
Temporal Changes in Growth–Climate Relationship of Pinus taiwanensis Hayata in Subtropical China

Abstract
:1. Introduction
2. Materials and Methods
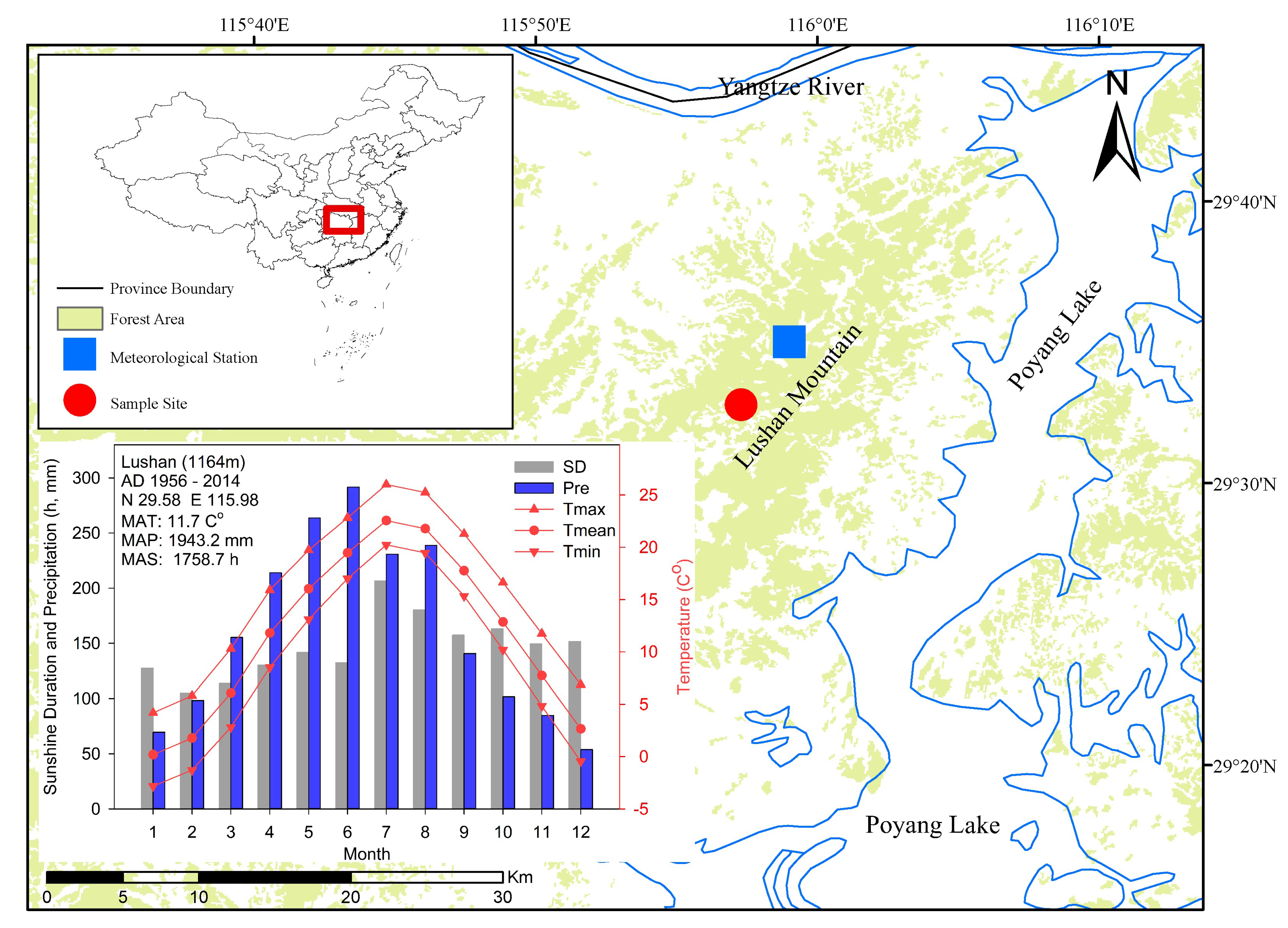
2.1. Study Area
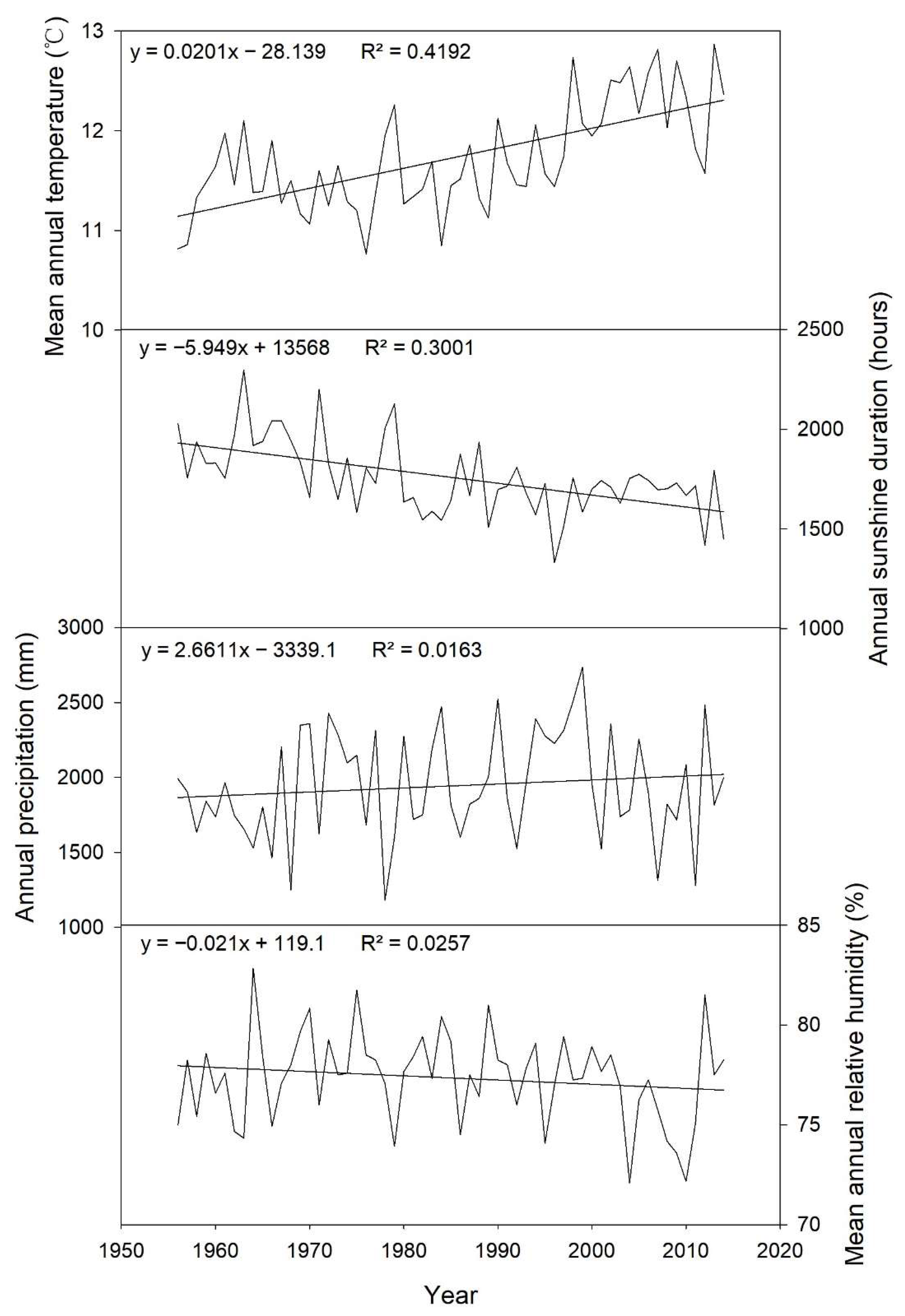
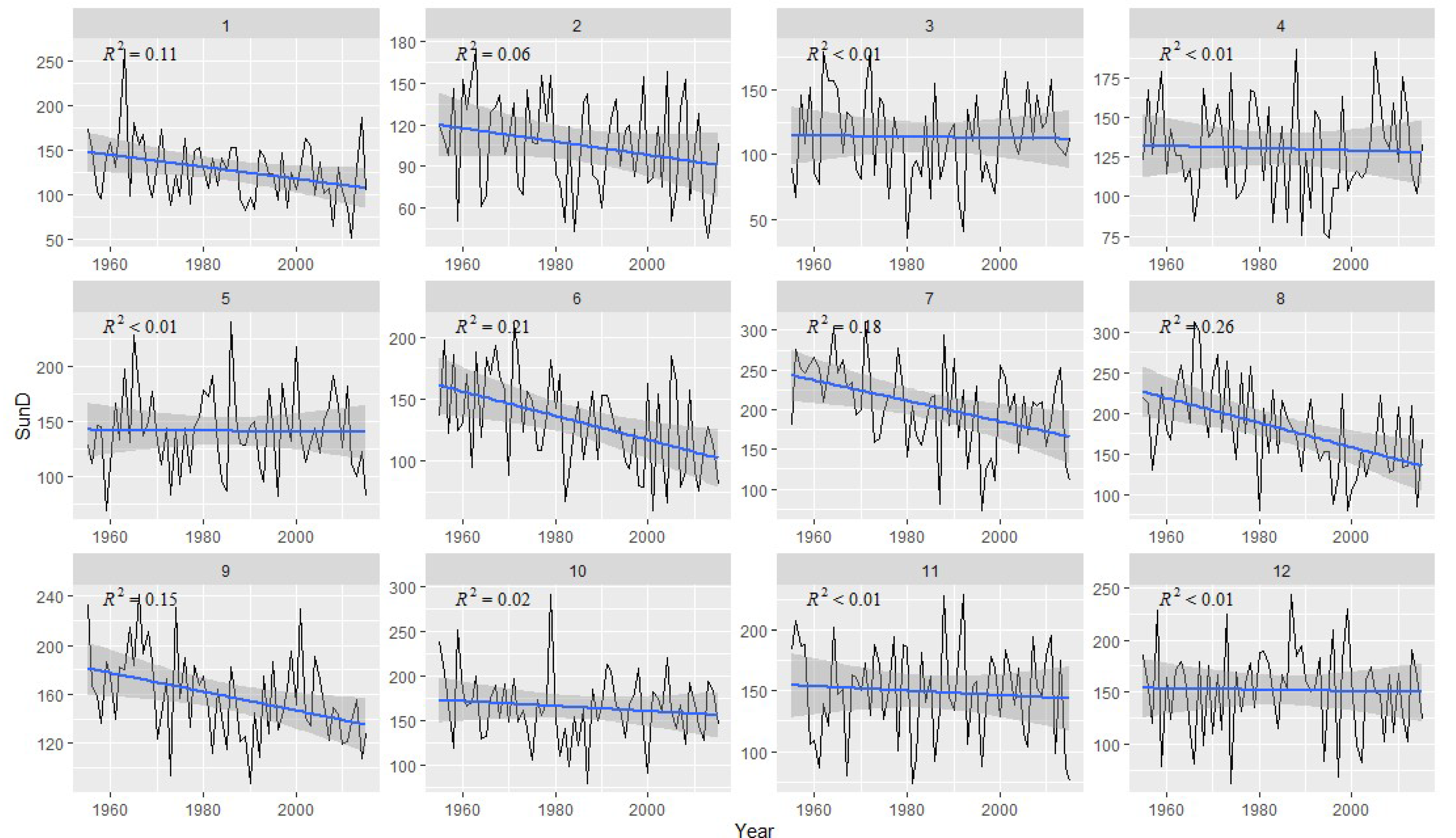
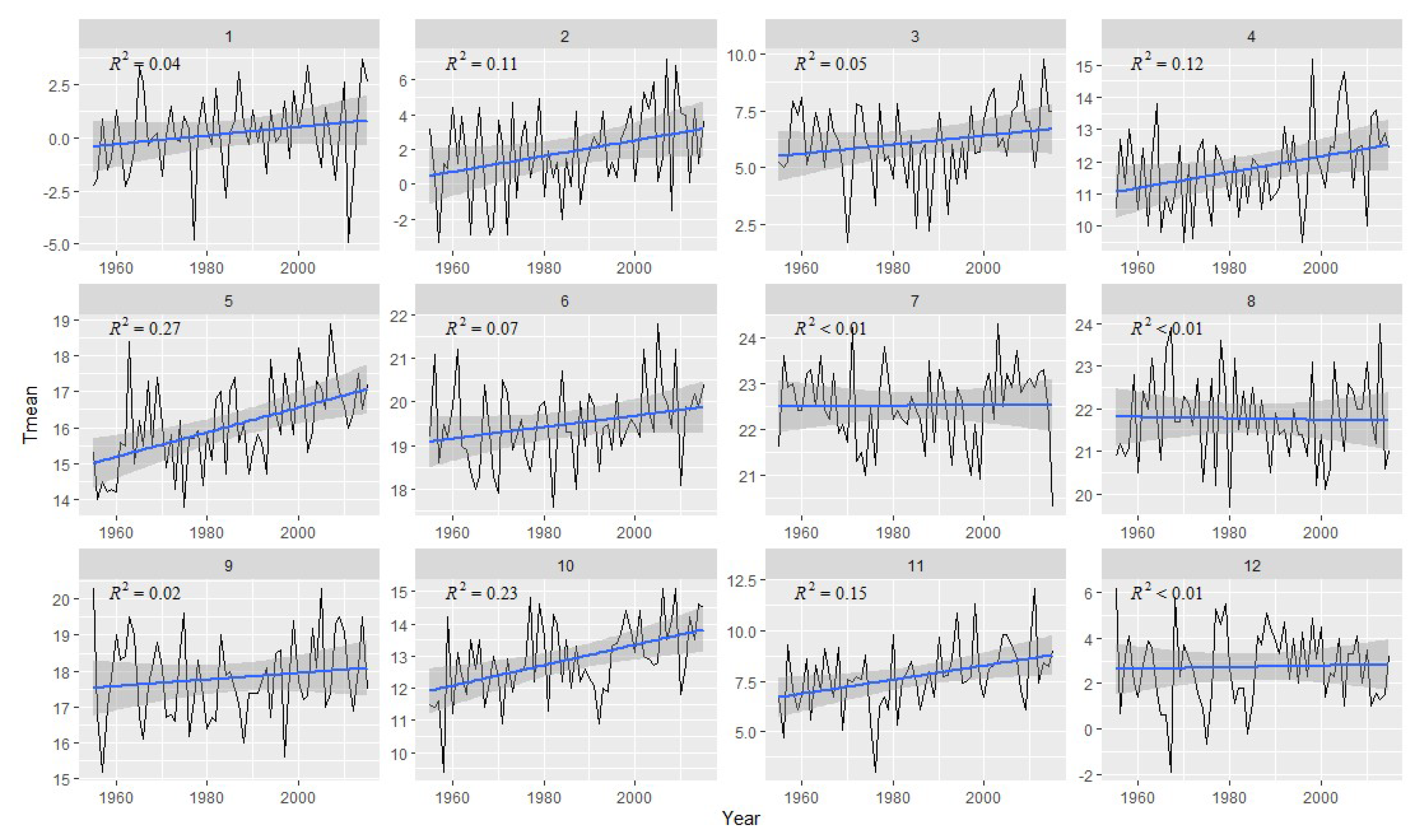
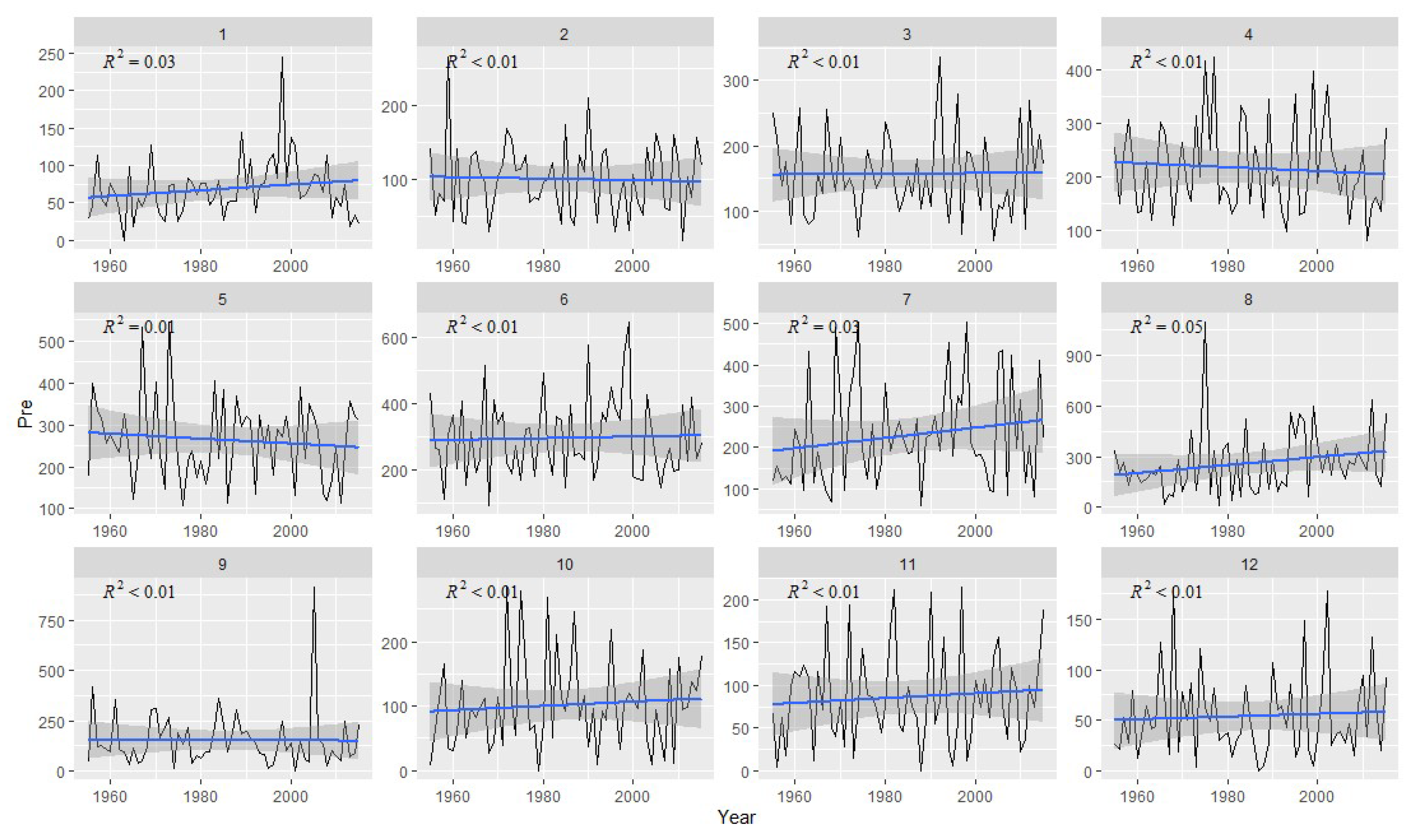
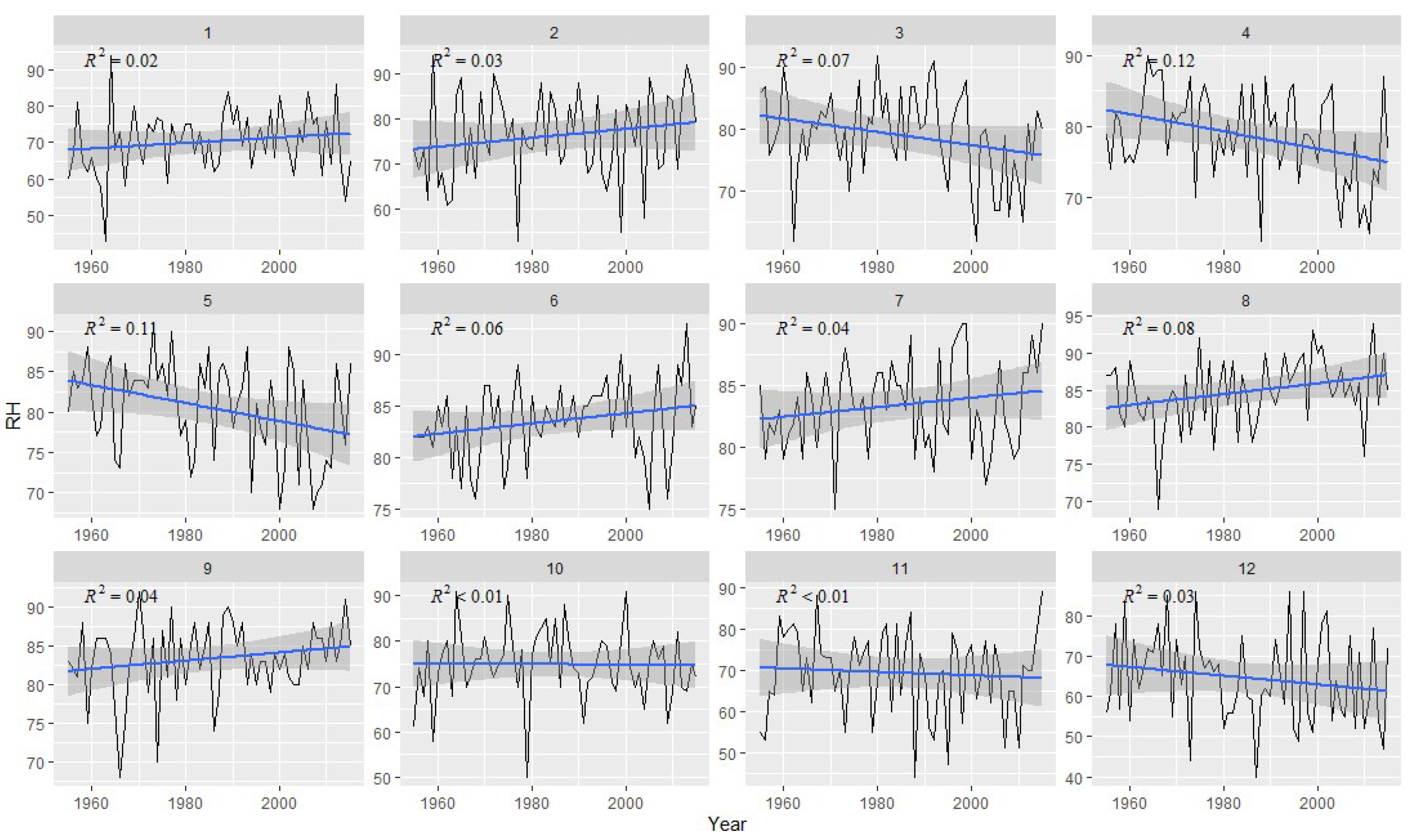
2.2. Meteorological Data
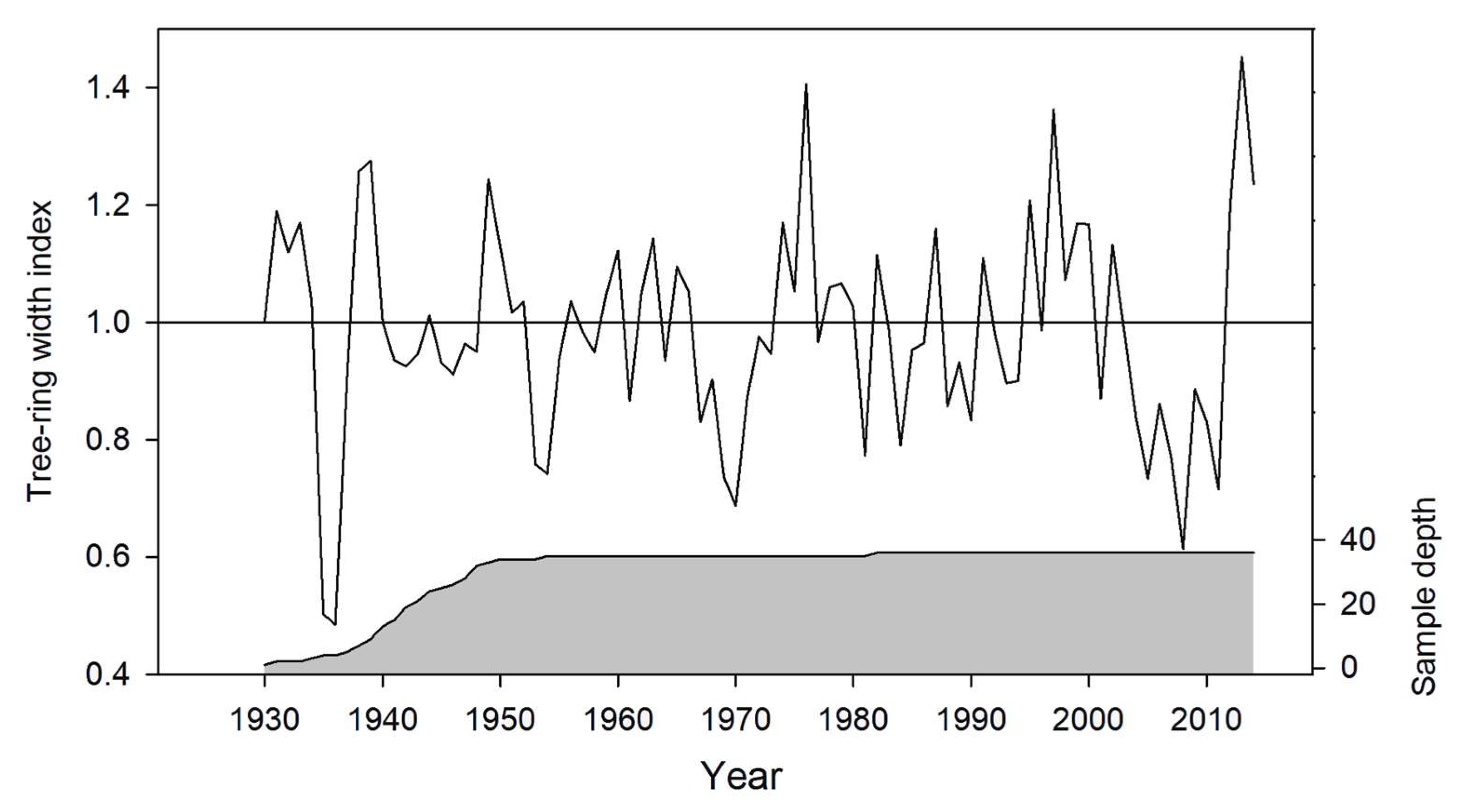
2.3. Tree-Ring Data
2.4. Statistical Analysis
3. Results
3.1. Statistics of Chronology
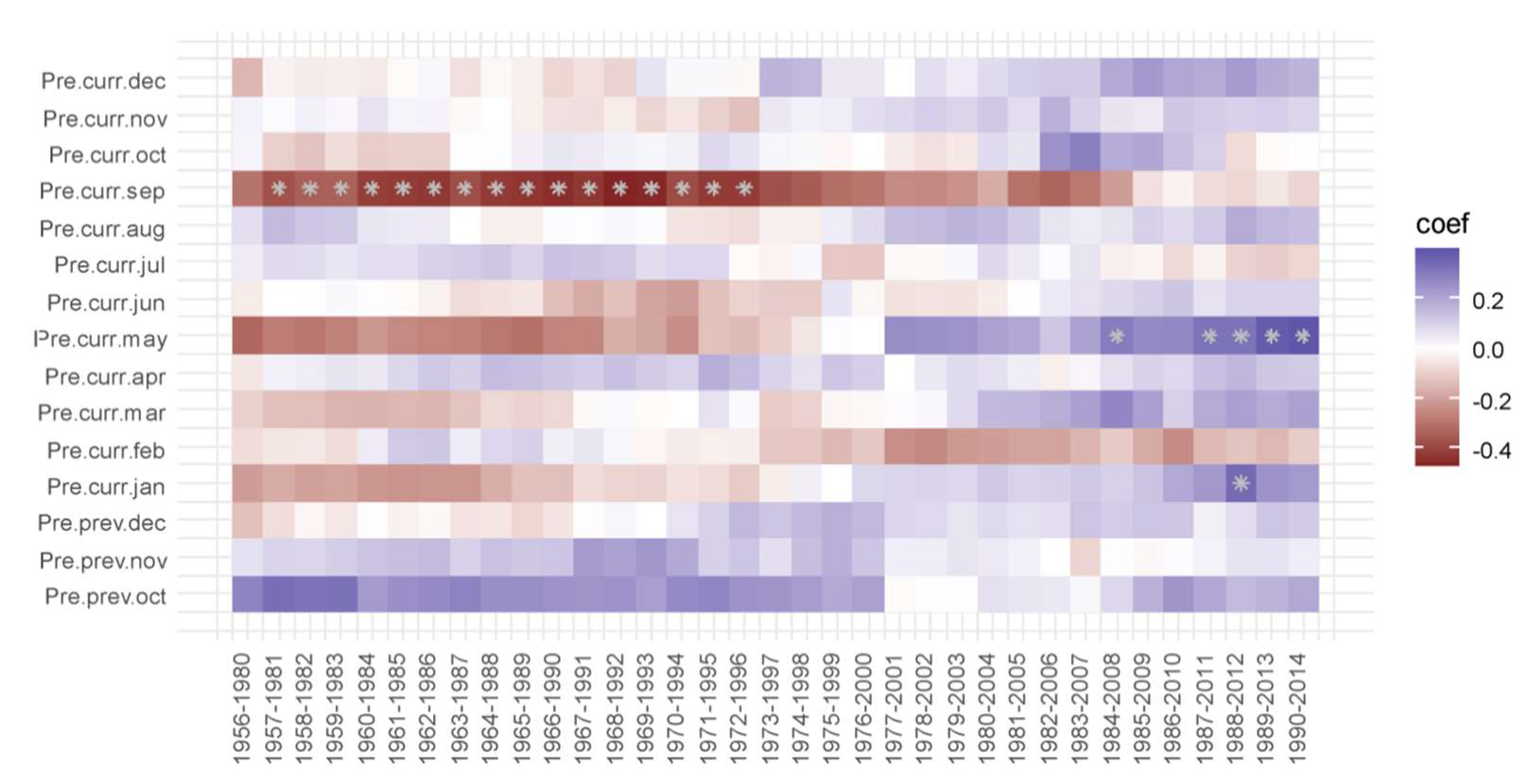
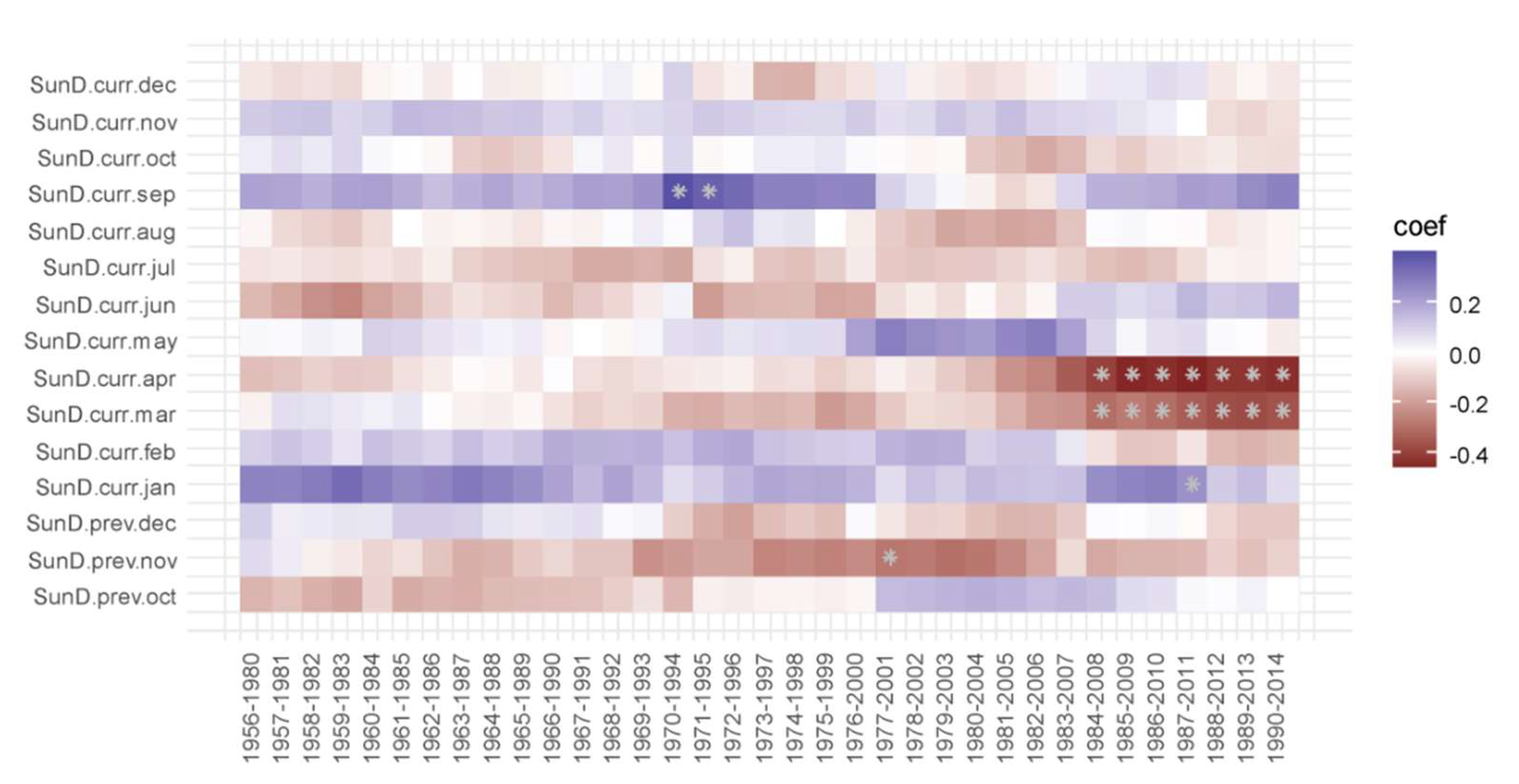
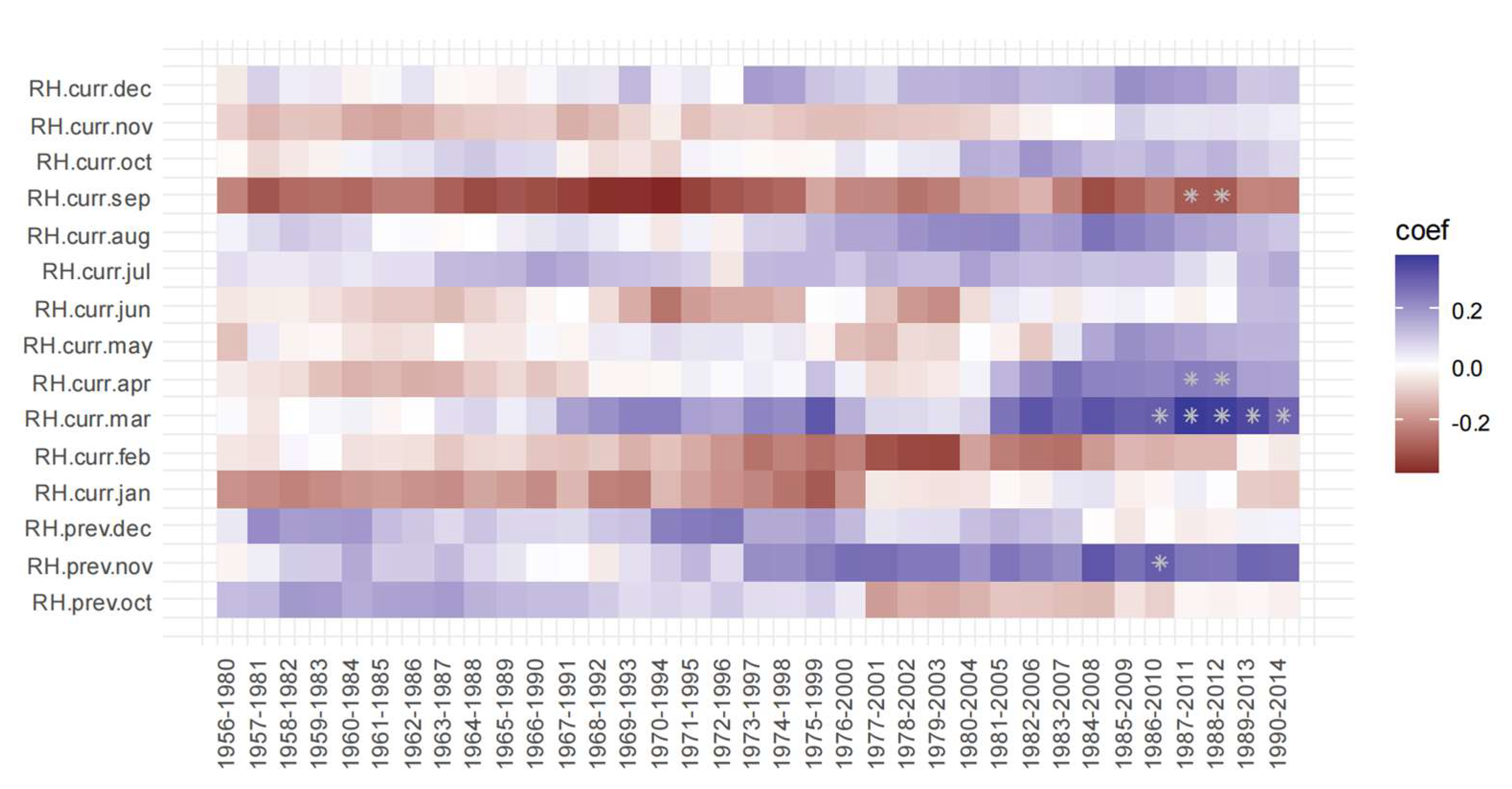
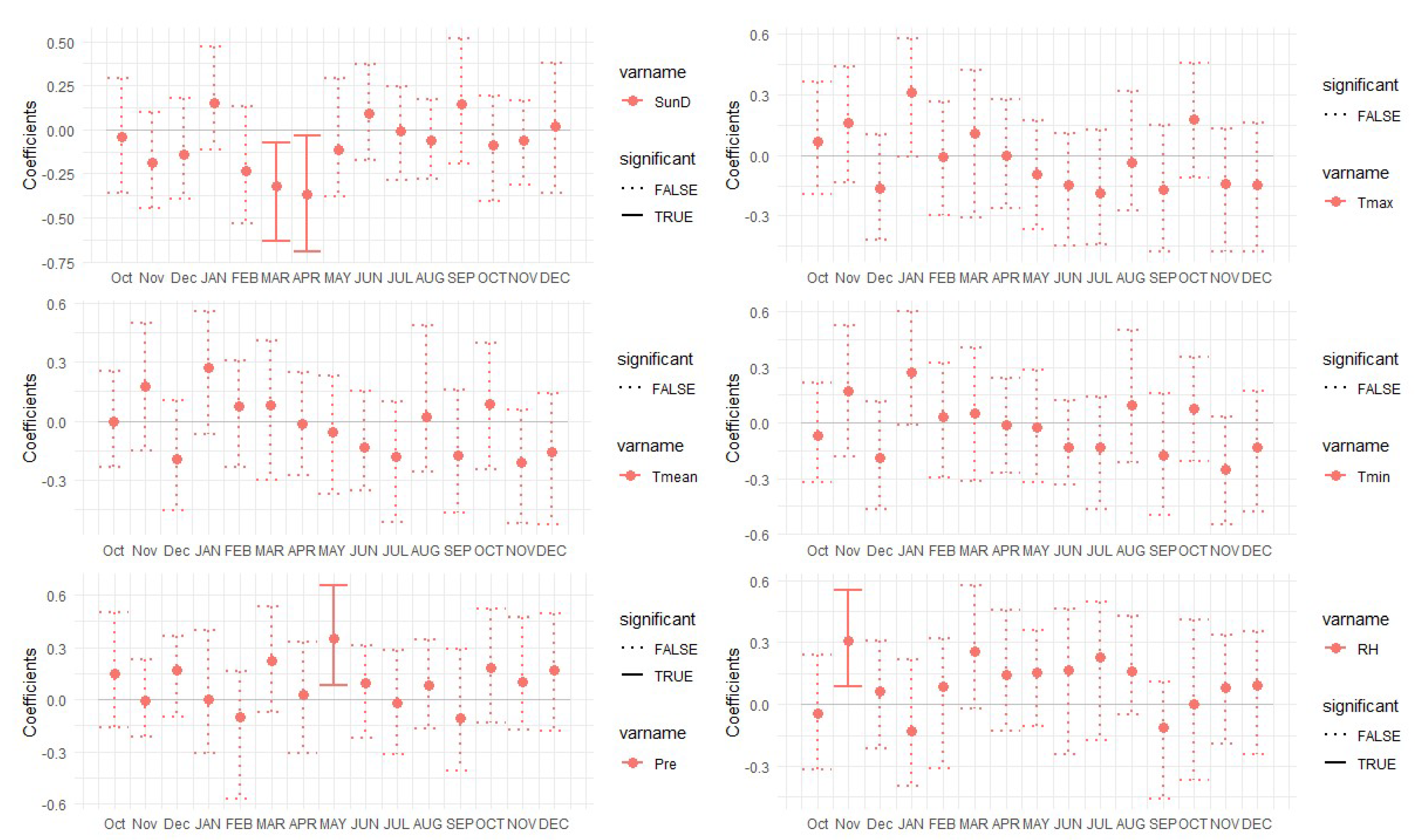
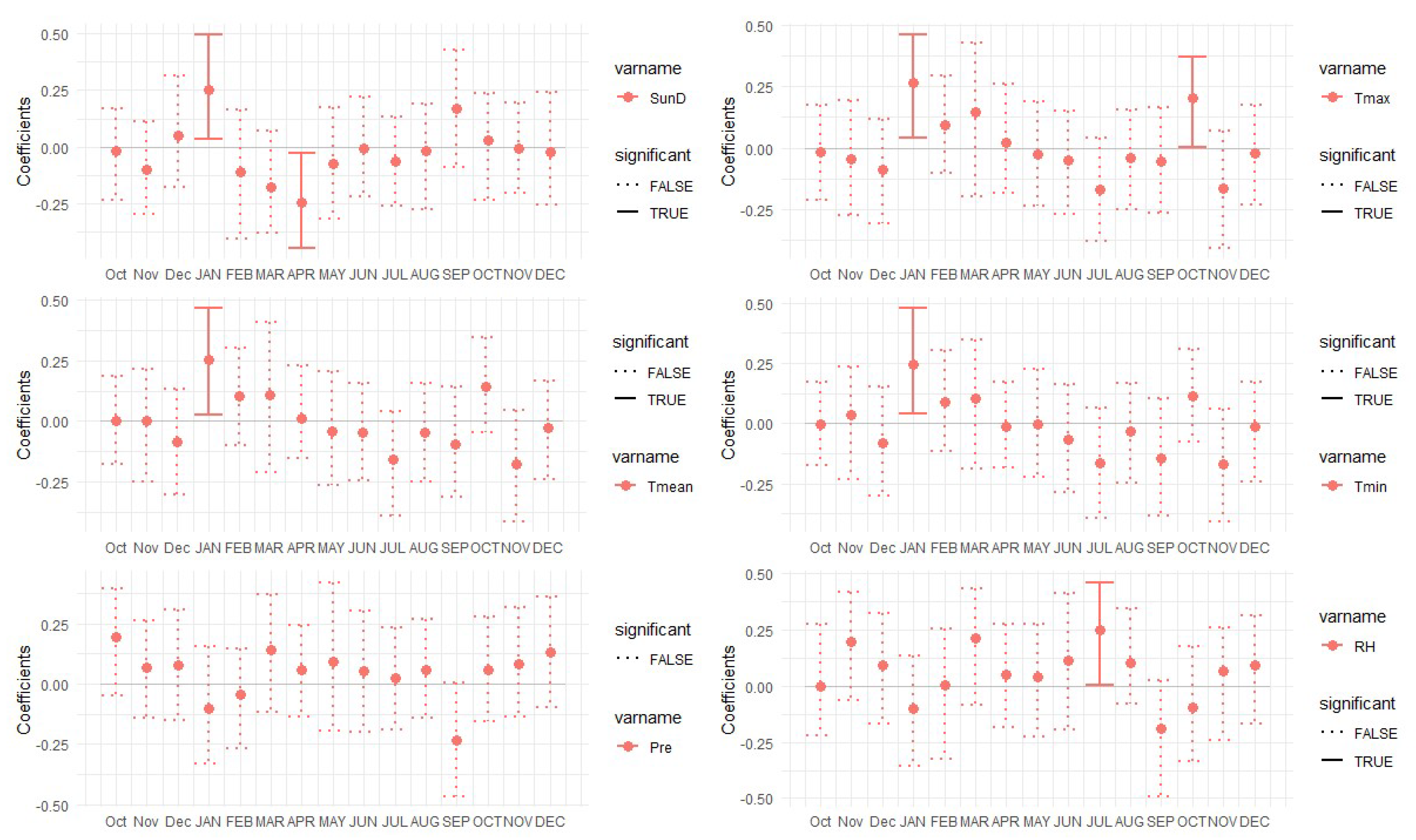
3.2. Correlation Analysis Results
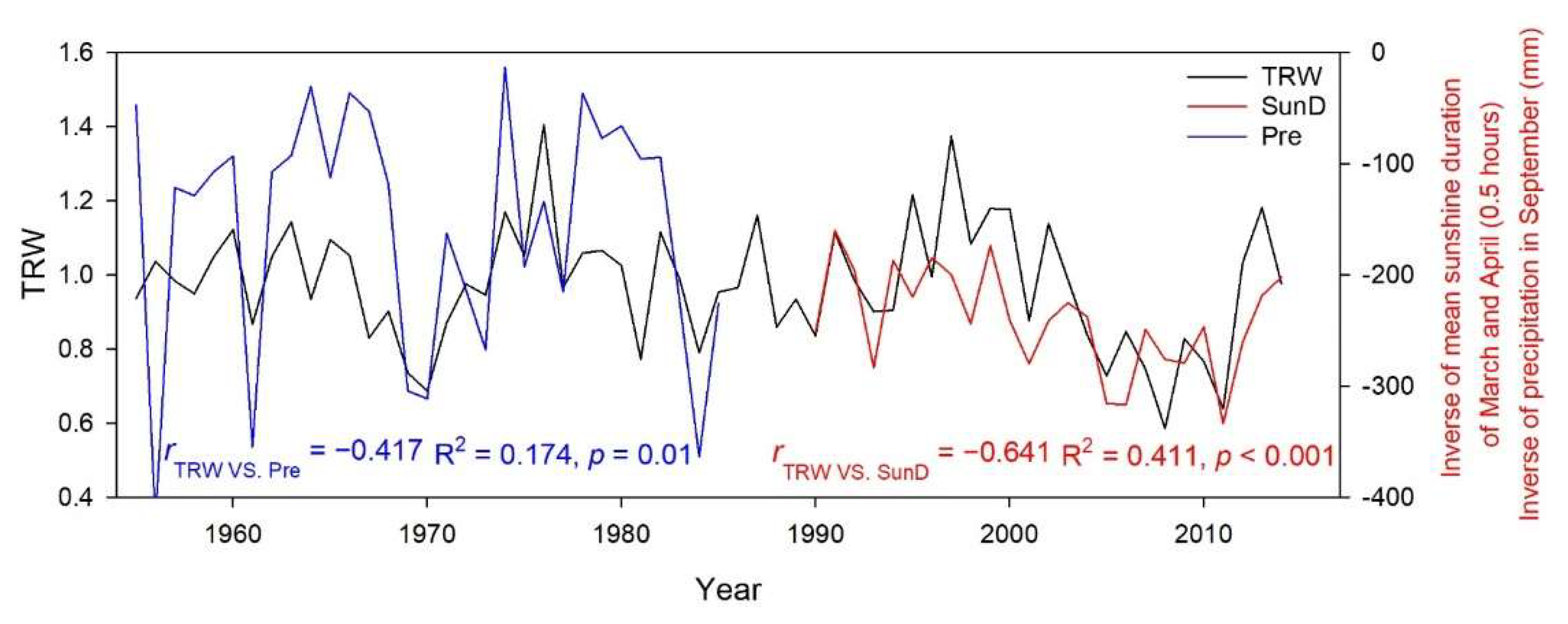
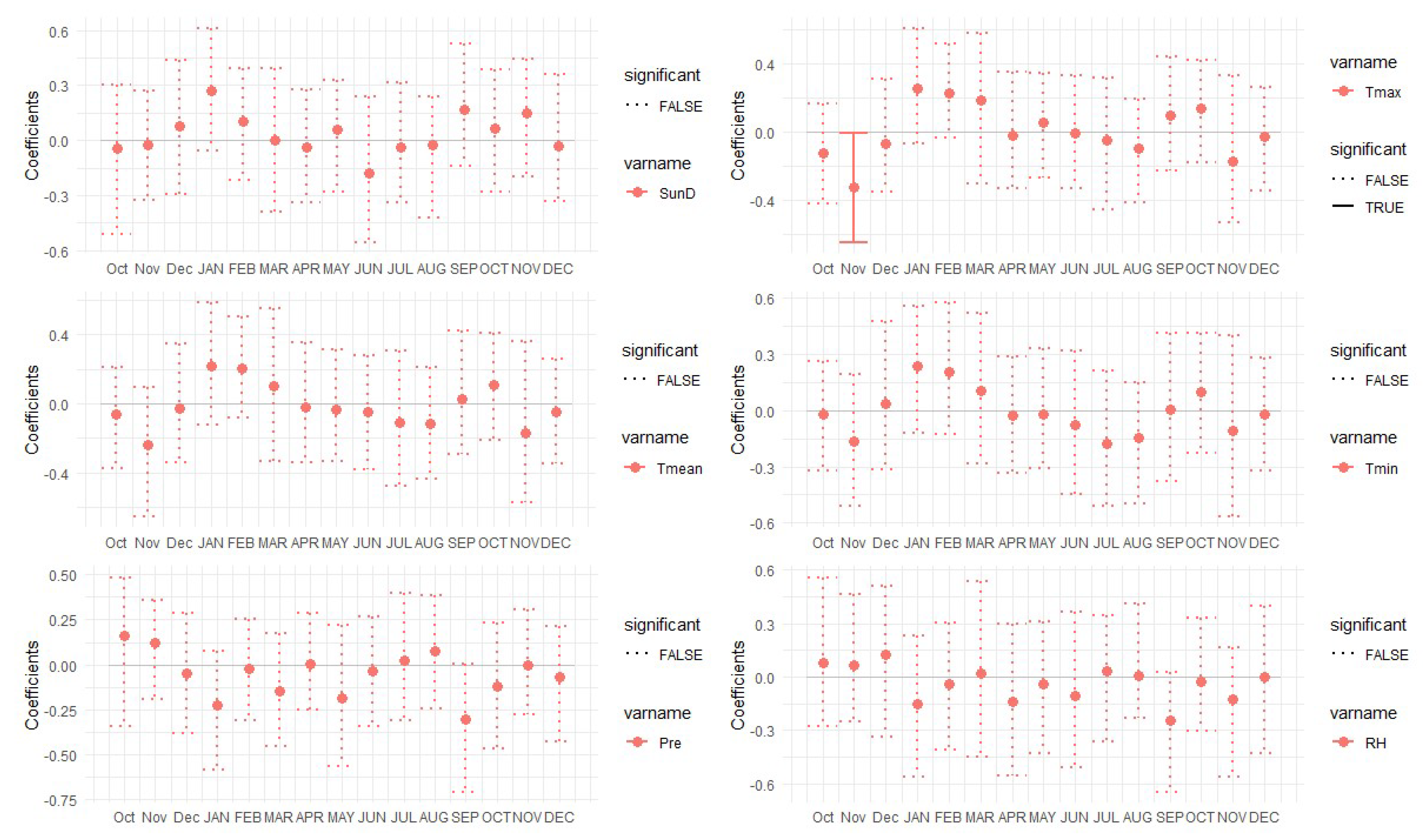
3.3. Linear Regression between TRW and Climate Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | TRW | Sample Depth | Year | TRW | Sample Depth |
|---|---|---|---|---|---|
| 1930 | 0.998957 | 1 | 1972 | 0.977521 | 35 |
| 1931 | 1.192125 | 2 | 1973 | 0.945807 | 35 |
| 1932 | 1.123558 | 2 | 1974 | 1.16978 | 35 |
| 1933 | 1.175056 | 2 | 1975 | 1.048711 | 35 |
| 1934 | 1.05426 | 3 | 1976 | 1.398248 | 35 |
| 1935 | 0.505486 | 4 | 1977 | 0.963854 | 35 |
| 1936 | 0.488438 | 4 | 1978 | 1.056816 | 35 |
| 1937 | 0.91265 | 5 | 1979 | 1.061208 | 35 |
| 1938 | 1.255427 | 7 | 1980 | 1.022838 | 35 |
