

Mechanisms Underlying Aboveground and Belowground Litter Decomposition Converge over Time under Nutrient Deposition
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
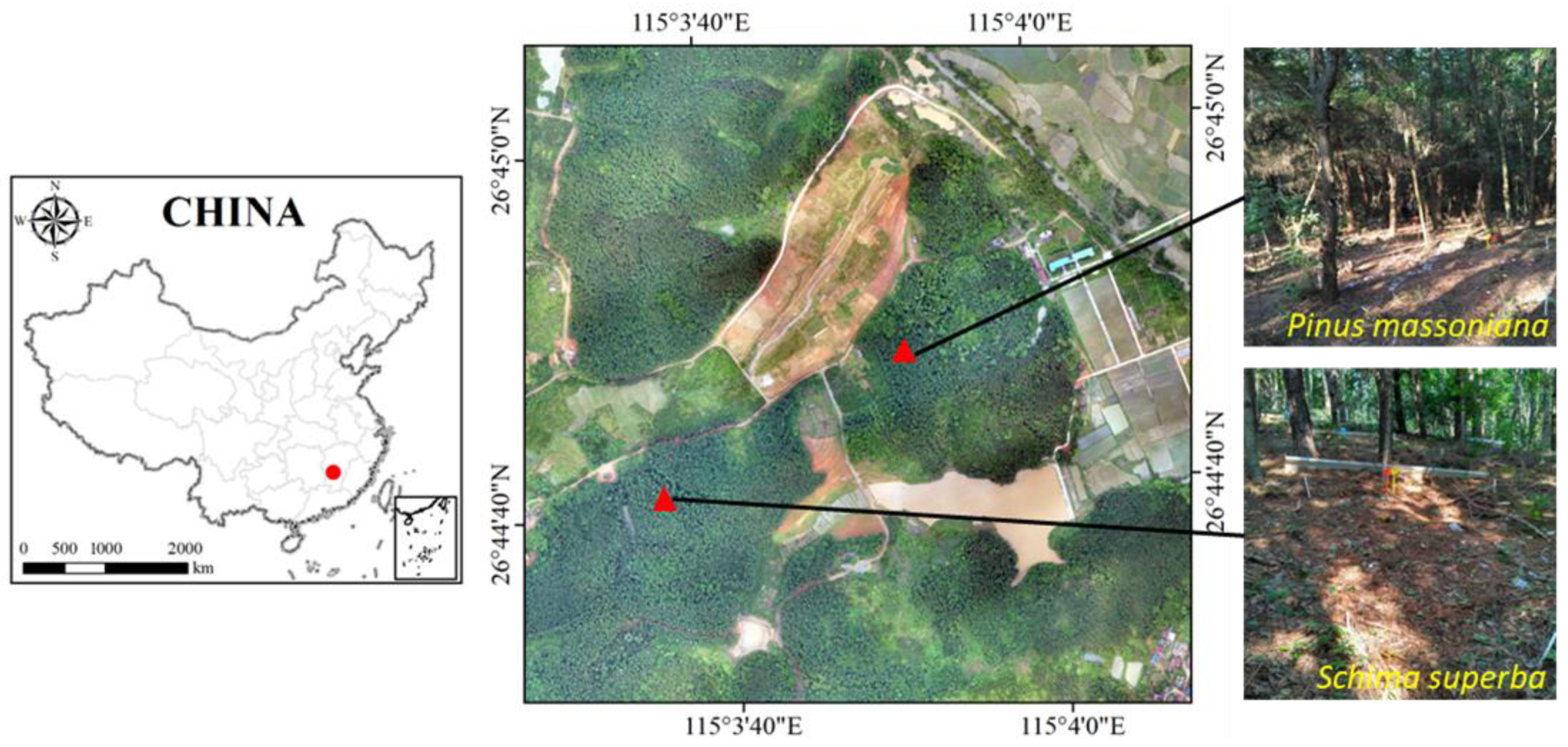
2.1. Site Description
2.2. Experimental Treatment
2.3. Litter Material Preparation
2.4. Litterbag Deployment and Retrieval
2.5. Chemistry Analysis
2.6. Microbial Extracellular Enzymatic Activity Assay
2.7. Statistical Analysis
3. Results
3.1. Initial Chemical Parameters
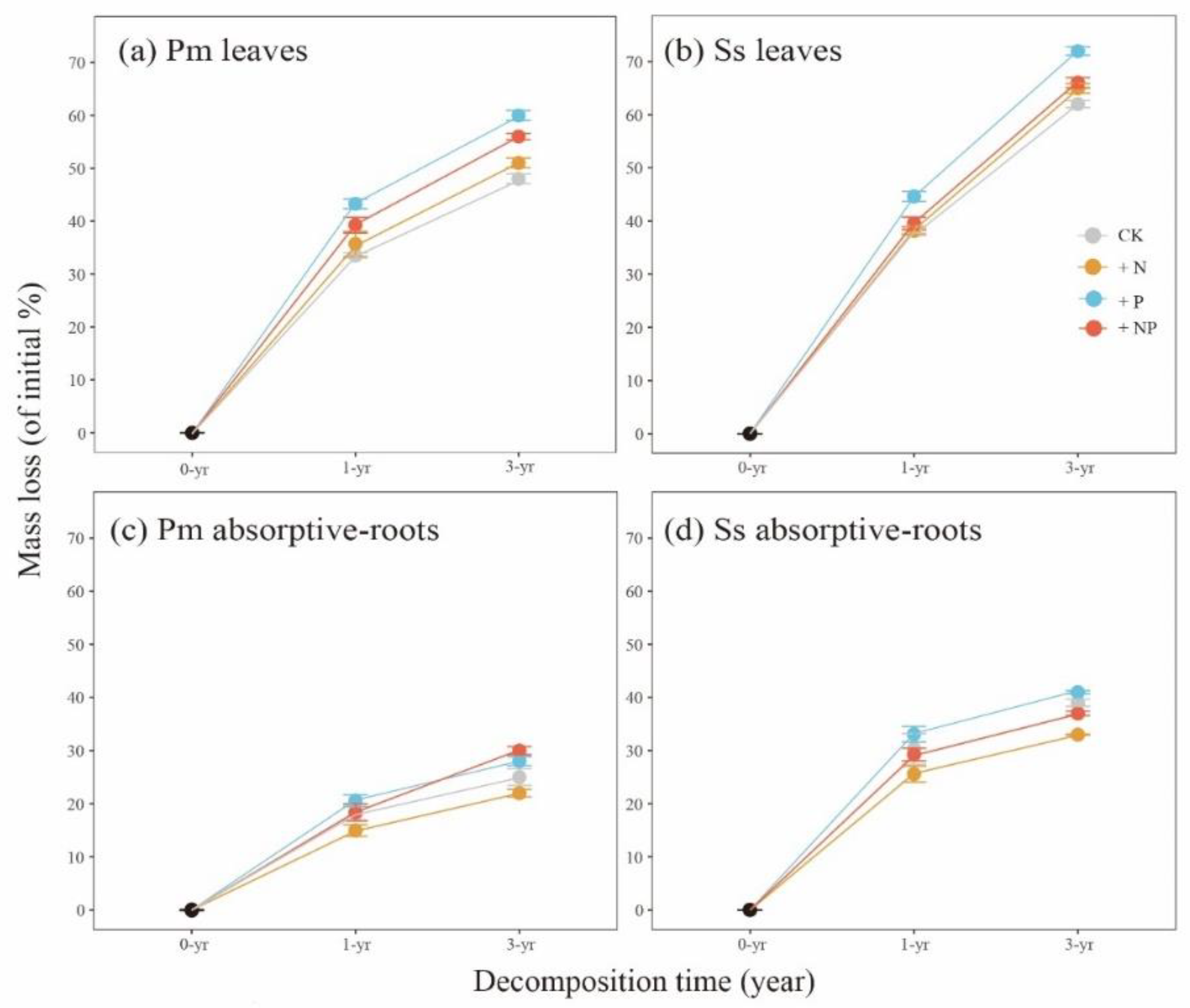
3.2. Effects of Different Treatments on the Mass Loss of Four Substrates
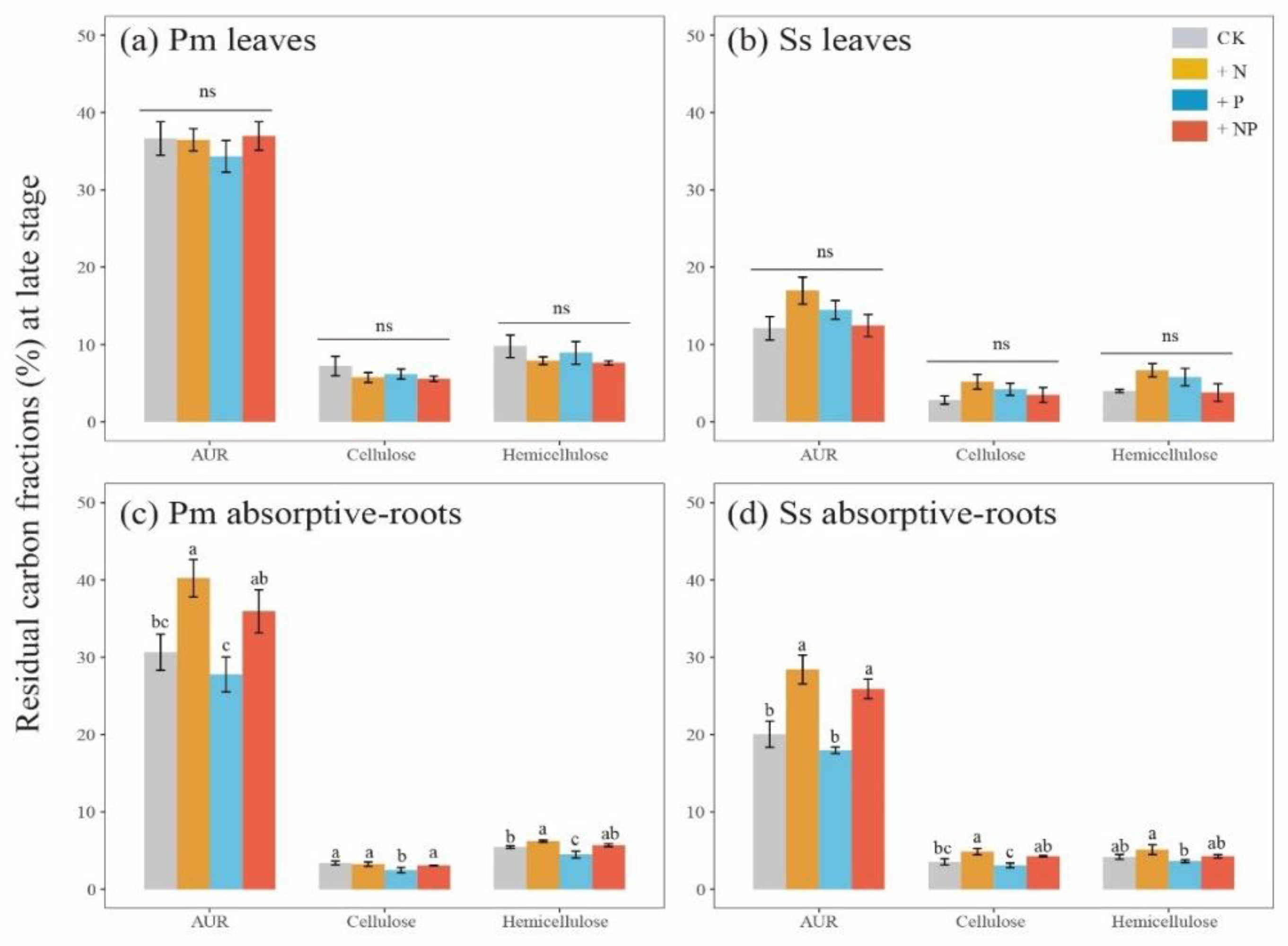
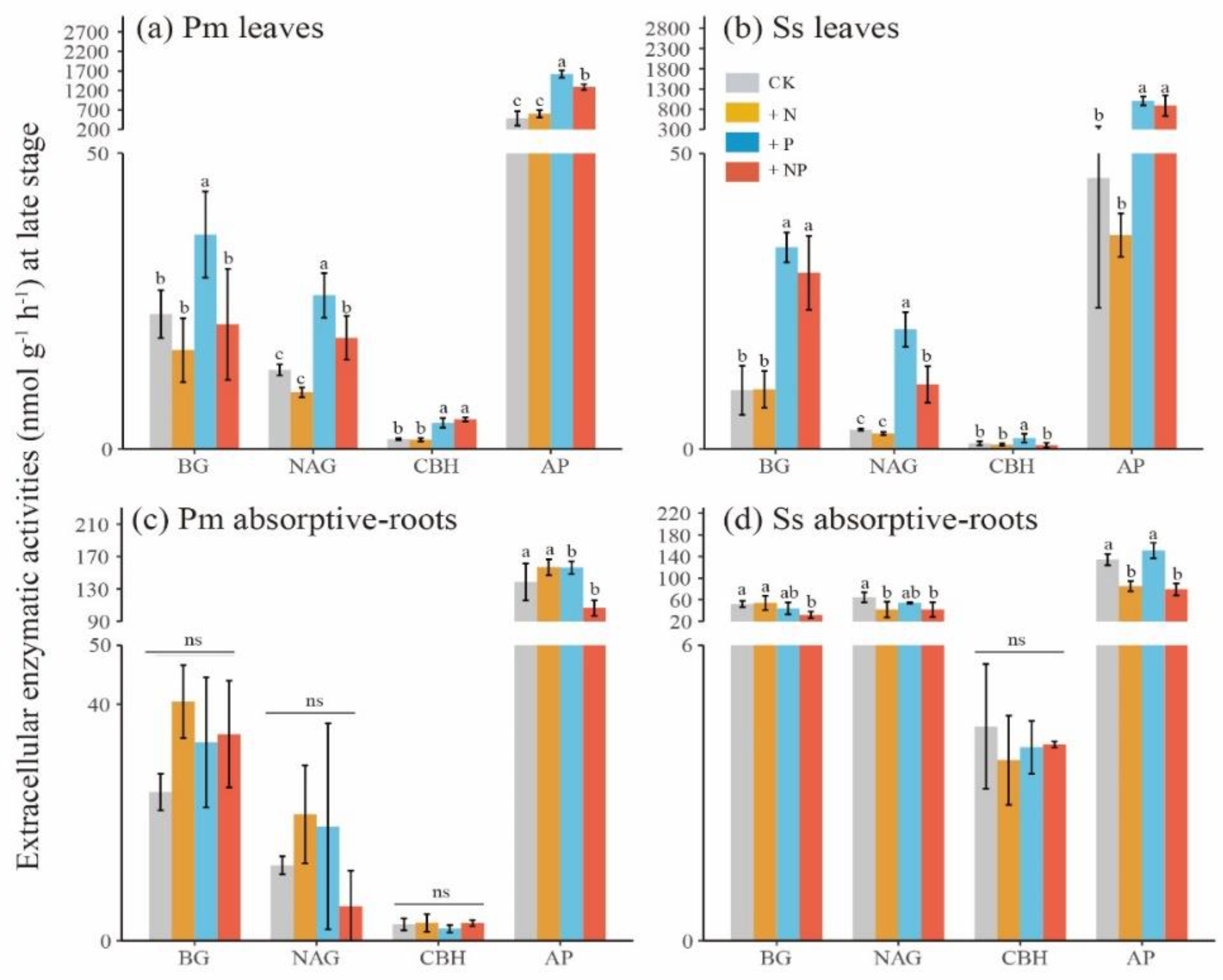
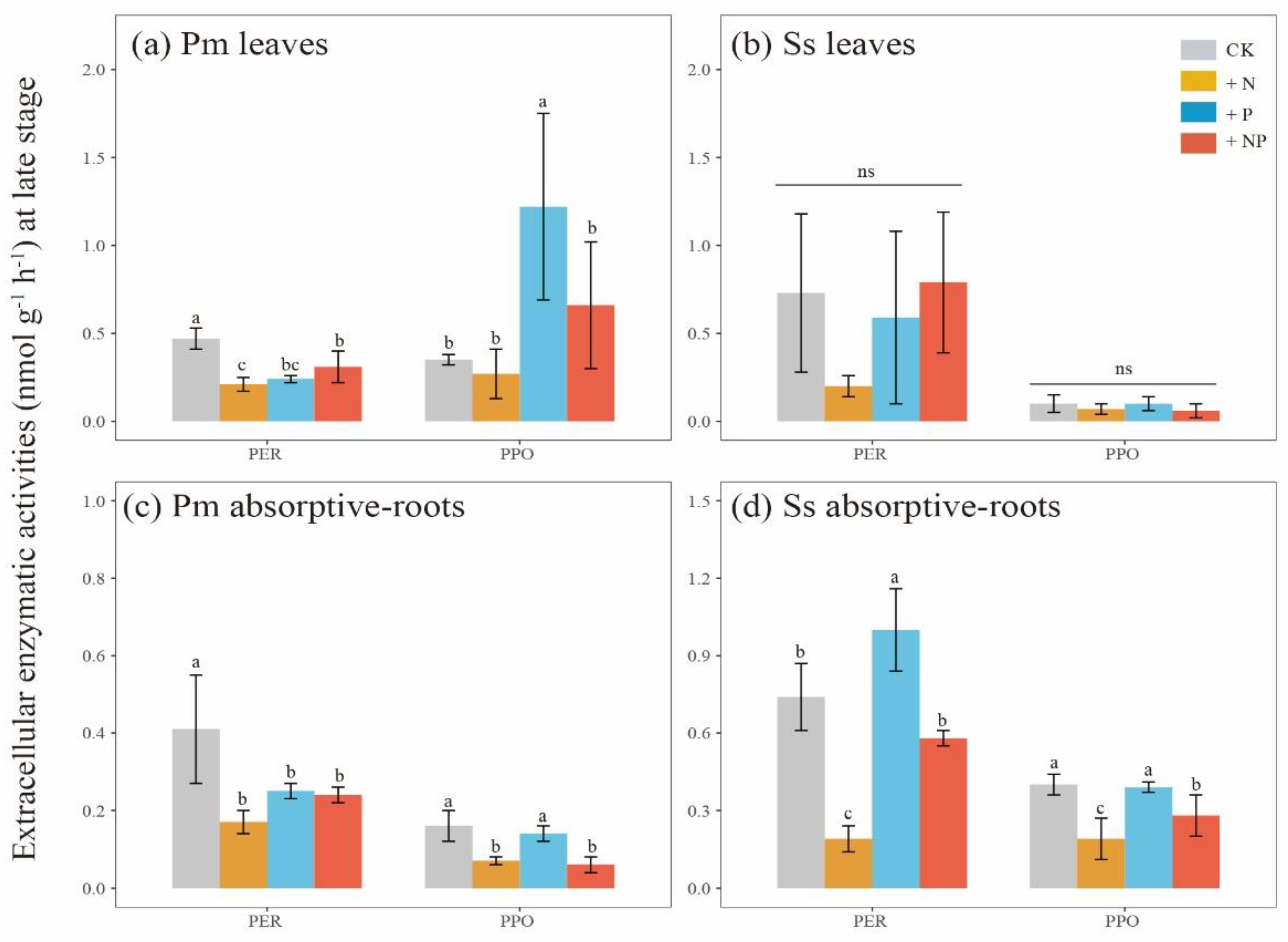
3.3. Carbon Fractions and Microbial Enzymatic Activity
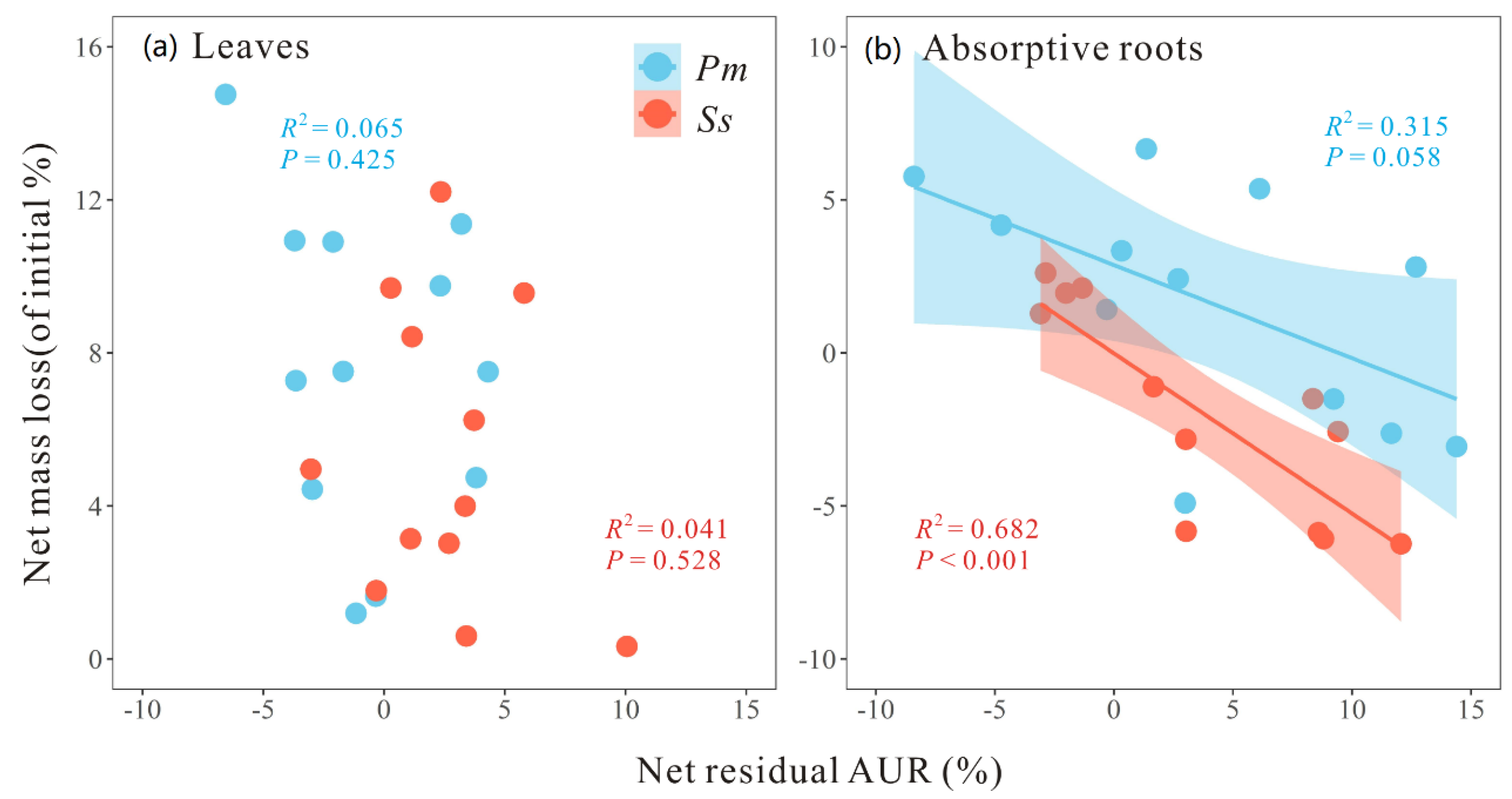
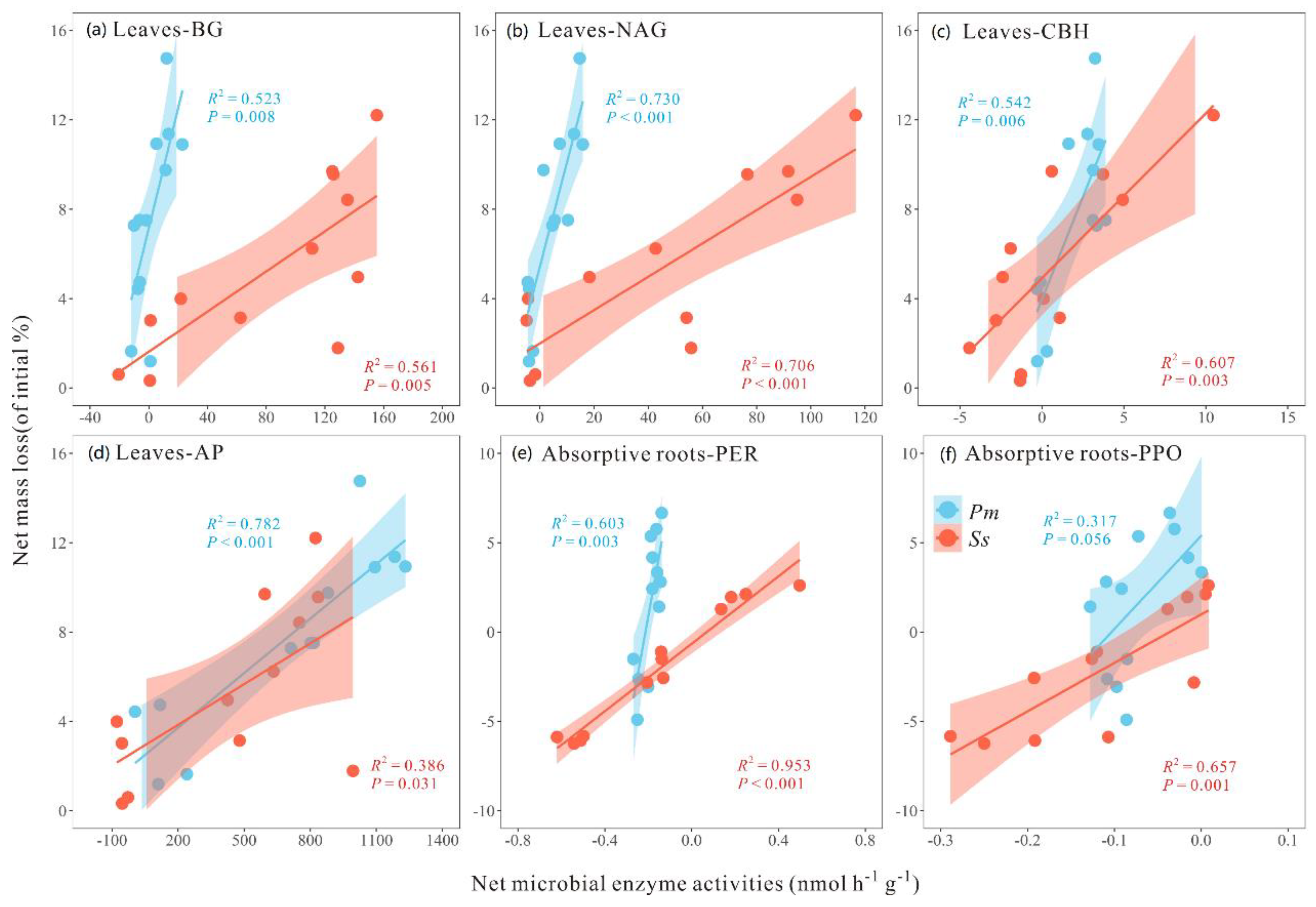
3.4. Relationships among Mass Loss, AUR Concentration, and Enzymatic Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knorr, M.; Frey, S.D.; Curtis, P.S. Nitrogen additions and litter decomposition: A meta-analysis. Ecology 2005, 86, 3252–3257. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.A.; Luo, Y.Q.; Chen, H.Y.H.; Ruan, H.H. Responses of litter decomposition and nutrient release to N addition: A meta-analysis of terrestrial ecosystems. Appl. Soil Ecol. 2018, 128, 35–42. [Google Scholar] [CrossRef]
- Berg, B. Initial rates and limit values for decomposition of Scots pine and Norway spruce needle litter: A synthesis for N-fertilized forest stands. Can. J. For. Res. 2000, 30, 122–135. [Google Scholar] [CrossRef]
- Lee, H.; Rahn, T.; Throop, H. An accounting of C-based trace gas release during abiotic plant litter degradation. Glob. Chang. Biol. 2012, 18, 1185–1195. [Google Scholar] [CrossRef]
