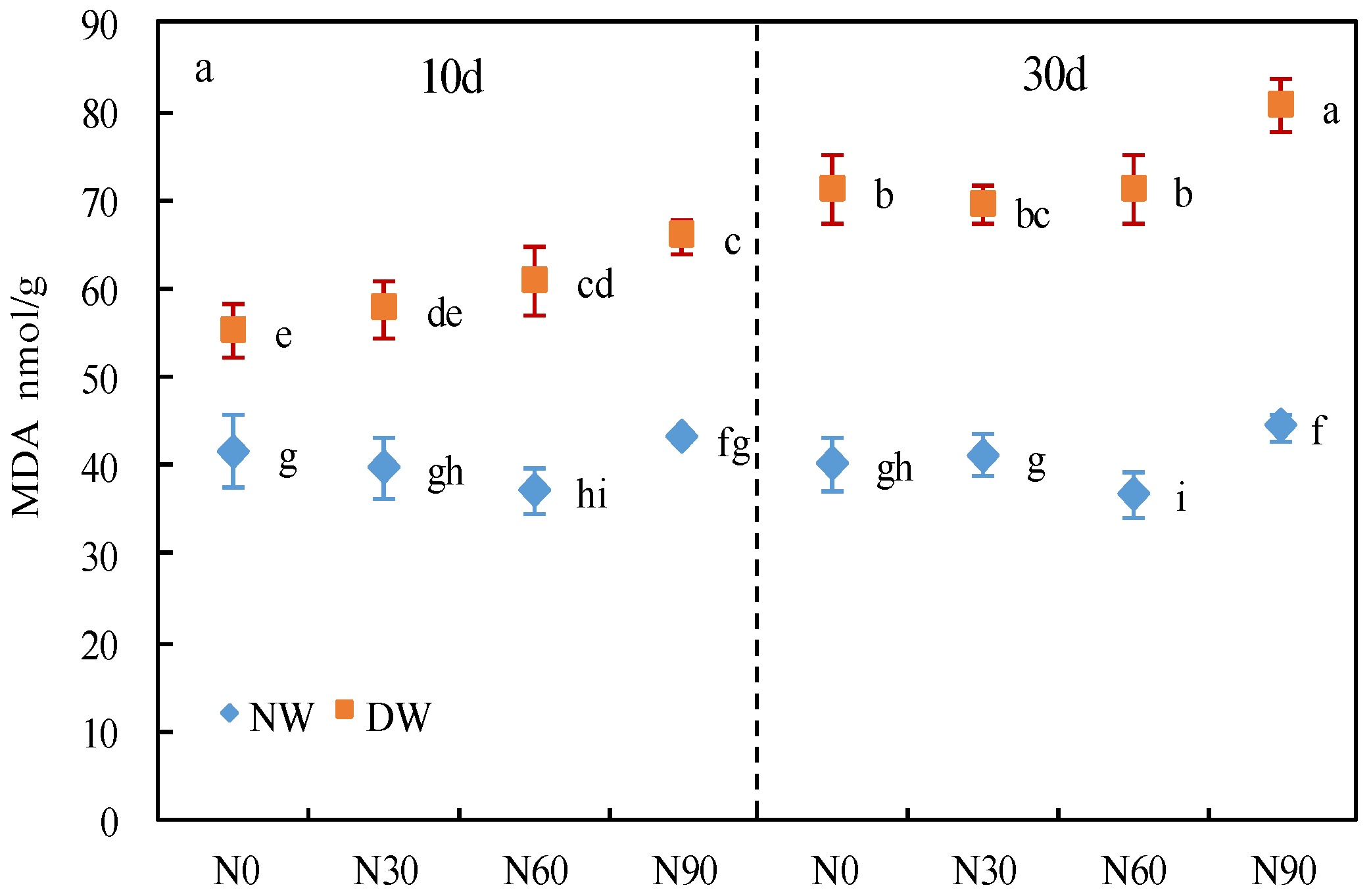
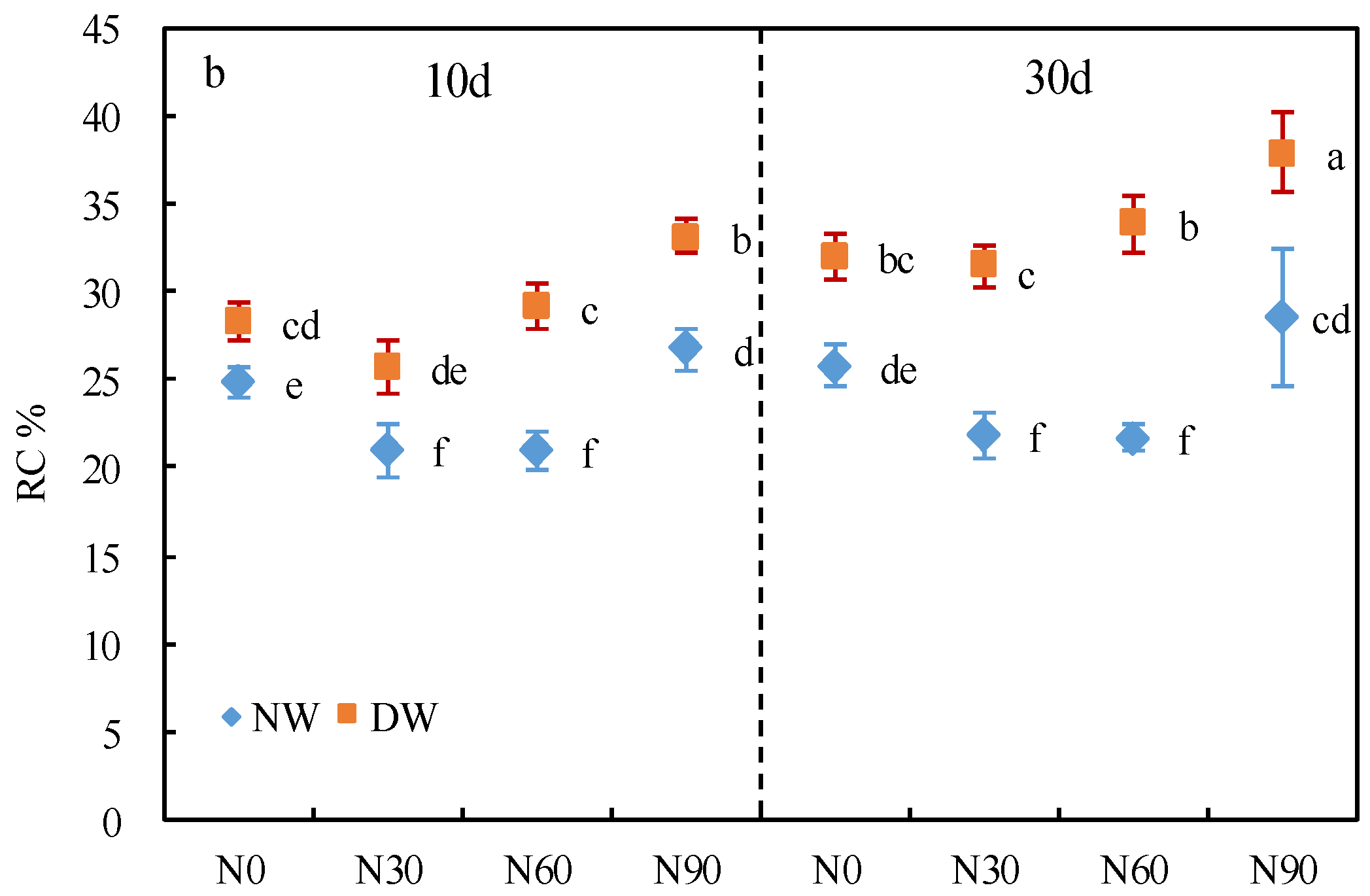
2.3. Determination of Cell Membrane Permeability
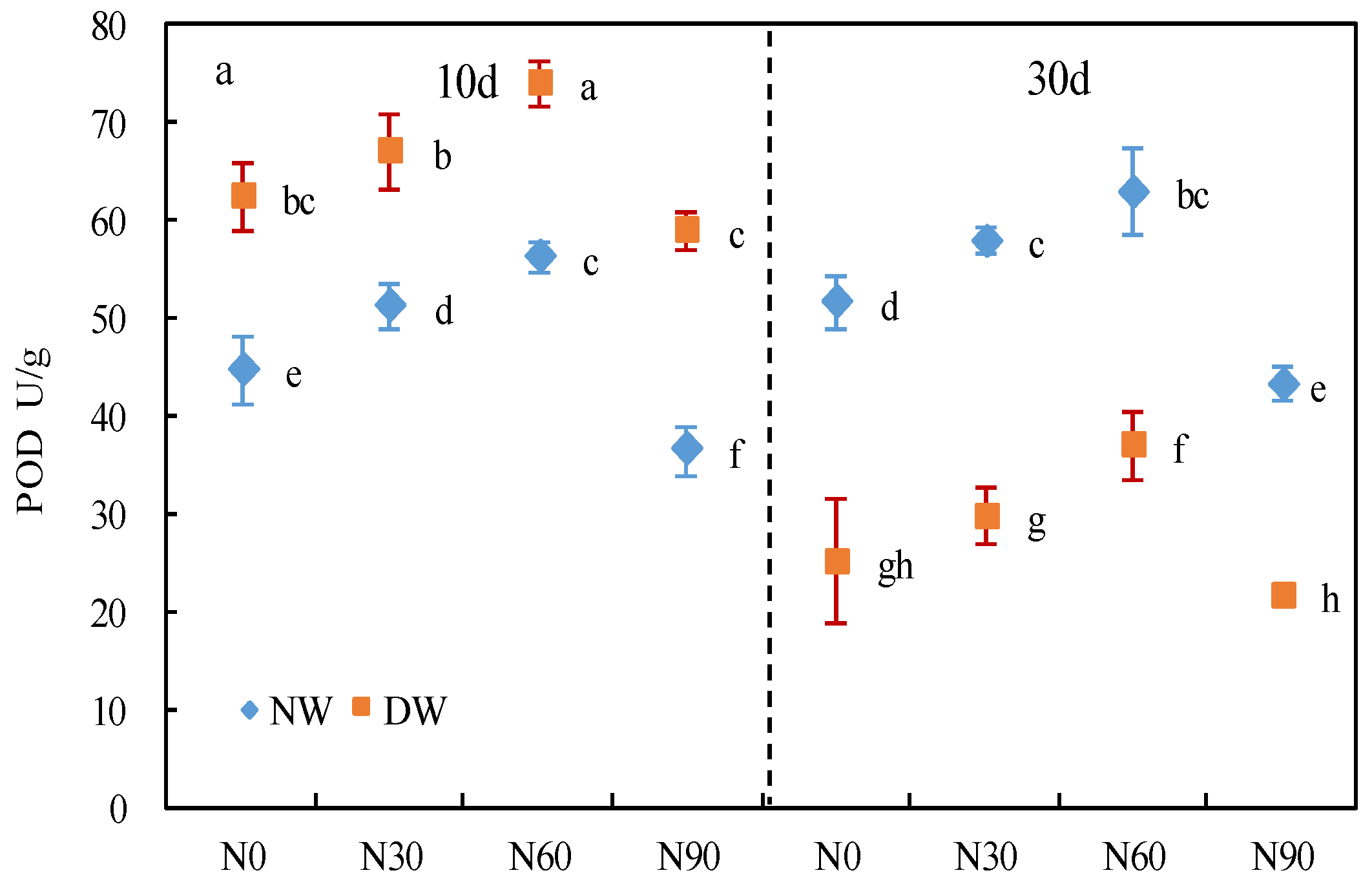
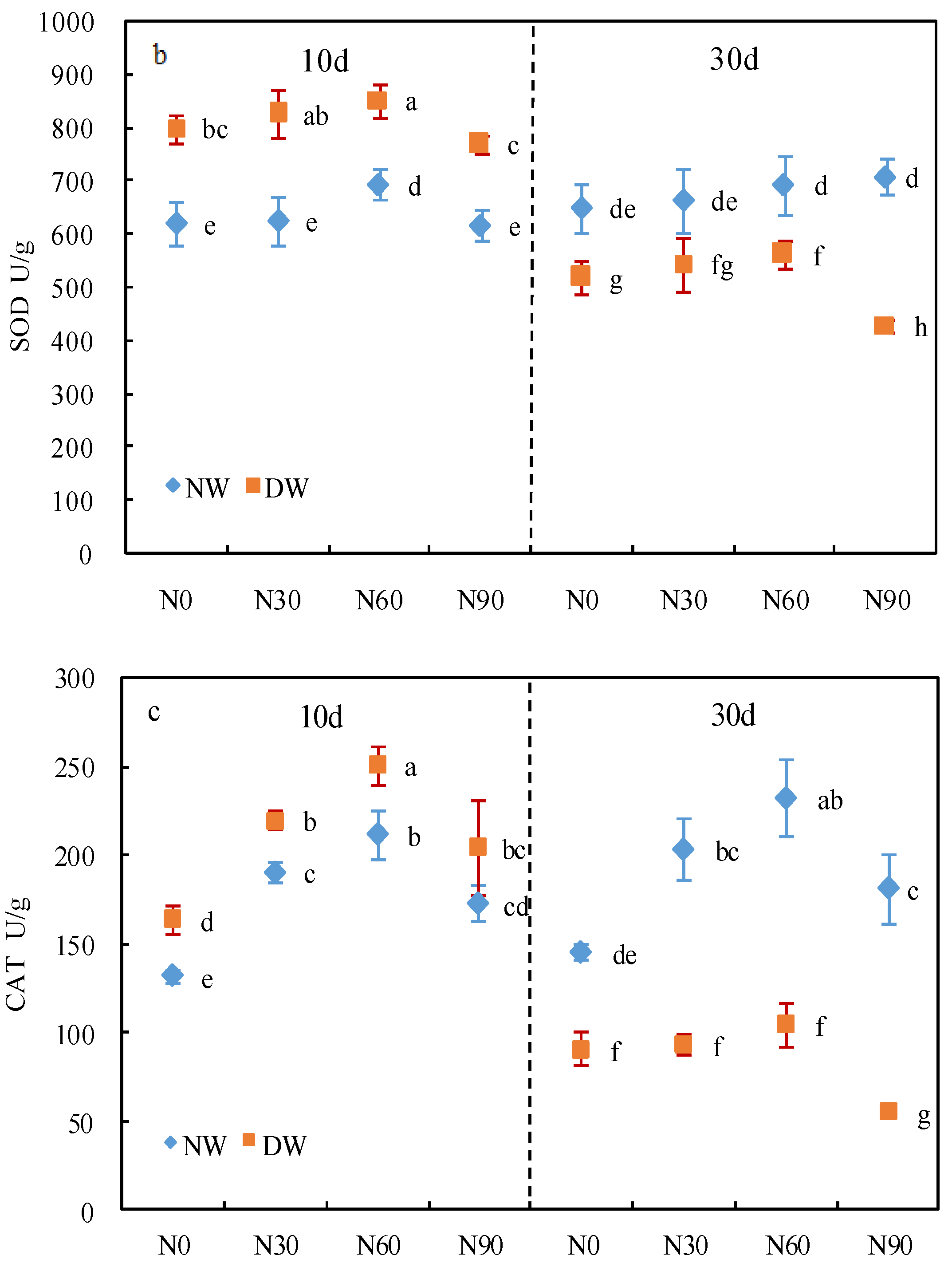
2.4. Determination of Protective Enzyme Activity
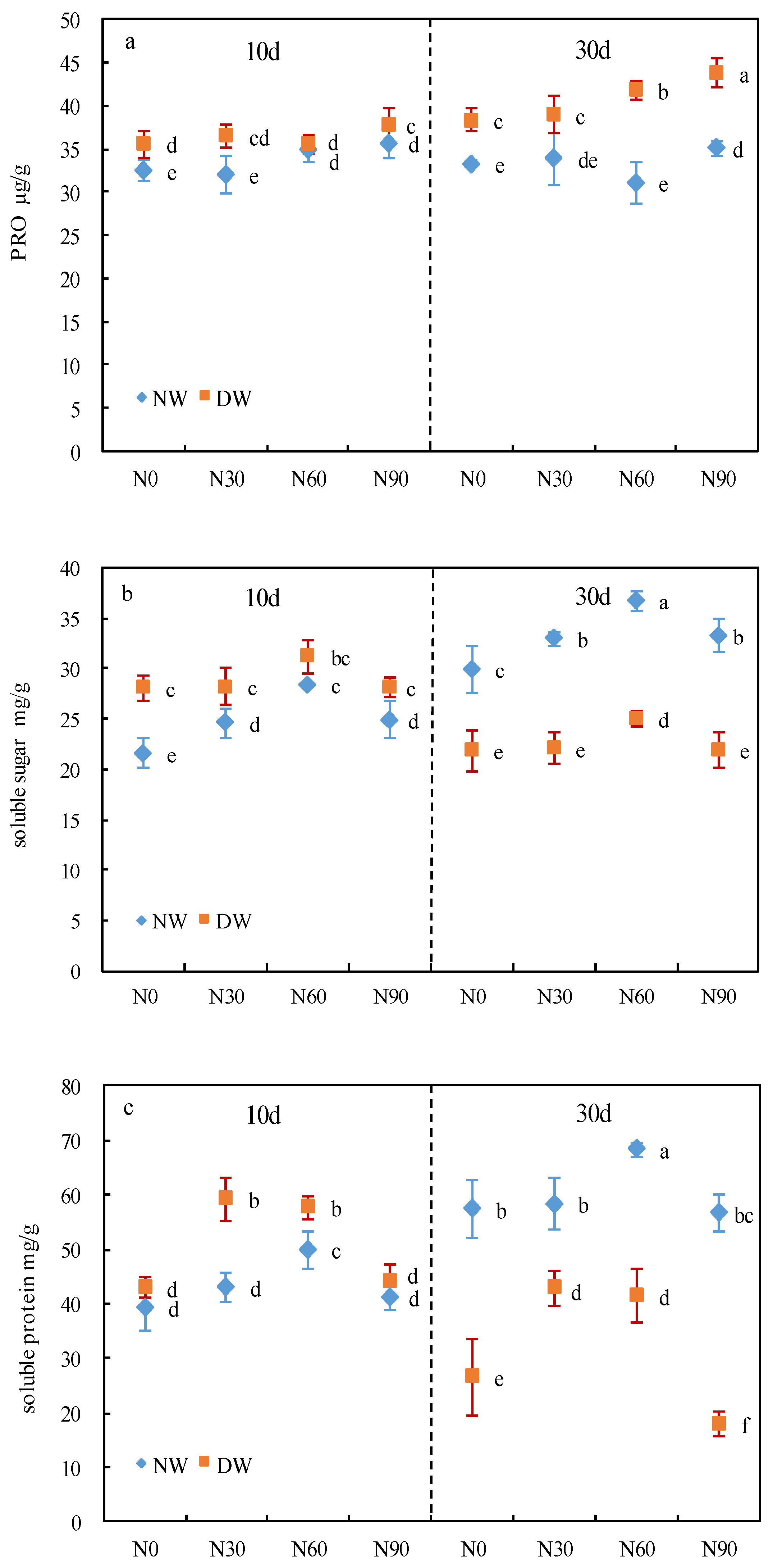
2.5. Determination of Osmotic Adjustment Substance Content
2.6. Determination of Chlorophyll Content
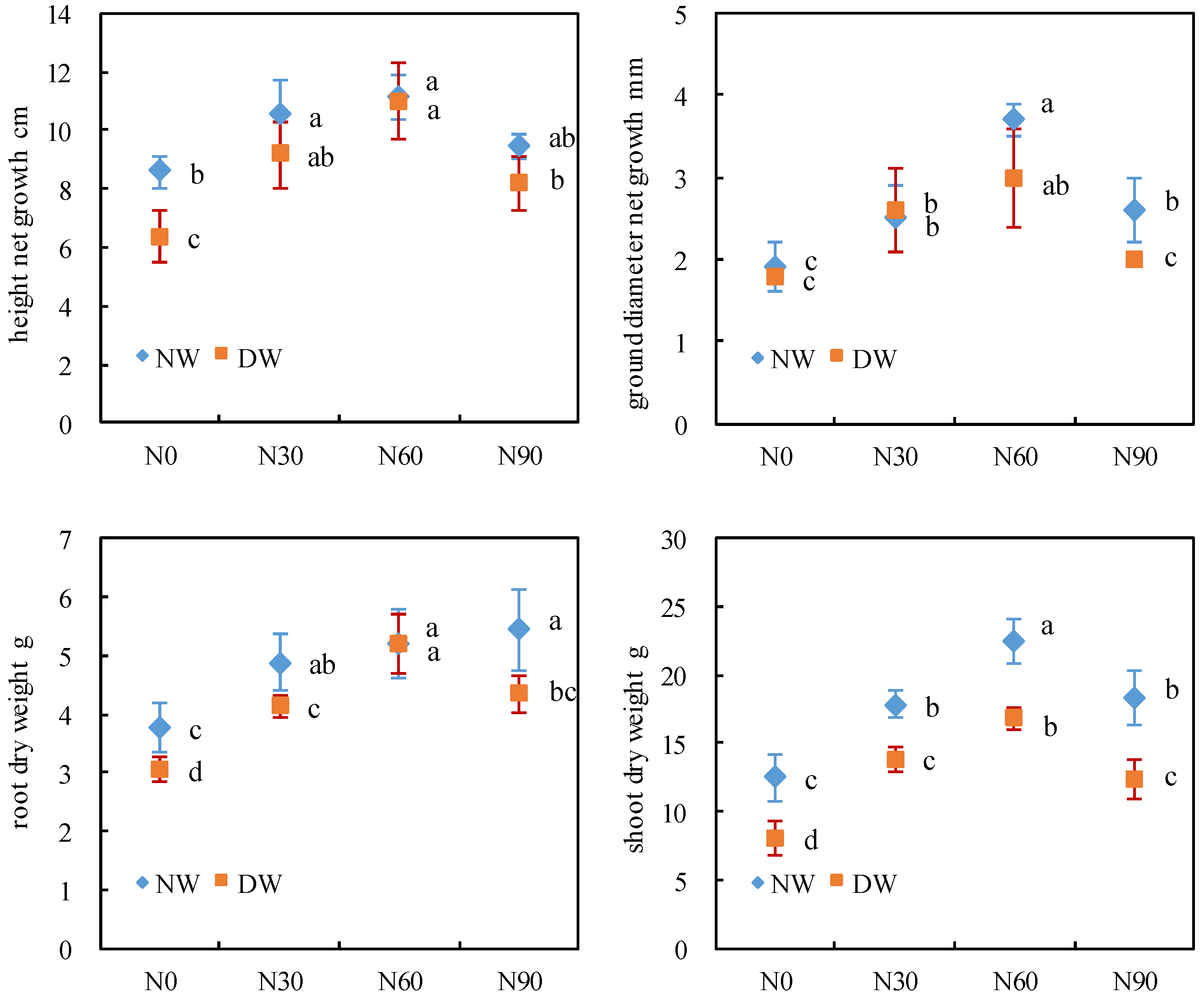
2.7. Determination of Growth Parameters
2.8. Statistical Analysis
3. Results
3.1. Cell Membrane Permeability
3.2. Protective Enzyme System
3.3. Osmoregulatory Substances
3.4. Chlorophyll Content
3.5. Growth Indicators
3.6. ANOVA Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Payne, R.J.; Dise, N.B.; Field, C.D.; Dore, A.J.; Caporn, S.J.; Stevens, C.J. Nitrogen deposition and plant biodiversity: Past, present, and future. Front. Ecol. Environ. 2017, 15, 431–436. [Google Scholar] [CrossRef]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Reay, D.S.; Dentener, F.; Smith, P.; Grace, J.; Feely, R.A. Global nitrogen deposition and carbon sinks. Nat. Geosci. 2008, 1, 430–437. [Google Scholar] [CrossRef]
- Wilkins, K.; Aherne, J. Vegetation community change in Atlantic oak woodlands along a nitrogen deposition gradient. Environ. Pollut. 2016, 216, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Stevens, C.J.; David, T.I.; Storkey, J. Atmospheric nitrogen deposition in terrestrial ecosystems: Its impact on plant communities and consequences across trophic levels. Funct. Ecol. 2018, 32, 1757–1769. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Jia, Y.; He, N.; Zhu, J.; Chen, Z.; Wang, Q.; Piao, S.; Liu, X.; He, H.; Guo, X.; et al. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nat. Geosci. 2019, 12, 424–429. [Google Scholar] [CrossRef]
- Stevens, C.J. Impact of Nitrogen Deposition on the Species Richness of Grasslands. Science 2004, 303, 1876–1879. [Google Scholar] [CrossRef] [Green Version]
