
Intra-Annual Growth and Its Response to Climatic Factors of Two Salix Species under Warm Temperate Environment

Abstract
:1. Introduction
2. Materials and Methods
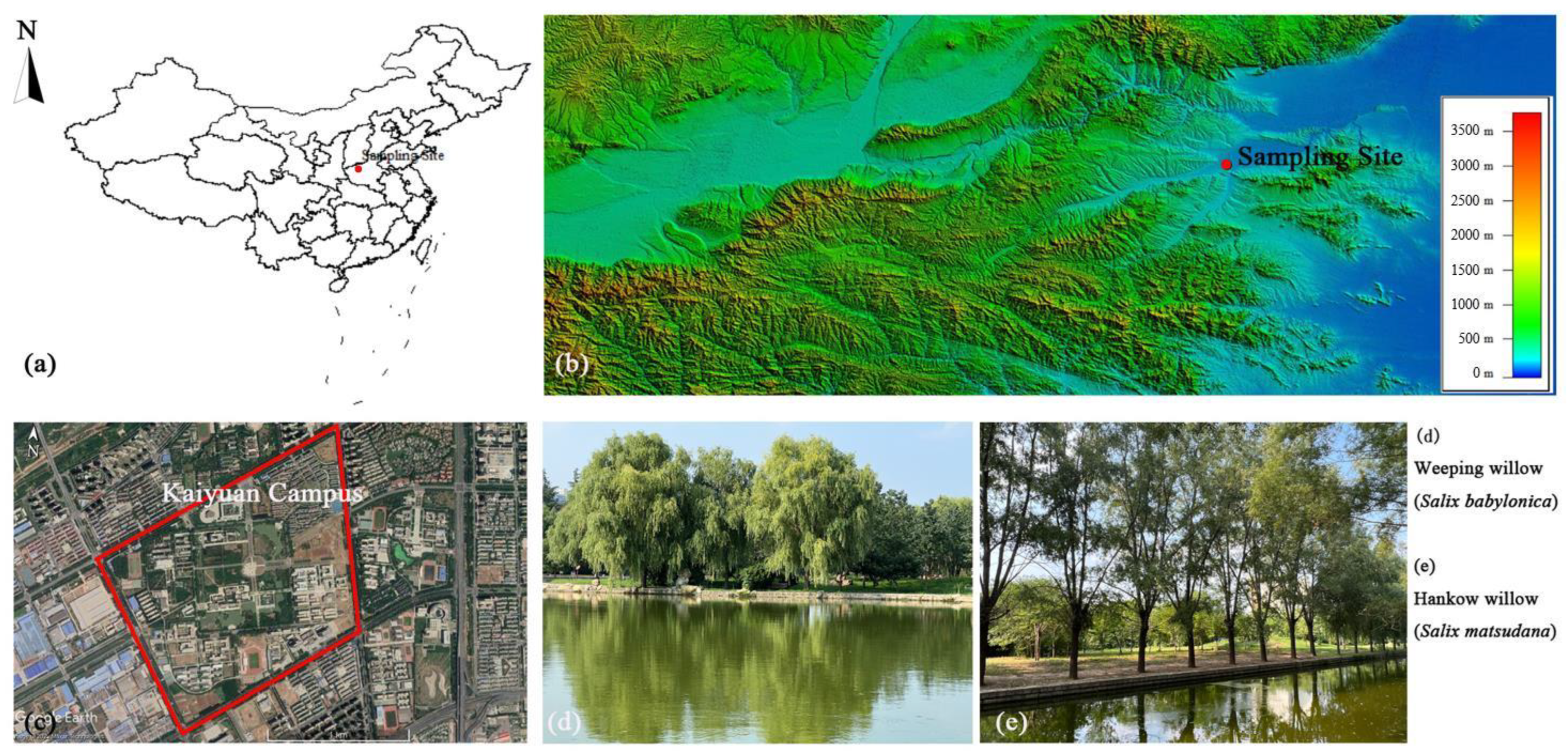
2.1. Study Site
2.2. Tree Selection and Lab Preparation
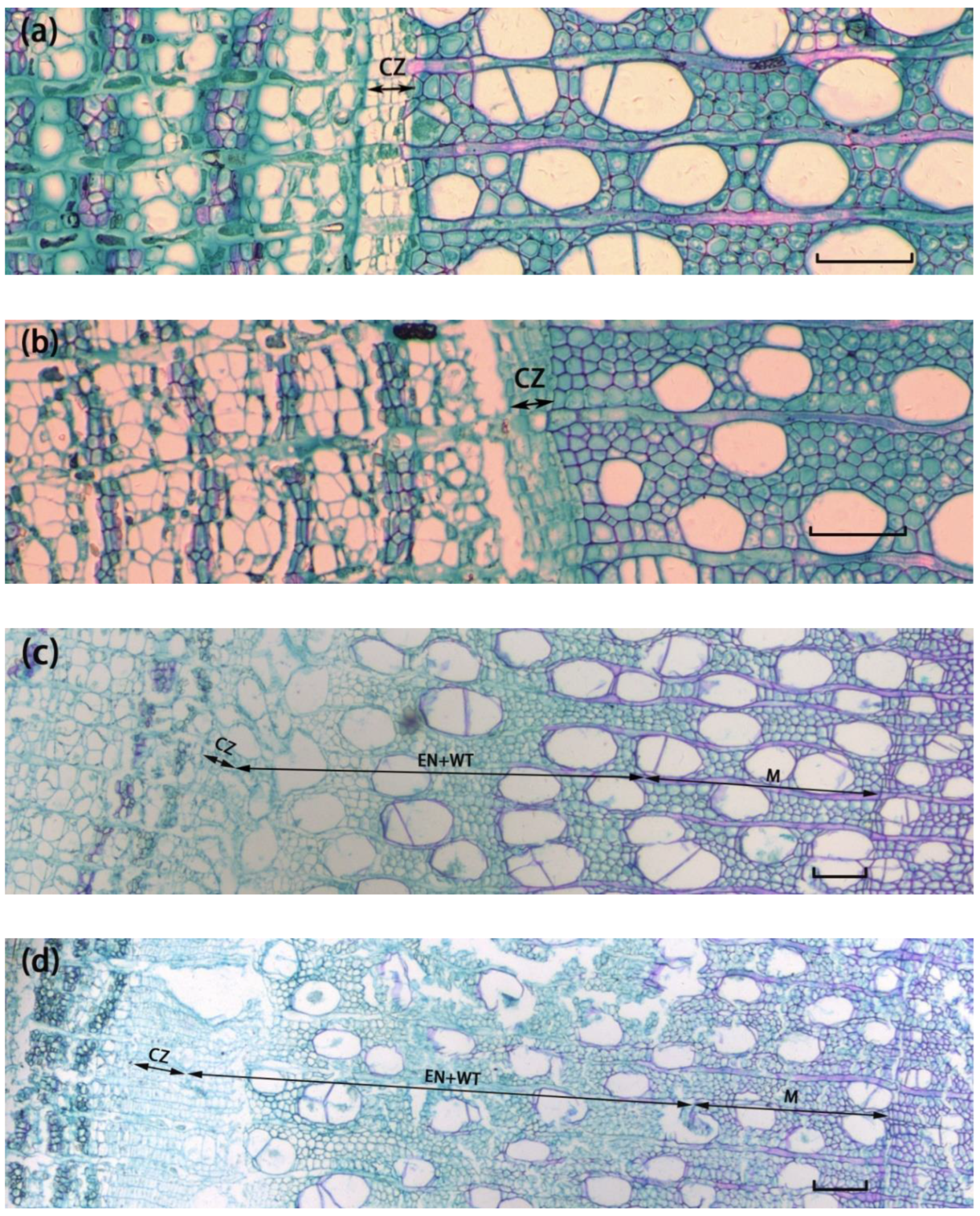
2.3. Cambial Activity and Xylem Differentiation
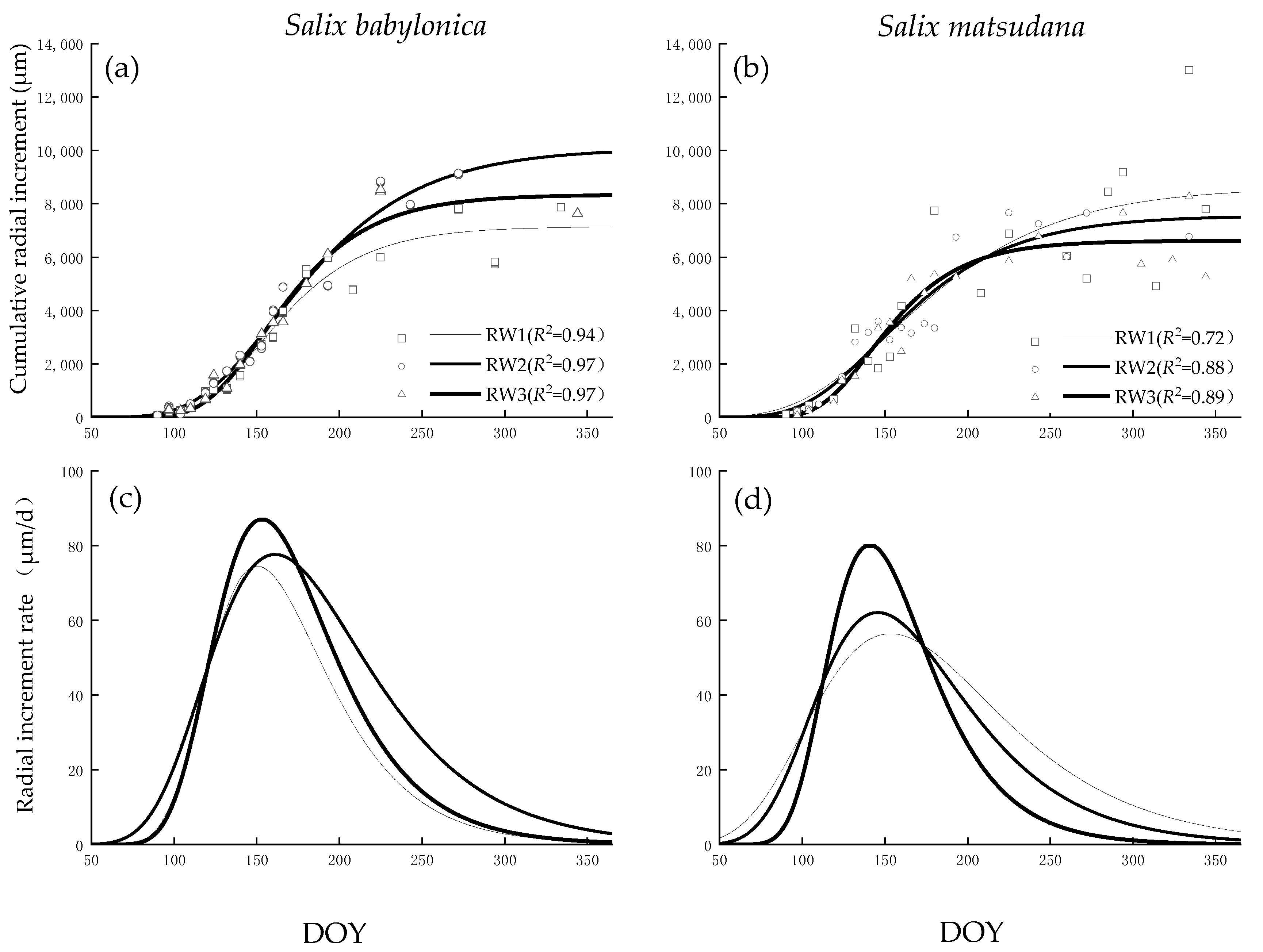
2.4. Intra-Annual Xylem Growth Fitting
2.5. Climate and Growth Relationship Analysis
3. Results
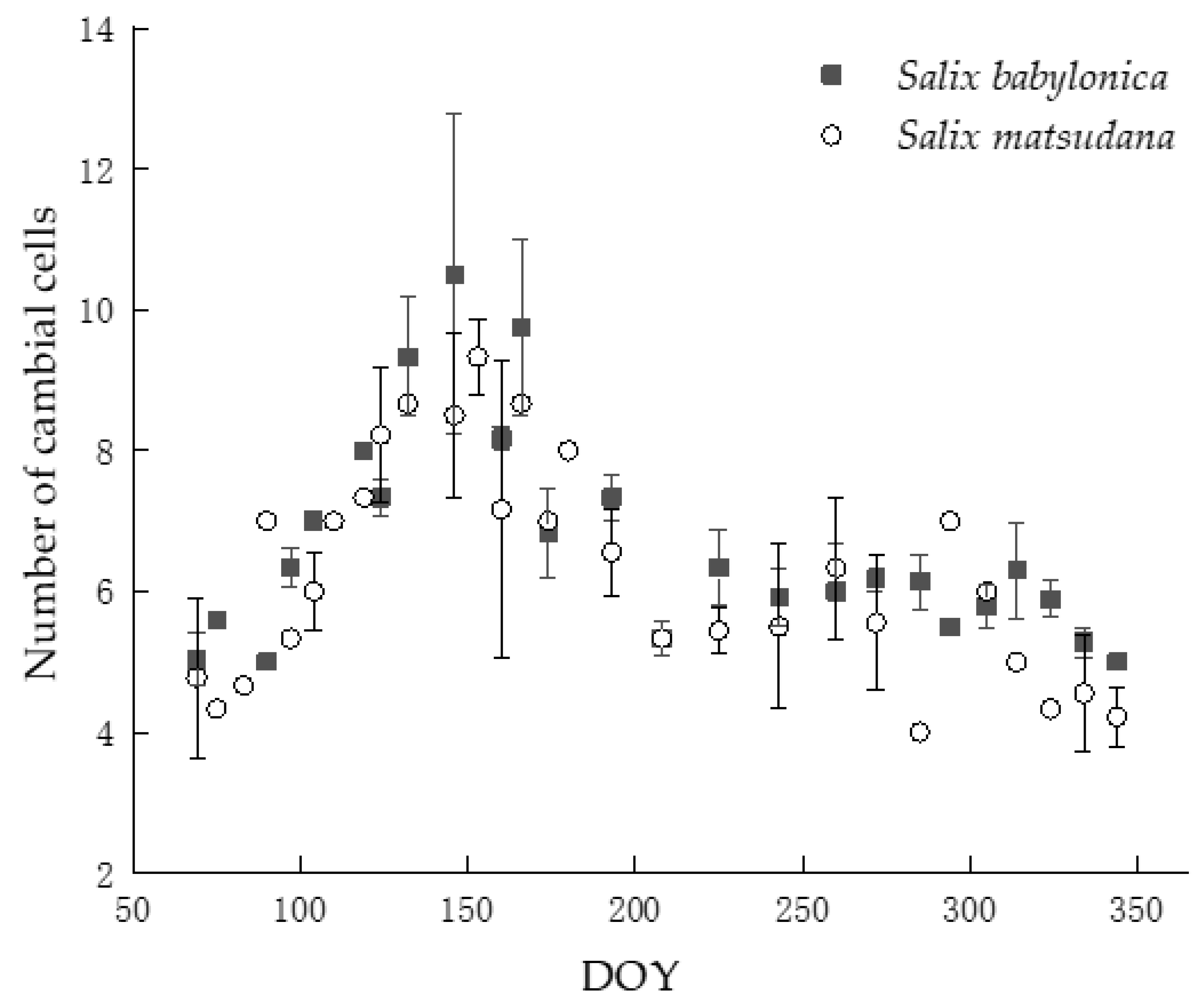
3.1. Cambial Activity
3.2. Xylem Differentiation
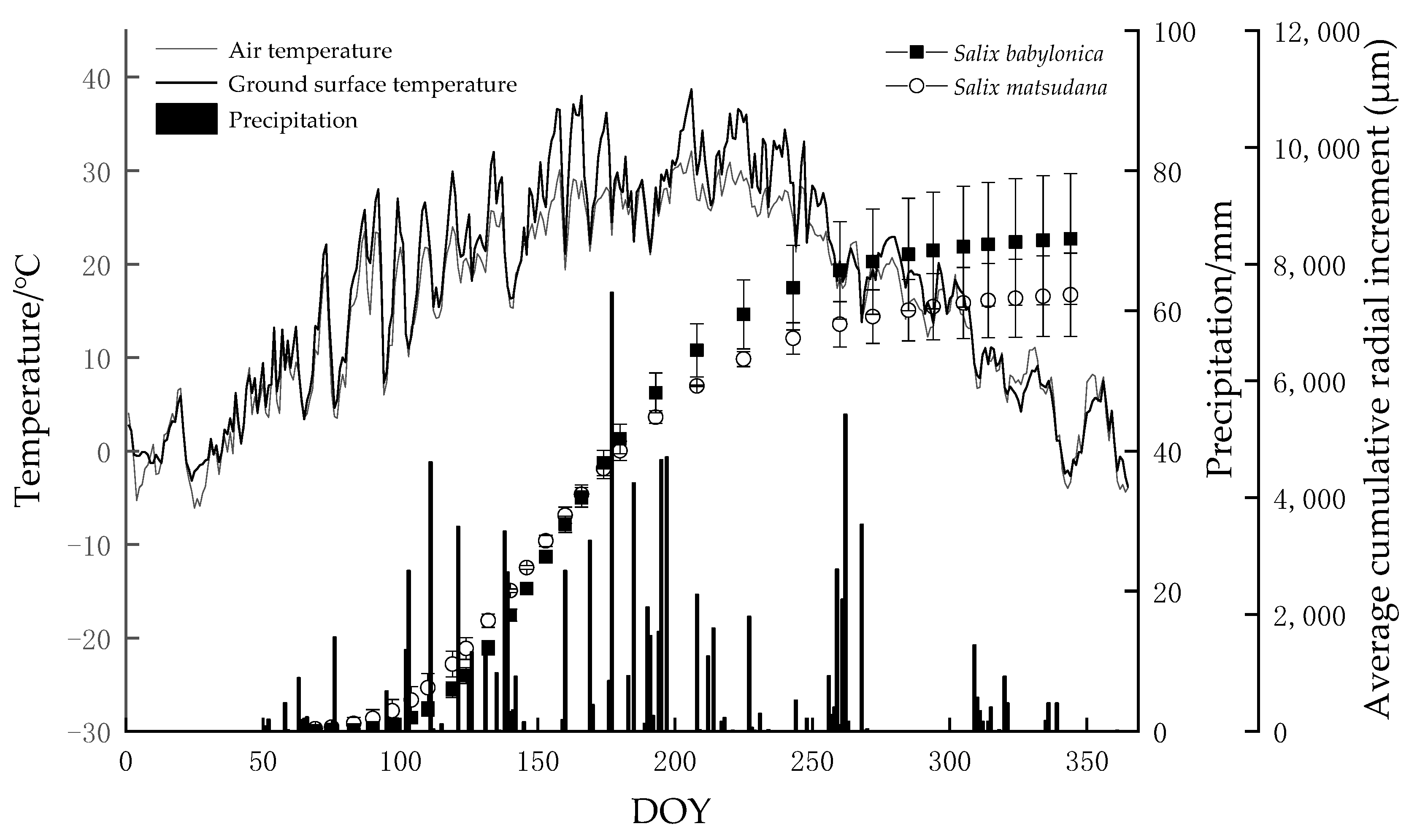
3.3. Xylem Growth Comparison
3.4. Climate–Xylem Growth Relationship
4. Discussion
4.1. Cambial Activity of Two Willows in Warm Temperate Zone
4.2. Xylem Growth with High Growth Rate
4.3. Higher Climate Growth Sensitivity in Salix matsudana Than Salix babylonica
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cuny, H.E.; Rathgeber, C.; Frank, D.; Fonti, P.; Mkinen, H.; Prislan, P.; Rossi, S.; Castillo, E.D.; Campelo, F.; Vavrík, H. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 2015, 1, 15160. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The physical science basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar] [CrossRef]
- Peters, R.L.; Steppe, K.; Cuny, H.E.; De Pauw, D.J.W.; Frank, D.C.; Schaub, M.; Rathgeber, C.B.K.; Cabon, A.; Fonti, P. Turgor—A limiting factor for radial growth in mature conifers along an elevational gradient. New Phytol. 2021, 229, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Zindros, A.; Radoglou, K.; Milios, E.; Kitikidou, K. Tree line shift in the Olympus Mountain (Greece) and climate change. Forests 2020, 11, 985. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Trugman, A.T.; Badgley, G.; Anderson, C.M.; Bartuska, A.; Ciais, P.; Cullenward, D.; Field, C.B.; Freeman, J.; Goetz, S.J.; et al. Climate-driven risks to the climate mitigation potential of forests. Science 2020, 368, eaaz7005. [Google Scholar] [CrossRef]