- Kou, L.; Jiang, L.; Fu, X.L.; Dai, X.Q.; Wang, H.M.; Li, S.G. Nitrogen deposition increases root production and turnover but slows root decomposition in Pinus elliottii plantations. New Phytol. 2018, 218, 1450–1461. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, H.M.; Li, S.G.; Fu, X.L.; Dai, X.Q.; Yan, H.; Kou, L. Mycorrhizal and environmental controls over root trait-decomposition linkage of woody trees. New Phytol. 2021, 229, 284–295. [Google Scholar] [CrossRef]
- Sun, T.; Hobbie, S.E.; Berg, B.; Zhang, H.F.; Wang, Q.K.; Wang, Z.W.; Hättenschwiler, S. Contrasting dynamics and trait controls in first-order root compared with leaf litter decomposition. Proc. Natl. Acad. Sci. USA 2018, 115, 10392–10397. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Kou, L.; Li, S.G. Alterations of early-stage decomposition of leaves and absorptive roots by deposition of nitrogen and phosphorus have contrasting mechanisms. Soil Biol. Biochem. 2018, 127, 213–222. [Google Scholar] [CrossRef]
- Jiang, L.; Kou, L.; Li, S.G. Decomposition of leaf mixtures and absorptive-root mixtures synchronously changes with deposition of nitrogen and phosphorus. Soil Biol. Biochem. 2019, 138, 107602. [Google Scholar] [CrossRef]
- Fahey, T.J.; Siccama, T.G.; Driscoll, C.T.; Likens, G.E.; Campbell, J.; Johnson, C.E.; Battles, J.J.; Aber, J.D.; Cole, J.J.; Fisk, M.C. The biogeochemistry of carbon at Hubbard Brook. Biogeochemistry 2005, 75, 109–176. [Google Scholar] [CrossRef]
- Fu, X.F.; Xu, C.H.; Geng, Q.H.; Ma, X.C.; Zhang, H.G.; Cai, B.; Hu, G.Q.; Xu, X. Effects of nitrogen application on the decomposition of fine roots in temperate forests: A meta-analysis. Plant Soil 2022, 472, 77–89. [Google Scholar] [CrossRef]
- Kou, L.; Chen, W.W.; Zhang, X.Y.; Gao, W.L.; Yang, H.; Li, D.D.; Li, S.G. Differential responses of needle and branch order-based root decay to nitrogen addition: Dominant effects of acid-unhydrolyzable residue and microbial enzymes. Plant Soil 2015, 394, 315–327. [Google Scholar] [CrossRef]
- Norris, M.D.; Avis, P.G.; Reich, P.B.; Hobbie, S.E. Positive feedbacks between decomposition and soil nitrogen availability along fertility gradients. Plant Soil 2013, 367, 347–361. [Google Scholar] [CrossRef]