- Lilleskov, E.A.; Kuyper, T.W.; Bidartondo, M.I.; Hobbie, E.A. Atmospheric nitrogen deposition impacts on the structure and function of forest mycorrhizal communities: A review. Environ. Pollut. 2019, 246, 148–162. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Chang, S.X.; Cai, Z.; Müller, C.; Zhang, J. Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystems: A review. Environ. Pollut. 2019, 244, 608–616. [Google Scholar] [CrossRef]
- Etzold, S.; Ferretti, M.; Reinds, G.J.; Solberg, S.; Gessler, A.; Waldner, P.; Schaub, M.; Simpson, D.; Benham, S.; Hansen, K.; et al. Nitrogen deposition is the most important environmental driver of growth of pure, even-aged and managed European forests. Forest Ecol. Manag. 2020, 458, 117762. [Google Scholar] [CrossRef]
- Schulte-Uebbing, L.F.; Ros, G.H.; de Vries, W. Experimental evidence shows minor contribution of nitrogen deposition to global forest carbon sequestration. Glob. Change Biol. 2021, 28, 899–917. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.J.; Li, M.Y.; Jiang, S.Y.; Li, W.; Guo, S.X.; Li, H.M.; Guo, X. Comparison between the exotic Coreopsis grandiflora and native Dendranthema indicum across variable nitrogen deposition conditions. Acta Physiol. Plant 2022, 44, 82. [Google Scholar] [CrossRef]
- Schulte-Uebbing, L.; de Vries, W. Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate, and boreal forests: A meta-analysis. Glob. Change Biol. 2017, 24, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Simkin, S.M.; Allen, E.B.; Bowman, W.D.; Clark, C.M.; Belnap, J.; Brooks, M.L.; Cade, B.S.; Collins, S.L.; Geiser, L.H.; Gilliam, F.S.; et al. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 4086–4091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arróniz-Crespo, M.; Leake, J.R.; Horton, P.; Phoenix, G.K. Bryophyte physiological responses to, and recovery from, long-term nitrogen deposition and phosphorus fertilisation in acidic grassland. New Phytol. 