- Kastridis, A.; Stathis, D.; Sapountzis, M.; Theodosiou, G. Insect outbreak and long-term post-fire effects on soil erosion in Mediterranean suburban Forest. Land 2022, 11, 911. [Google Scholar] [CrossRef]
- Stefanidis, S.; Alexandridis, V.; Mallinis, G. A cloud-based mapping approach for assessing spatiotemporal changes in erosion dynamics due to biotic and abiotic disturbances in a Mediterranean Peri-Urban Forest. Catena 2022, 218, 106564. [Google Scholar] [CrossRef]
- Huang, J.G.; Ma, Q.Q.; Rossi, S.; Biondi, F.; Deslauriers, A.; Fonti, P.; Liang, E.Y.; Makinen, H.; Oberhuber, W.; Rathgeber, C.B.K.; et al. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers. Proc. Natl. Acad. Sci. USA 2020, 117, 20645–20652. [Google Scholar] [CrossRef]
- Lu, X.M.; Huang, R.; Wang, Y.F.; Sigdel, S.R.; Dawadi, B.; Liang, E.Y.; Camarero, J.J. Summer temperature drives radial growth of alpine shrub willows on the northeastern Tibetan Plateau. Arct. Antarct. Alp. Res. 2016, 48, 461–468. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Cufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gricar, J.; Gruber, A.; Huang, J.G.; et al. Pattern of xylem phenology in conifers of cold ecosystems at the Northern Hemisphere. Glob. Change Biol. 2016, 22, 3804–3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tumajer, J.; Shishov, V.V.; Ilyin, V.A.; Camarero, J.J. Intra-annual growth dynamics of Mediterranean pines and junipers determines their climatic adaptability. Agric. For. Meteorol. 2021, 311, 108685. [Google Scholar] [CrossRef]
- Delpierre, N.; Lireux, S.; Hartig, F.; Camarero, J.J.; Cheaib, A.; Cufar, K.; Cuny, H.; Deslauriers, A.; Fonti, P.; Gricar, J.; et al. Chilling and forcing temperatures interact to predict the onset of wood formation in Northern Hemisphere conifers. Glob. Change Biol. 2019, 25, 1089–1105. [Google Scholar] [CrossRef] [PubMed]