| 1939 | 1.267796 | 9 | 1981 | 0.770513 | 35 |
| 1940 | 1.018137 | 13 | 1982 | 1.11466 | 36 |
| 1941 | 0.93703 | 15 | 1983 | 0.986659 | 36 |
| 1942 | 0.926429 | 19 | 1984 | 0.790929 | 36 |
| 1943 | 0.949574 | 21 | 1985 | 0.954859 | 36 |
| 1944 | 1.010751 | 24 | 1986 | 0.965173 | 36 |
| 1945 | 0.929943 | 25 | 1987 | 1.160598 | 36 |
| 1946 | 0.909081 | 26 | 1988 | 0.857617 | 36 |
| 1947 | 0.962927 | 28 | 1989 | 0.932668 | 36 |
| 1948 | 0.947237 | 32 | 1990 | 0.832178 | 36 |
| 1949 | 1.23344 | 33 | 1991 | 1.106878 | 36 |
| 1950 | 1.121501 | 34 | 1992 | 0.977515 | 36 |
| 1951 | 1.015939 | 34 | 1993 | 0.893167 | 36 |
| 1952 | 1.035711 | 34 | 1994 | 0.894887 | 36 |
| 1953 | 0.758882 | 34 | 1995 | 1.19978 | 36 |
| 1954 | 0.743368 | 35 | 1996 | 0.978716 | 36 |
| 1955 | 0.937655 | 35 | 1997 | 1.352266 | 36 |
| 1956 | 1.036171 | 35 | 1998 | 1.063549 | 36 |
| 1957 | 0.986152 | 35 | 1999 | 1.160516 | 36 |
| 1958 | 0.950814 | 35 | 2000 | 1.162281 | 36 |
| 1959 | 1.049728 | 35 | 2001 | 0.867065 | 36 |
| 1960 | 1.124773 | 35 | 2002 | 1.129707 | 36 |
| 1961 | 0.872579 | 35 | 2003 | 0.987388 | 36 |
| 1962 | 1.051172 | 35 | 2004 | 0.841395 | 36 |
| 1963 | 1.146505 | 35 | 2005 | 0.7391 | 36 |
| 1964 | 0.939582 | 35 | 2006 | 0.869069 | 36 |
| 1965 | 1.100443 | 35 | 2007 | 0.77514 | 36 |
| 1966 | 1.058614 | 35 | 2008 | 0.619257 | 36 |
| 1967 | 0.835391 | 35 | 2009 | 0.895106 | 36 |
| 1968 | 0.908211 | 35 | 2010 | 0.836877 | 36 |
| 1969 | 0.741137 | 35 | 2011 | 0.719122 | 36 |
| 1970 | 0.692645 | 35 | 2012 | 1.203209 | 36 |