2008, 180, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Granath, G.; Wiedermann, M.M.; Strengbom, J. Physiological responses to nitrogen and sulphur addition and raised temperature in Sphagnum balticum. Oecologia 2009, 161, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Blanco, J.A.; Jiang, H.; Kimmins, J.P.H. Effects of nitrogen deposition on carbon sequestration in Chinese fir forest ecosystems. Sci. Total Environ. 2012, 416, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Xu, W.; Li, Q.; Han, M.; Tang, A.; Zhang, Y.; Luo, X.; Shen, J.; Wang, W.; Li, K.; et al. Changes of nitrogen deposition in China from 1980 to 2018. Environ. Int. 2020, 144, 106022. [Google Scholar] [CrossRef]
- Hoerling, M. The Perfect Ocean for Drought. Science 2003, 299, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.D. An ecological perspective on extreme climatic events: A synthetic definition and framework to guide future research. J. Ecol. 2011, 99, 656–663. [Google Scholar] [CrossRef]
- Knapp, A.K.; Hoover, D.L.; Wilcox, K.R.; Avolio, M.L.; Koerner, S.E.; La Pierre, K.J.; Loik, M.E.; Luo, Y.; Sala, O.E.; Smith, M.D. Characterizing differences in precipitation regimes of extreme wet and dry years: Implications for climate change experiments. Glob. Change Biol. 2015, 21, 2624–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef]
- Zhu, K.; Chiariello, N.R.; Tobeck, T.; Fukami, T.; Field, C.B. Nonlinear, interacting responses to climate limit grassland production under global change. Proc. Natl. Acad. Sci. USA 2016, 113, 10589–10594. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Grünefeldt, M.; Friedrich, U.; Klotz, M.; Von Oheimb, G.; Härdtle, W. Nitrogen deposition and drought events have non-additive effects on plant growth—Evidence from greenhouse experiments. Plant Biosyst. 2013, 149, 424–432. [Google Scholar] [CrossRef]
- Yu, Y.; Cheng, H.; Wu, B.; Wang, C. Combined effects of drought stress and different forms of nitrogen deposition as response mechanism to environmental change. Int. J. Environ. Sci. 2022, 1–16. [Google Scholar] [CrossRef]