- Popkova, M.I.; Vaganov, E.A.; Shishov, V.V.; Babushkina, E.A.; Rossi, S.; Fonti, M.V.; Fonti, P. Modeled tracheidograms disclose drought influence on Pinus sylvestris tree-rings structure from Siberian forest-steppe. Front. Plant Sci. 2018, 9, 1144. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Rossi, S.; Deslauriers, A.; Liu, J.Q. Contrasting strategies of xylem formation between black spruce and balsam fir in Quebec, Canada. Tree Physiol. 2019, 39, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Rossi, S.; Gricar, J.; Liang, E.Y.; Cufar, K. Is precipitation a trigger for the onset of xylogenesis in Juniperus przewalskii on the north-eastern Tibetan Plateau? Ann. Bot. 2015, 115, 629–639. [Google Scholar] [CrossRef]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Braker, O.U.; Schoch, W.; Innes, J.L. Identification, measurement and interpretation of tree rings in woody species from Mediterranean climates. Biol. Rev. Camb. Philos. Soc. 2003, 78, 119–148. [Google Scholar] [CrossRef]
- Kastridis, A.; Kamperidou, V.; Stathis, D. Dendroclimatological analysis of Fir (A. borisii-regis) in Greece in the frame of climate change Investigation. Forests 2022, 13, 879. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef]
- Stefanidis, S.; Alexandridis, V. Precipitation and potential evapotranspiration temporal variability and their relationship in two forest ecosystems in Greece. Hydrology 2021, 8, 160. [Google Scholar] [CrossRef]
- Cuny, H.E.; Fonti, P.; Rathgeber, C.B.K.; von Arx, G.; Peters, R.L.; Frank, D.C. Couplings in cell differentiation kinetics mitigate air temperature influence on conifer wood anatomy. Plant Cell Environ. 2019, 42, 1222–1232. [Google Scholar] [CrossRef]