- Xia, M.X.; Talhelm, A.F.; Pregitzer, K.S. Chronic nitrogen deposition influences the chemical dynamics of leaf litter and fine roots during decomposition. Soil Biol. Biochem. 2017, 112, 24–34. [Google Scholar] [CrossRef]
- Chen, H.; Dong, S.F.; Liu, L.; Ma, C.; Zhang, T.; Zhu, X.M.; Mo, J.M. Effects of experimental nitrogen and phosphorus addition on litter decomposition in an old-growth tropical forest. PLoS ONE 2013, 8, e84101. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Wang, H.M.; Dai, X.Q.; Kou, L.; Meng, S.W.; Fu, X.L. Decoupled responses of leaf and root decomposition to nutrient deposition in a subtropical plantation. Soil Biol. Biochem. 2022, 168, 108643. [Google Scholar] [CrossRef]
- Harantová, L.; Baldrian, P.; Angst, G.; Cajthaml, T.; Straková, P.; Blahut, J.; Veselá, H.; Frouz, J. Tree species identity alters decomposition of understory litter and associated microbial communities: A case study. Biol. Fertil. Soil. 2019, 55, 525–538. [Google Scholar]
- Moorhead, D.L.; Sinsabaugh, R.L. A theoretical model of litter decay and microbial interaction. Ecol. Monogr. 2006, 76, 151–174. [Google Scholar] [CrossRef]
- Bray, S.R.; Kitajima, K.; Mack, M.C. Temporal dynamics of microbial communities on decomposing leaf litter of 10 plant species in relation to decomposition rate. Soil Biol. Biochem. 2012, 49, 30–37. [Google Scholar] [CrossRef]
- Purahong, W.; Krüger, D.; Buscot, F.; Wubet, T. Correlations between the composition of modular fungal communities and litter decomposition-associated ecosystem functions. Fungal Ecol. 2016, 22, 106–114. [Google Scholar] [CrossRef]
- Atiwesh, G.; Parrish, C.C.; Banoub, J.; Le, T.A.T. Lignin degradation by microorganisms: A review. Biotechnol. Prog. 2022, 38, e3226. [Google Scholar] [CrossRef]