- Kou, L.; Chen, W.; Jiang, L.; Dai, X.; Fu, X.; Wang, H.; Li, S. Simulated nitrogen deposition affects stoichiometry of multiple elements in resource-acquiring plant organs in a seasonally dry subtropical forest. Sci. Total Environ. 2018, 624, 611–620. [Google Scholar] [CrossRef]
- Wang, Y.; Meng, B.; Zhong, S.; Wang, D.; Ma, J.; Sun, W. Aboveground biomass and root/shoot ratio regulated drought susceptibility of ecosystem carbon exchange in a meadow steppe. Plant Soil 2018, 432, 259–272. [Google Scholar] [CrossRef]
- Friedrich, U.; von Oheimb, G.; Kriebitzsch, W.-U.; Schleßelmann, K.; Weber, M.S.; Härdtle, W. Nitrogen deposition increases susceptibility to drought—Experimental evidence with the perennial grass Molinia caerulea (L.) Moench. Plant Soil 2011, 353, 59–71. [Google Scholar] [CrossRef]
- Shi, W.; Lin, L.; Shao, S.; He, A.; Ying, Y. Effects of simulated nitrogen deposition on Phyllostachys edulis (Carr.) seedlings under different watering conditions: Is seedling drought tolerance related to nitrogen metabolism? Plant Soil 2020, 448, 539–552. [Google Scholar] [CrossRef]
- Valliere, J.M.; Irvine, I.C.; Santiago, L.; Allen, E.B. High N, dry: Experimental nitrogen deposition exacerbates native shrub loss and nonnative plant invasion during extreme drought. Glob. Change Biol. 2017, 23, 4333–4345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivovaroff, A.L.; Santiago, L.S.; Vourlitis, G.L.; Grantz, D.A.; Allen, M.F. Plant hydraulic responses to long-term dry season nitrogen deposition alter drought tolerance in a Mediterranean-type ecosystem. Oecologia 2016, 181, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Drewniak, B.; Gonzalez-Meler, M. Earth system model needs for including the interactive representation of nitrogen deposition and drought effects on forested ecosystems. Forests 2017, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Van der Graaf, S.C.; Janssen, T.A.J.; Erisman, J.W.; Schaap, M. Nitrogen deposition shows no consistent negative nor positive effect on the response of forest productivity to drought across European FLUXNET forest sites. Environ. Res. Commun. 2021, 3, 125003. [Google Scholar] [CrossRef]