- Marchand, L.J.; Dox, I.; Gricar, J.; Prislan, P.; Van den Bulcke, J.; Fonti, P.; Campioli, M. Timing of spring xylogenesis in temperate deciduous tree species relates to tree growth characteristics and previous autumn phenology. Tree Physiol. 2021, 41, 1161–1170. [Google Scholar] [CrossRef]
- Sprengel, L.; Stangler, D.F.; Sheppard, J.; Morhart, C.; Spiecker, H. Comparative analysis of the effects of stem height and artificial pruning on seasonal radial growth dynamics of Wild Cherry (Prunus avium L.) and Sycamore (Acer pseudoplatanus L.) in a widely spaced system. Forests 2018, 9, 174. [Google Scholar] [CrossRef]
- Guada, G.; Vazquez-Ruiz, R.A.; Garcia-Gonzalez, I. Meteorological conditions control the cessation rather than the beginning of wood formation in a sub-Mediterranean ring-porous oak. Agric. For. Meteorol. 2020, 281, 107833. [Google Scholar] [CrossRef]
- Michelot, A.; Simard, S.; Rathgeber, C.; Dufrene, E.; Damesin, C. Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol. 2012, 32, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-de-Una, L.; Aranda, I.; Rossi, S.; Fonti, P.; Canellas, I.; Gea-Izquierdo, G. Divergent phenological and leaf gas exchange strategies of two competing tree species drive contrasting responses to drought at their altitudinal boundary. Tree Physiol. 2018, 38, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Liang, E.; Gricar, J.; Prislan, P.; Rossi, S.; Cufar, K. Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau. Tree Physiol. 2013, 33, 48–56. [Google Scholar] [CrossRef]
- Li, X.X.; Rossi, S.; Liang, E.Y.; Camarero, J.J. Temperature thresholds for the onset of xylogenesis in alpine shrubs on the Tibetan Plateau. Trees 2016, 30, 2091–2099. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Gou, X.H.; Manzanedo, R.D.; Zhang, F.; Pederson, N. Cambial phenology and xylogenesis of Juniperus przewalskii over a climatic gradient is influenced by both temperature and drought. Agric. For. Meteorol. 2018, 260, 165–175. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Gou, X.H.; Pederson, N.; Zhang, F.; Niu, H.G.; Zhao, S.D.; Wang, F. Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region. Tree Physiol. 2018, 38, 840–852. [Google Scholar] [CrossRef]