- Gołębiewski, M.; Tarasek, A.; Sikora, M.; Deja-Sikora, E.; Tretyn, A.; Niklińska, M. Rapid microbial community changes during initial stages of pine litter decomposition. Microb. Ecol. 2019, 77, 56–75. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Li, Y.J.; Song, S.Y.; Chen, Y.Q.; Chen, G.T.; Tu, L.H. Nitrogen addition slows litter decomposition accompanied by accelerated manganese release: A five-year experiment in a subtropical evergreen broadleaf forest. Soil Biol. Biochem. 2022, 165, 108511. [Google Scholar] [CrossRef]
- Zhu, J.X.; Wang, Q.F.; He, N.P.; Smith, M.D.; Elser, J.J.; Du, J.Q.; Yuan, G.F.; Yu, G.R.; Yu, Q. Imbalanced atmospheric nitrogen and phosphorus depositions in China: Implications for nutrient limitation. J. Geophys. Res. Biogeosci. 2016, 121, 1605–1616. [Google Scholar] [CrossRef]
- Zhu, J.X.; He, N.P.; Wang, Q.F.; Yuan, G.F.; Wen, D.; Yu, G.R.; Jia, Y.L. The composition, spatial patterns, and influencing factors of atmospheric wet nitrogen deposition in Chinese terrestrial ecosystems. Sci. Total Environ. 2015, 511, 777–785. [Google Scholar] [CrossRef]
- Kou, L.; Jiang, L.; Hättenschwiler, S.; Zhang, M.M.; Niu, S.L.; Fu, X.L.; Dai, X.Q.; Yan, H.; Li, S.G.; Wang, H.M. Diversity-decomposition relationships in forests worldwide. eLife 2020, 9, e55813. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine north American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- Bocock, K.L.; Gilbert, O.J.W. The disappearance of leaf litter under different woodland conditions. Plant Soil 1957, 9, 179–185. [Google Scholar] [CrossRef]
- Kampichler, C.; Bruckner, A. The role of microarthropods in terrestrial decomposition: A meta-analysis of 40 years of litterbag studies. Biol. Rev. 2009, 84, 375–389. [Google Scholar] [CrossRef]
- Lytle, E.D.; Cronan, C.S. Comparative soil CO2 evolution, litter decay, and root dynamics in clearcut and uncut spruce-fir forest. For. Ecol. Manag. 1998, 103, 121–128. [Google Scholar] [CrossRef]
- Jing, H.; Zhang, P.; Li, J.J.; Yao, X.; Liu, G.B.; Wang, G.L. Effect of nitrogen addition on the decomposition and release of compounds from fine roots with different diameters: The importance of initial substrate chemistry. Plant Soil 2019, 438, 281–296. [Google Scholar] [CrossRef]