| 1971 | 0.878336 | 35 | 2013 | 1.440285 | 36 |



References
- Briffa, K.R.; Schweingruber, F.H.; Jones, P.D.; Osborn, T.J.; Shiyatov, S.G.; Vaganov, E.A. Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature 1998, 391, 678–682. [Google Scholar] [CrossRef]
- Esper, J.; Cook, E.R.; Schweingruber, F.H. Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. Science 2002, 295, 2250–2253. [Google Scholar] [CrossRef] [PubMed]
- Liang, E.; Shao, X.; Qin, N. Tree-ring based summer temperature reconstruction for the source region of the Yangtze River on the Tibetan Plateau. Glob. Planet. Chang. 2008, 61, 313–320. [Google Scholar] [CrossRef]
- Lyu, L.; Büntgen, U.; Treydte, K.; Yu, K.; Liang, H.; Reinig, F.; Nievergelt, D.; Li, M.-H.; Cherubini, P. Tree-rings reveal hydroclimatic fingerprints of the Pacific Decadal Oscillation on the Tibetan Plateau. Clim. Dyn. 2019, 53, 1023–1037. [Google Scholar] [CrossRef]
- Yang, B.; Qin, C.; Wang, J.; He, M.; Melvin, T.M.; Osborn, T.J.; Briffa, K.R. A 3500-year tree-ring record of annual precipitation on the northeastern Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2014, 111, 2903–2908. [Google Scholar] [CrossRef] [PubMed]
- Fritts, H. Tree-Rings and Climate; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- D’Arrigo, R.; Wilson, R.; Liepert, B.; Cherubini, P. On the ‘Divergence Problem’ in Northern Forests: A review of the tree-ring evidence and possible causes. Glob. Planet. Chang. 2008, 60, 289–305. [Google Scholar] [CrossRef]