- Huang, J.G.; Guo, X.; Rossi, S.; Zhai, L.; Yu, B.; Zhang, S.; Zhang, M. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season. Tree Physiol. 2018, 38, 1225–1236. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Gou, X.; Xia, J.; Wang, F.; Fang, F.; Zhang, J. Research progress on cambial activity of trees and the influencing factors. Chin. J. Appl. Ecol. 2021, 32, 3761–3770. [Google Scholar] [CrossRef]
- Rao, G.D.; Sui, J.K.; Zeng, Y.F.; He, C.Y.; Duan, A.G.; Zhang, J.G. De novo transcriptome and small RNA analysis of two Chinese willow cultivars reveals stress response genes in Salix matsudana. PLoS ONE 2014, 9, 1932–6203. [Google Scholar] [CrossRef]
- Tao, Z.X.; Xu, Y.J.; Ge, Q.S.; Dai, J.H.; Wang, H.J. Reduced frost hardiness in temperate woody species due to climate warming: A model-based analysis. Clim. Change 2021, 165, 35. [Google Scholar] [CrossRef]
- Feng, X.Y. A Comparative Study of Plant Hydraulic and Gas Exchange Traits in Salix Species. Master’s Thesis, Lanzhou University, Lanzhou, China, 2020. [Google Scholar] [CrossRef]
- Wu, Z.Y. Flora of China; Science Press: Beijing, China, 1999; pp. 162–274. [Google Scholar]
- Bhatta, S.P.; Sharma, K.P.; Balami, S. Variation in carbon storage among tree species in the planted forest of Kathmandu, Central Nepal. Curr. Sci. 2018, 115, 274–282. [Google Scholar] [CrossRef]
- Ogasa, M.; Miki, N.H.; Murakami, Y.; Yoshikawa, K. Recovery performance in xylem hydraulic conductivity is correlated with cavitation resistance for temperate deciduous tree species. Tree Physiol. 2013, 33, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Fan, J.; Ge, J.M.; Wang, Q.M.; Fu, W. Discrepancy in tree transpiration of Salix matsudana, Populus simonii under distinct soil, topography conditions in an ecological rehabilitation area on the northern Loess Plateau. For. Ecol. Manag. 