- Xiong, Y.M.; Fan, P.P.; Fu, S.L.; Zeng, H.; Guo, D.L. Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees. Plant Soil 2013, 363, 19–31. [Google Scholar] [CrossRef]
- Weng, C.H.; Peng, X.W.; Han, Y.J. Depolymerization and conversion of lignin to value-added bioproducts by microbial and enzymatic catalysis. Biotechnol. Biofuels. 2021, 14, 84. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.X.; Talhelm, A.F.; Pregitzer, K.S. Long-term simulated atmospheric nitrogen deposition alters leaf and fine root decomposition. Ecosystems 2018, 21, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zabel, R.A.; Morrell, J.J. Wood Microbiology Decay and Its Prevention, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Sun, T.; Dong, L.L.; Wang, Z.W.; Lu, X.T.; Mao, Z.J. Effects of long-term nitrogen deposition on fine root decomposition and its extracellular enzyme activities in temperate forests. Soil Biol. Biochem. 2016, 93, 50–59. [Google Scholar] [CrossRef]
- Hobbie, S.E. Nitrogen effects on litter decomposition: A five-year experiment in eight temperate grassland and forest sites. Ecology 2008, 89, 2633–2644. [Google Scholar] [CrossRef]
- Zhang, W.; Chao, L.; Yang, Q.; Wang, Q.; Fang, Y.; Wang, S. Litter quality mediated nitrogen effect on plant litter decomposition regardless of soil fauna presence. Ecology 2016, 97, 2834–2843. [Google Scholar] [CrossRef]
- Carreiro, M.M.; Repert, D.A.; Parkhurst, D.F.; Sinsabaugh, R.L. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition. Ecology 2000, 81, 2359–2365. [Google Scholar] [CrossRef]
- Tan, X.; Machmuller, M.B.; Cotrufo, M.F.; Shen, W.J. Shifts in fungal biomass and activities of hydrolase and oxidative enzymes explain different responses of litter decomposition to nitrogen addition. Biol. Fertil. Soils 2020, 56, 423–438. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Kou, L.; Li, S.G.; Wang, H.M.; Fu, X.L.; Dai, X.Q. Unaltered phenology but increased production of ectomycorrhizal roots of Pinus elliottii under 4 years of nitrogen addition. New Phytol. 2019, 221, 2228–2238. [Google Scholar] [CrossRef]