- Briffa, K.R.; Bartholin, T.S.; Eckstein, D.; Jones, P.D.; Karlén, W.; Schweingruber, F.H.; Zetterberg, P. A 1400-year tree-ring record of summer temperatures in Fennoscandia. Nature 1990, 346, 434–439. [Google Scholar] [CrossRef]
- Bastida, J.M.; Rey, P.J.; Alcántara, J.M. Local adaptation to distinct elevational cores contributes to current elevational divergence of two Aquilegia vulgaris subspecies. J. Plant Ecol. 2014, 8, 273–283. [Google Scholar] [CrossRef]
- Büntgen, U.; Frank, D.C.; Schmidhalter, M.; Neuwirth, B.; Seifert, M.; Esper, J. Growth/climate response shift in a long subalpine spruce chronology. Trees 2006, 20, 99–110. [Google Scholar] [CrossRef]
- Franceschini, T.; Bontemps, J.-D.; Perez, V.; Leban, J.-M. Divergence in latewood density response of Norway spruce to temperature is not resolved by enlarged sets of climatic predictors and their non-linearities. Agric. For. Meteorol. 2013, 180, 132–141. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Y.; Song, H.; Yang, Y.; Zhao, B. Divergence of tree-ring-based drought reconstruction between the individual sampling site and the Monsoon Asia Drought Atlas: An example from Guancen Mountain. Sci. Bull. 2015, 60, 1688–1697. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Ji, Y.; Li, Z.; Li, M.; Zhang, Y. Temperature signals in tree-ring width and divergent growth of Korean pine response to recent climate warming in northeast Asia. Trees 2016, 31, 415–427. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Age-dependent tree-ring growth responses to climate in Larix decidua and Pinus uncinata. Ecology 2004, 85, 730–740. [Google Scholar] [CrossRef]