2019, 432, 675–685. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, Y.; Chen, Y.; Feng, W.; Zhang, J. Uncovering candidate genes responsive to salt stress in Salix matsudana (Koidz) by transcriptomic analysis. PLoS ONE. 2020, 15, e0236129. [Google Scholar] [CrossRef]
- Zhang, J.; Yuan, H.W.; Yang, Q.S.; Li, M.; Wang, Y.; Li, Y.J.; Ma, X.J.; Tan, F.; Wu, R.L. The genetic architecture of growth traits in Salix matsudana under salt stress. Hortic. Res. 2017, 4, 17024. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Xu, J.L.; Jiang, Y.; Mandra, T.; Rademacher, T.T.; Xue, F.; Dong, M.Y.; Pederson, N. Higher plasticity of water uptake in spruce than larch in an alpine habitat of North-Central China. Agric. For. Meteorol. 2021, 311, 108696. [Google Scholar] [CrossRef]
- Rossi, S.; Menardi, R.; Anfodillo, T. Trephor: A new tool for sampling microcores from tree stems. IAWA J. 2006, 27, 89–97. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Xu, J.L.; Su, W.; Zhao, X.P.; Xu, X.L. Spring precipitation effects on formation of first row of earlywood vessels in Quercus variabilis at Qinling Mountain (China). Trees 2019, 33, 457–468. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Jiang, Y.; Wen, Y.; Ding, X.Y.; Wang, B.; Xu, J.L. Comparing primary and secondary growth of co-occurring deciduous and evergreen conifers in an alpine habitat. Forests 2019, 10, 574. [Google Scholar] [CrossRef]
- Zhang, J.Z. Cambial Phenology and Intra-Annual Radial Growth Dynamics of Conifers over the Qilian Mountains. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2018. [Google Scholar] [CrossRef]