- Li, P.; Zhang, L.; Yu, G.; Liu, C.; Ren, X.; He, H.; Liu, M.; Wang, H.; Zhu, J.; Ge, R.; et al. Interactive effects of seasonal drought and nitrogen deposition on carbon fluxes in a subtropical evergreen coniferous forest in the East Asian monsoon region. Agric. Forest Meteorol. 2018, 263, 90–99. [Google Scholar] [CrossRef]
- Hu, P.W.; Duan, L.; Wang, M.N.; Wang, Z.F.; Chen, H.F. Phylogeographic study on Bretschneidera sinensis inferred from AFLP data. Plant Sci. J. 2017, 35, 10. [Google Scholar]
- Qiao, Q.; Qin, X.S.; Qi, F.W.; Chen, H.F.; Liu, D.M. Death causes and conservation strategies of the annual regenerated seedlings of rare plant, Bretschneidera sinensis. Acta Ecol. Sin. 2011, 31, 8. [Google Scholar]
- Gao, Y.; Ma, J.; Zheng, J.C.; Chen, J.; Chen, M.; Zhou, Y.B.; Fu, J.D.; Xu, Z.S.; Ma, Y.Z. The elongation factor GmEF4 is involved in the response to drought and salt tolerance in soybean. Int. J. Mol. Sci. 2019, 20, 3001. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Sun, Y.; Li, L.; Bi, Y.; Hussain, R.; Zhang, R.; Long, H.; Nan, M.; Pu, L. Acetylsalicylic acid (ASA) induced fusarium rot resistance and suppressed neosolaniol production by elevation of ROS metabolism in muskmelon fruit. Sci. Hrotic. 2020, 265, 109264. [Google Scholar] [CrossRef]
- Wang, X.; Wei, X.; Wu, G.; Chen, S. High nitrate or ammonium applications alleviated photosynthetic decline of Phoebe bournei seedlings under elevated carbon dioxide. Forests 2020, 11, 293. [Google Scholar] [CrossRef] [Green Version]
- Yaméogo, C.W.; Bengaly, M.D.; Savadogo, A.; Nikiema, P.A.; Traore, S.A. Determination of chemical composition and nutritional values of Moringa oleifera leaves. Pak. J. Nutr. 2011, 10, 264–268. [Google Scholar] [CrossRef]