- Čermák, P.; Rybníček, M.; Žid, T.; Steffenrem, A.; Kolář, T. Site and age-dependent responses of Picea abies growth to climate variability. Eur. J. For. Res. 2019, 138, 445–460. [Google Scholar] [CrossRef]
- Konter, O.; Büntgen, U.; Carrer, M.; Timonen, M.; Esper, J. Climate signal age effects in boreal tree-rings: Lessons to be learned for paleoclimatic reconstructions. Quaternary. Sci. Rev. 2016, 142, 164–172. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carrer, M. Age-dependent xylogenesis in timberline conifers. New Phytol. 2008, 177, 199–208. [Google Scholar] [CrossRef]
- Li, X.; Liang, E.; Gričar, J.; Prislan, P.; Rossi, S.; Čufar, K. Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau. Tree Physiol. 2012, 33, 48–56. [Google Scholar] [CrossRef]
- Wu, G.; Xu, G.; Chen, T.; Liu, X.; Zhang, Y.; An, W.; Wang, W.; Fang, Z.-A.; Yu, S. Age-dependent tree-ring growth responses of Schrenk spruce (Picea schrenkiana) to climate—A case study in the Tianshan Mountain, China. Dendrochronologia 2013, 31, 318–326. [Google Scholar] [CrossRef]
- Xu, G.; Wu, G.; Liu, X.; Chen, T.; Wang, B.; Hudson, A.; Trouet, V. Age-related climate response of tree-ring δ13C and δ18O from spruce in northwestern China, with implications for relative humidity reconstructions. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005513. [Google Scholar]