- Vavrčík, H.; Gryc, V.; Vichrová, G. Xylem formation in young Norway spruce trees in drahany highland, Czech Republic. IAWA J. 2013, 34, 231–244. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T. Assessment of cambial activity and xylogenesis by microsampling tree species: An example at the alpine timberline. IAWA J. 2006, 27, 383–394. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Begin, Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Can. J. For. Res. 2003, 33, 190. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, Y.P.; Guo, Y.Y.; Kang, M.Y.; Wang, M.C.; Wang, B. Intra-annual xylem growth of Larix principis-rupprechtii at its upper and lower distribution limits on the Luyashan mountain in North-Central China. Forests 2015, 6, 3809–3827. [Google Scholar] [CrossRef]
- Rathgeber, C.B.K.; Rossi, S.; Bontemps, J.D. Cambial activity related to tree size in a mature silver-fir plantation. Ann. Bot. 2011, 108, 429–438. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Bauman, D.; Fortunel, C.; Delhaye, G.; Malhi, Y.; Cernusak, L.A.; Bentley, L.P.; Rifai, S.W.; Aguirre-Gutiérrez, J.; Menor, I.O.; Phillips, O.L.; et al. Tropical tree mortality has increased with rising atmospheric water stress. Nature 2022, 608, 528–533. [Google Scholar] [CrossRef]
- Zhang, S.K.; Huang, J.G.; Rossi, S.; Ma, Q.Q.; Yu, B.Y.; Zhai, L.H.; Luo, D.W.; Guo, X.L.; Fu, S.L.; Zhang, W. Intra-annual dynamics of xylem growth in Pinus massoniana submitted to an experimental nitrogen addition in Central China. Tree Physiol. 2017, 37, 1546–1553. [Google Scholar] [CrossRef]
- Wang, Q.B.; Qiao, H.B.; Zhang, D. Expected maturity age of individual Salix matsudana. Prot. For. Sci. Technol. 2014, 12, 24–25. [Google Scholar] [CrossRef]
- Ma, X.D.; Pang, Z.; Wu, J.Y.; Zhang, G.F.; Dai, Y.C.A.; Zou, J.L.; Kan, H.M. Seasonal pattern of stem radial growth of Salix matsudana and its response to climatic and soil factors in a semi-arid area of North China. Glob. Ecol. Conserv. 2021, 28, e01701. [Google Scholar] [CrossRef]