- Hsiao, T.C. Plant responses to water stress. Annu. Rev. Plant Physiol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Jaffel, K.; Sai, S.; Bouraoui, N.K.; Ammar, R.B.; Legendre, L.; Lachâal, M.; Marzouk, B. Influence of salt stress on growth, lipid peroxidation and antioxidative enzyme activity in borage (Borago officinalis L.). Plant Biosyst. 2011, 145, 362–369. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Setia, N.; Kaur, S.; Kohli, R.K. 2-Benzoxazolinone (BOA) induced oxidative stress, lipid peroxidation and changes in some antioxidant enzyme activities in mung bean (Phaseolus aureus). Plant Physiol. Bioch. 2006, 44, 819–827. [Google Scholar] [CrossRef]
- Xu, J.; Jin, J.; Zhao, H.; Li, K. Drought stress tolerance analysis of Populus ussuriensis clones with different ploidies. J. For. Res. 2018, 30, 1267–1275. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Pessarakli, M. Effect of silicon on photosynthetic gas exchange, photosynthetic pigments, cell membrane stability and relative water content of different wheat cultivars under drought stress conditions. J. Plant Nutr. 2016, 39, 1001–1015. [Google Scholar] [CrossRef]
- Tao, C.Y.; Shao, S.L.; Shi, W.H.; Lin, L.; Tang, Y.L.; Ying, Y.Q. Effects of nitrogen deposition on biomass and protective enzyme activities of Phyllostachysedulis seedlings under drought stress. Sci. Silvae Sin. 2019, 55, 31–40. [Google Scholar]
- Alici, E.; Arabaci, G. Determination of SOD, POD, PPO and CAT Enzyme Activities in Rumex obtusifolius L. Annu. Res. Rev. Biol. 2016, 11, 1–7. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Ashraf, M. Bioregulators: Unlocking their potential role in regulation of the plant oxidative defense system. Plant Mol. Biol. 2020, 105, 11–41. [Google Scholar] [CrossRef]
- Thatoi, H.N.; Patra, J.K.; Das, S.K. Free radical scavenging and antioxidant potential of mangrove plants: A review. Acta Physiol. Plant. 2013, 36, 561–579. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.; Osuna, C.; Gitto, E. Actions of melatonin in the reduction of oxidative stress. J. Biomed. Sci. 2000, 7, 444–458. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Guo, W.; Liu, J.; Du, N.; Wang, R. Increased nitrogen deposition alleviated the adverse effects of drought stress on Quercus variabilis and Quercus mongolica seedlings. Acta Physiol. Plant. 2015, 37, 107. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plantarum. 2021, 172, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wu, J.; Li, Q.; Hänninen, H.; Peng, C.; Yao, H.; Song, X.; Ying, Y. Nitrogen deposition enhances photosynthesis in Moso bamboo but increases susceptibility to other stress factors. Front. Plant Sci. 2017, 8, 1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wei, X. Dark septate endophyte improves the drought-stress resistance of Ormosia hosiei seedlings by altering leaf morphology and photosynthetic characteristics. Plant Ecol. 2021, 222, 761–771. [Google Scholar] [CrossRef]
- Zhou, L.X.; Cao, H.X.; Liu, Y.J. Effects of seawater stress on physiological and biochemical characteristics of coconut seedling. J. South. Agric. 2018, 49, 2013–2019. [Google Scholar]
- Duan, H.L.; Wu, J.P.; Liu, W.F.; Liao, Y.C.; Zhang, H.N.; Fan, H.B. Water relations and carbon dynamics under drought stress and the mechanisms of drought-induced tree mortality. Sci. Silvae Sin. 2015, 51, 113–120. [Google Scholar]
- Shi, D.; Zhuang, K.; Chen, Y.; Xu, F.; Hu, Z.; Shen, Z. Effects of excess ammoniacal nitrogen (NH4+-N) on pigments, photosynthetic rates, chloroplast ultrastructure, proteomics, formation of reactive oxygen species and enzymatic activity in submerged plant Hydrilla verticillata (Lf) Royle. Aquat. Toxicol. 2020, 226, 105585. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Chl a (mg·g−1) | Chl b (mg·g−1) | Chl a + b (mg·g−1) | Chl a/b | ||
|---|---|---|---|---|---|---|
| 10 d | NW | N0 | 1.46 ± 0.04 c | 0.43 ± 0.03 bc | 1.89 ± 0.07 b | 3.42 ± 0.05 ab |
| N30 | 1.51 ± 0.05 bc | 0.47 ± 0.01 b | 1.98 ± 0.05 b | 3.25 ± 0.03 bc | ||
| N60 | 1.61 ± 0.11 b | 0.49 ± 0.04 b | 2.10 ± 0.17 ab | 3.32 ± 0.11 bc | ||