- Yu, G.; Chen, Z.; Piao, S.; Peng, C.; Ciais, P.; Wang, Q.; Li, X.; Zhu, X. High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region. Proc. Natl. Acad. Sci. USA 2014, 111, 4910–4915. [Google Scholar] [CrossRef]
- Duan, J.; Zhang, Q.-B.; Lv, L.; Zhang, C. Regional-scale winter-spring temperature variability and chilling damage dynamics over the past two centuries in southeastern China. Clim. Dyn. 2012, 39, 919–928. [Google Scholar] [CrossRef]
- Shi, J.; Lu, H.; Li, J.; Shi, S.; Wu, S.; Hou, X.; Li, L. Tree-ring based February-April precipitation reconstruction for the lower reaches of the Yangtze River, southeastern China. Glob. Planet. Chang. 2015, 131, 82–88. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y.; Fang, C.; Zhang, H.; Song, H.; Li, Q.; Sun, C.; Wan, C.; Liu, R. Ground surface temperature reconstruction for the Jinggangshan Mountains: Interpreting the hydro-thermal coupling pattern in southeastern China. Quaternary. Sci. Rev. 2020, 249, 106591. [Google Scholar] [CrossRef]
- Liang, H.; Huang, J.-G.; Ma, Q.; Li, J.; Wang, Z.; Guo, X.; Zhu, H.; Jiang, S.; Zhou, P.; Yu, B.; et al. Contributions of competition and climate on radial growth of Pinus massoniana in subtropics of China. Agric. For. Meteorol. 2019, 274, 7–17. [Google Scholar] [CrossRef]
- Huang, X.; Dai, D.; Xiang, Y.; Yan, Z.; Teng, M.; Wang, P.; Zhou, Z.; Zeng, L.; Xiao, W. Radial growth of Pinus massoniana is influenced by temperature, precipitation, and site conditions on the regional scale: A meta-analysis based on tree-ring width index. Ecol. Indic. 2021, 126, 107659. [Google Scholar] [CrossRef]