- Kou, L.; Guo, D.L.; Yang, H.; Gao, W.L.; Li, S.G. Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China. Plant Soil 2016, 391, 207–218. [Google Scholar] [CrossRef]
- Waring, B.G. Exploring relationships between enzyme activities and leaf litter decomposition in a wet tropical forest. Soil Biol. Biochem. 2013, 64, 89–95. [Google Scholar] [CrossRef]
- Zheng, J.J.; Li, S.G.; Wang, H.M.; Dai, X.Q.; Meng, S.W.; Jiang, L.; Ma, N.; Yan, H.; Fu, X.L.; Kou, L. Home-filed advantage meets priming effect in root decomposition: Implications for belowground carbon dynamics. Funct. Ecol. 2022, 1–14. [Google Scholar] [CrossRef]
- Martins, N.P.; Fuchslueger, L.; Fleischer, K.; Andersen, K.M.; Assis, R.L.; Baccaro, F.B.; Camargo, P.B.; Cordeiro, A.L.; Grandis, A.; Hartley, I.P.; et al. Fine roots stimulate nutrient release during early stages of leaf litter decomposition in a Central Amazon rainforest. Plant Soil 2021, 469, 287–303. [Google Scholar] [CrossRef]
- Marklein, A.R.; Houlton, B.Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef]
- Olander, L.P.; Vitousek, P.M. Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 2000, 49, 175–190. [Google Scholar] [CrossRef]
- Vivanco, L.; Austin, A.T. Intrinsic effects of species on leaf litter and root decomposition: A comparison of temperate grasses from North and South America. Oecologia 2006, 150, 97–107. [Google Scholar] [CrossRef]
- Arenberg, M.R.; Arai, Y. Effects of native leaf litter amendments on phosphorus mineralization in temperate floodplain soils. Chemosphere 2021, 226, 129210. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Reed, S.C.; Townsend, A.R. Nutrient regulation of organic matter decomposition in a tropical rain forest. Ecology 2006, 87, 492–503. [Google Scholar] [CrossRef]