| N90 | 1.14 ± 0.07 de | 0.34 ± 0.02 c | 1.48 ± 0.02 d | 3.45 ± 0.09 a | ||
| DW | N0 | 1.02 ± 0.06 e | 0.30 ± 0.02 cd | 1.32 ± 0.02 e | 3.44 ± 0.06 a | |
| N30 | 0.97 ± 0.04 e | 0.29 ± 0.01 d | 1.26 ± 0.05 e | 3.40 ± 0.12 ab | ||
| N60 | 1.17 ± 0.05 d | 0.34 ± 0.01 c | 1.52 ± 0.05 cd | 3.46 ± 0.07 a | ||
| N90 | 1.01 ± 0.10 e | 0.29 ± 0.02 d | 1.30 ± 0.03 e | 3.46 ± 0.03 a | ||
| 30 d | NW | N0 | 1.45 ± 0.06 c | 0.44 ± 0.05 bc | 1.89 ± 0.08 b | 3.32 ± 0.07 bc |
| N30 | 1.56 ± 0.05 b | 0.48 ± 0.01 b | 2.04 ± 0.06 b | 3.29 ± 0.03 bc | ||
| N60 | 1.89 ± 0.18 a | 0.60 ± 0.05 a | 2.49 ± 0.18 a | 3.19 ± 0.15 c | ||
| N90 | 1.25 ± 0.02 d | 0.36 ± 0.01 c | 1.61 ± 0.03 c | 3.49 ± 0.08 a | ||
| DW | N0 | 1.03 ± 0.02 e | 0.30 ± 0.01 cd | 1.33 ± 0.03 e | 3.42 ± 0.09 ab | |
| N30 | 0.84 ± 0.07 f | 0.24 ± 0.01 d | 1.07 ± 0.07 f | 3.54 ± 0.19 a | ||
| N60 | 0.98 ± 0.03 e | 0.28 ± 0.01 d | 1.26 ± 0.04 e | 3.46 ± 0.05 a | ||
| N90 | 0.70 ± 0.08 g | 0.20 ± 0.02 d | 0.90 ± 0.10 f | 3.56 ± 0.04 a | ||
| Effects | MDA | RC | SOD | POD | CAT | PRO | Soluble Protein | Soluble Sugar | Chl a | Chl b | Chl a + b | Chl a/b |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Water supply level (W) | *** | *** | ns | ns | * | ** | ns | ns | *** | *** | *** | *** |
| Nitrogen deposition level (N) | * | *** | ns | ns | ns | ns | * | ns | *** | *** | *** | ns |
| W × N | ns | ns | ns | ns | ns | ns | * | ns | *** | ** | *** | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wu, G.; Li, D.; Song, X. Moderate Nitrogen Deposition Alleviates Drought Stress of Bretschneidera sinensis. Forests 2023, 14, 137. https://doi.org/10.3390/f14010137
Wang X, Wu G, Li D, Song X. Moderate Nitrogen Deposition Alleviates Drought Stress of Bretschneidera sinensis. Forests. 2023; 14(1):137. https://doi.org/10.3390/f14010137
Chicago/Turabian StyleWang, Xiao, Gaoyin Wu, Deyan Li, and Xiaohui Song. 2023. "Moderate Nitrogen Deposition Alleviates Drought Stress of Bretschneidera sinensis" Forests 14, no. 1: 137. https://doi.org/10.3390/f14010137