- Su, J.; Gou, X.; HilleRisLambers, J.; Deng, Y.; Fan, H.; Zheng, W.; Zhang, R.; Manzanedo, R.D. Increasing climate sensitivity of subtropical conifers along an aridity gradient. For. Ecol. Manag. 2021, 482, 118841. [Google Scholar] [CrossRef]
- Yang, R.-Q.; Fu, P.-L.; Fan, Z.-X.; Panthi, S.; Gao, J.; Niu, Y.; Li, Z.-S.; Bräuning, A. Growth-climate sensitivity of two pine species shows species-specific changes along temperature and moisture gradients in southwest China. Agric. For. Meteorol. 2022, 318, 108907. [Google Scholar] [CrossRef]
- Zuidema, P.A.; Babst, F.; Groenendijk, P.; Trouet, V.; Abiyu, A.; Acuña-Soto, R.; Adenesky-Filho, E.; Alfaro-Sánchez, R.; Aragão, J.R.V.; Assis-Pereira, G.; et al. Tropical tree growth driven by dry-season climate variability. Nat. Geosci. 2022, 15, 269–276. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y.; Liu, H.; Sun, C.; Wang, Y. Growing-season precipitation since 1872 in the coastal area of subtropical southeast China reconstructed from tree-rings and its relationship with the East Asian summer monsoon system. Ecol. Indic. 2017, 82, 441–450. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. Treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. URL. R Foundation for Statistical Computing, Vienna, Austria. 2015. Available online: https://www.R-project.org/ (accessed on 13 June 2022).