- Yan, H.; Freschet, G.T.; Wang, H.M.; Hogan, J.A.; Li, S.G.; Valverde-Barrants, O.J.; Fu, X.L.; Wang, R.L.; Dai, X.Q.; Jiang, L.; et al. Mycorrhizal symbiosis pathway and edaphic fertility frame root economics space among tree species. New phytol. 2022, 234, 1639–1653. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L. Microbial community response to nitrogen deposition in northern forest ecosystems. Soil Biol. Biochem. 2004, 36, 1443–1451. [Google Scholar] [CrossRef]
- Fraterrigo, J.M.; Balser, T.C.; Turner, M.G. Microbial community variation and its relationship with nitrogen mineralization in historically altered forests. Ecology 2006, 87, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Forstner, S.J.; Wechselberger, V.; Stecher, S.; Müller, S.; Keiblinger, K.M.; Wanek, W.; Schleppi, P.; Gundersen, P.; Tatzber, M.; Gerzabek, M.H.; et al. Resistant soil microbial communities show signs of increasing phosphorus limitation in two temperate forests after long-term nitrogen addition. Front. For. Glob. Chang. 2019, 2, 1–23. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Type | C (mg g−1) | N (mg g−1) | P (mg g−1) | AUR (mg g−1) | Cellulose (mg g−1) | Hemicellulose (mg g−1) | C:N | C:P | N:P | AUR:N | AUR:P |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pinus massoniana | Leaves | 506.90 a (2.83) | 8.07 c (0.51) | 0.59 a (0.03) | 460.43 b (3.62) | 212.40 a (1.65) | 201.34 a (2.91) | 63.50 a (3.49) | 874.12 a (54.13) | 13.83 c (0.82) | 57.66 a (3.09) | 793.72 b (47.86) |
| Absorptive roots | 426.68 c (7.19) | 11.88 b (0.43) | 0.55 a (0.01) | 589.13 a (9.91) | 90.71 c (2.70) | 112.44 c (5.70) | 36.06 b (1.52) | 782.46 a (24.91) | 21.77 b (0.88) | 49.77 a (1.93) | 1080.25 a (33.06) | |
| Schima superba | Leaves | 436.43 c (3.39) | 15.13 a (0.45) | 0.56 a (0.02) | 381.74 c (10.54) | 183.85 b (5.45) | 174.29 b (4.78) | 28.92 b (0.86) | 788.63 a (31.99) | 27.26 a (0.67) | 25.31 b (1.12) | 690.80 b (39.71) |
| Absorptive roots | 463.44 b (2.63) | 14.59 a (0.28) | 0.63 a (0.01) | 429.14 b (4.46) | 201.12 a (2.81) | 179.33 ab (7.22) | 31.79 b (0.55) | 734.07 a (10.45) | 23.10 b (0.36) | 29.43 b (0.39) | 679.68 b (10.16) |
| Source of Variation | C (mg g−1) | N (mg g−1) | P (mg g−1) | AUR (mg g−1) | Cellulose (mg g−1) | Hemicellulose (mg g−1) | C:N | C:P | N:P | AUR:N | AUR:P |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | 0.003 | <0.001 | 0.199 | <0.001 | <0.001 | 0.003 | <0.001 | 0.074 | <0.001 | <0.001 | <0.001 |
| Tissue | <0.001 | 0.002 | 0.396 | <0.001 | <0.001 | <0.001 | <0.001 | 0.054 | 0.021 | 0.344 | 0.002 |
| Species × Tissue | <0.001 | <0.001 | 0.016 | <0.001 | <0.001 | <0.001 | <0.001 | 0.598 | <0.001 | 0.009 | 0.001 |
| Substrates | +N vs. CK | +P vs. CK | +NP vs. CK | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Effect | F | P | Effect | F | P | Effect | F | P | |
| Pinus massoniana leaves | ns | 5.30 | 0.061 | + | 83.99 | <0.001 | + | 53.98 | 0.000 |
| Pinus massoniana absorptive roots | ns | 5.55 | 0.057 | + | 14.56 | 0.009 | ns | 4.89 | 0.069 |
| Schima superba leaves | ns | 3.24 | 0.122 | + | 95.35 | <0.001 | + | 11.86 | 0.014 |
| Schima superba absorptive roots | − | 88.35 | <0.001 | + | 8.38 | 0.028 | − | 7.01 | 0.038 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, L.; Li, S.; Wang, H.; Dai, X.; Meng, S.; Fu, X.; Zheng, J.; Yan, H.; Ma, N.; Xue, Y.; et al. Mechanisms Underlying Aboveground and Belowground Litter Decomposition Converge over Time under Nutrient Deposition. Forests 2023, 14, 130. https://doi.org/10.3390/f14010130
Jiang L, Li S, Wang H, Dai X, Meng S, Fu X, Zheng J, Yan H, Ma N, Xue Y, et al. Mechanisms Underlying Aboveground and Belowground Litter Decomposition Converge over Time under Nutrient Deposition. Forests. 2023; 14(1):130. https://doi.org/10.3390/f14010130
Chicago/Turabian StyleJiang, Lei, Shenggong Li, Huimin Wang, Xiaoqin Dai, Shengwang Meng, Xiaoli Fu, Jiajia Zheng, Han Yan, Ning Ma, Yafang Xue, and et al. 2023. "Mechanisms Underlying Aboveground and Belowground Litter Decomposition Converge over Time under Nutrient Deposition" Forests 14, no. 1: 130. https://doi.org/10.3390/f14010130