- Buras, A. A comment on the expressed population signal. Dendrochronologia 2017, 44, 130–132. [Google Scholar] [CrossRef]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Appl. Meteorol. Climatol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Wilson, R.; D’Arrigo, R.; Buckley, B.; Büntgen, U.; Esper, J.; Frank, D.; Luckman, B.; Payette, S.; Vose, R.; Youngblut, D. A matter of divergence: Tracking recent warming at hemispheric scales using tree-ring data. J. Geophys. Res. Atmos. 2007, 112, 107–114. [Google Scholar] [CrossRef]
- Trouillier, M.; van der Maaten-Theunissen, M.; Scharnweber, T.; Würth, D.; Burger, A.; Schnittler, M.; Wilmking, M. Size matters—A comparison of three methods to assess age- and size-dependent climate sensitivity of trees. Trees 2018, 33, 183–192. [Google Scholar] [CrossRef]
- Cai, Q.F.; Liu, Y. Two centuries temperature variations over subtropical southeast China inferred from Pinus taiwanensis Hayata tree-ring width. Clim. Dyn. 2017, 48, 1813–1825. [Google Scholar] [CrossRef]
- Huang, J.G.; Guo, X.; Rossi, S.; Zhai, L.; Yu, B.; Zhang, S.; Zhang, M. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season. Tree. Physiol. 2018, 38, 1225–1236. [Google Scholar] [CrossRef]
- Liang, H.; Lyu, L.; Wahab, M. A 382-year reconstruction of August mean minimum temperature from tree-ring maximum latewood density on the southeastern Tibetan Plateau, China. Dendrochronologia 2016, 37, 1–8. [Google Scholar] [CrossRef]
- Panthi, S.; Bräuning, A.; Zhou, Z.K.; Fan, Z.X. Growth response of Abies georgei to climate increases with elevation in the central Hengduan Mountains, southwestern China. Dendrochronologia 2018, 47, 1–9. [Google Scholar] [CrossRef]
- Fan, Z.X.; Brauning, A.; Cao, K.F. Tree-ring based drought reconstruction in the central Hengduan Mountains region (China) since AD 1655. Int. J. Climatol. 2008, 28, 1879–1887. [Google Scholar] [CrossRef]
- Sano, M.; Buckley, B.M.; Sweda, T. Tree-ring based hydroclimate reconstruction over northern Vietnam from Fokienia hodginsii: Eighteenth century mega-drought and tropical Pacific influence. Clim. Dyn. 2009, 33, 331–340. [Google Scholar] [CrossRef]
- Esper, J.; Niederer, R.; Bebi, P.; Frank, D. Climate signal age effects—Evidence from young and old trees in the Swiss Engadin. For. Ecol. Manag. 2008, 255, 3783–3789. [Google Scholar] [CrossRef]
- Linares, J.C.; Taïqui, L.; Sangüesa-Barreda, G.; Seco, J.I.; Camarero, J.J. Age-related drought sensitivity of Atlas cedar (Cedrus atlantica) in the Moroccan Middle Atlas forests. Dendrochronologia 2013, 31, 88–96. [Google Scholar] [CrossRef]
- Yu, G.; Liu, Y.; Wang, X.; Ma, K. Age-dependent tree-ring growth responses to climate in Qilian juniper (Sabina przewalskii Kom.). Trees 2008, 22, 197–204. [Google Scholar] [CrossRef]
- Gomez-Aparicio, L.O.; Garcia-Valdes, R.A.; Ruiz-Benito, P.A.; Zavala, M.A. Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change. Glob. Chang. Biol. 2011, 17, 2400–2414. [Google Scholar] [CrossRef] [Green Version]







| September Precipitation | Mean Sunshine Duration from March to April | ||||
|---|---|---|---|---|---|
| Time Span | r | R-Square | Time Span | r | R-Square |
| 1956–1976 | −0.379 | 0.144 | 1976–2014 | −0.535 | 0.286 |
| 1956–1977 | −0.380 | 0.144 | 1977–2014 | −0.491 | 0.241 |
| 1956–1978 | −0.390 | 0.152 | 1978–2014 | −0.491 | 0.241 |
| 1956–1979 | −0.398 | 0.159 | 1979–2014 | −0.516 | 0.267 |
| 1956–1980 | −0.400 | 0.160 | 1980–2014 | −0.545 | 0.297 |
| 1956–1981 | −0.354 | 0.125 | 1981–2014 | −0.545 | 0.297 |
| 1956–1982 | −0.362 | 0.131 | 1982–2014 | −0.588 | 0.346 |
| 1956–1983 | −0.359 | 0.129 | 1983–2014 | −0.606 | 0.368 |
| 1956–1984 | −0.416 | 0.173 | 1984–2014 | −0.624 | 0.390 |
| 1956–1985 | −0.417 | 0.174 | 1985–2014 | −0.624 | 0.389 |
| 1956–1986 | −0.413 | 0.170 | 1986–2014 | −0.630 | 0.396 |
| 1956–1987 | −0.389 | 0.151 | 1987–2014 | −0.630 | 0.396 |
| 1956–1988 | −0.410 | 0.168 | 1988–2014 | −0.626 | 0.392 |
| 1956–1989 | −0.412 | 0.169 | 1989–2014 | −0.622 | 0.387 |
| 1956–1990 | −0.414 | 0.172 | 1990–2014 | −0.641 | 0.411 |
| 1956–1991 | −0.407 | 0.166 | 1991–2014 | −0.641 | 0.411 |
| 1956–1992 | −0.407 | 0.166 | 1992–2014 | −0.633 | 0.401 |
| 1956–1993 | −0.392 | 0.153 | 1993–2014 | −0.645 | 0.416 |
| 1956–1994 | −0.376 | 0.141 | 1994–2014 | −0.646 | 0.418 |
| 1956–1995 | −0.413 | 0.170 | 1995–2014 | −0.696 | 0.484 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, H.; Wang, Z.; Jiang, S.; Zhang, C.; Huang, J. Temporal Changes in Growth–Climate Relationship of Pinus taiwanensis Hayata in Subtropical China. Forests 2022, 13, 1502. https://doi.org/10.3390/f13091502
Liang H, Wang Z, Jiang S, Zhang C, Huang J. Temporal Changes in Growth–Climate Relationship of Pinus taiwanensis Hayata in Subtropical China. Forests. 2022; 13(9):1502. https://doi.org/10.3390/f13091502
Chicago/Turabian StyleLiang, Hanxue, Zhou Wang, Shaowei Jiang, Cun Zhang, and Jianguo Huang. 2022. "Temporal Changes in Growth–Climate Relationship of Pinus taiwanensis Hayata in Subtropical China" Forests 13, no. 9: 1502. https://doi.org/10.3390/f13091502