- Huang, Y.Q.; Deng, X.W.; Zhao, Z.H.; Xiang, W.H.; Yan, W.D.; Ouyang, S.; Lei, P.F. Monthly radial growth model of Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.), and the relationships between radial increment and climate factors. Forests 2019, 10, 757. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Danpradit, S.; Tadang, N.; Buajan, S.; Muangsong, C. Cambial activity and radial growth dynamics of three tropical tree species at Chang Island, Thailand. J. Trop. For. Sci. 2019, 31, 404–414. [Google Scholar] [CrossRef]
- Liu, T.D.; Jiang, K.; Tan, Z.Y.; He, Q.F.; Zhang, H.; Wang, C. A method for performing reforestation to effectively recover soil water content in extremely degraded tropical rain forests. Front. Ecol. Evol. 2021, 9, 643994. [Google Scholar] [CrossRef]
- Rossi, S.; Girard, M.J.; Morin, H. Lengthening of the duration of xylogenesis engenders disproportionate increases in xylem production. Glob. Change Biol. 2014, 20, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Saderi, S.; Rathgeber, C.B.K.; Rozenberg, P.; Fournier, M. Phenology of wood formation in larch (Larix decidua Mill.) trees growing along a 1000 m elevation gradient in the French Southern Alps. Ann. For. Sci. 2019, 76, 89. [Google Scholar] [CrossRef]
- He, M.; Yang, B.; Wang, Z.; Bruning, A.; Pourtahmasi, K.; Oladi, R. Climatic forcing of xylem formation in Qilian juniper on the northeastern Tibetan Plateau. Trees 2016, 30, 923–933. [Google Scholar] [CrossRef]
- Vieira, J.; Carvalho, A.; Campelo, F. Tree growth under climate change: Evidence from xylogenesis timings and kinetics. Front. Plant Sci. 2020, 11, 90. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Enlarging Cell | First Mature Cell | End of Cambial Activity | Completion Lignification | |
|---|---|---|---|---|
| Salix babylonica | 85 ± 9 | 111 ± 6 | 290 ± 14 | 321 ± 5 |
| Salix matsudana | 78 ± 4 | 113 ± 4 | 285 ± 0 | 311 ± 4 |
| z | −0.943 | −0.471 | −0.696 | −1.65 |
| p | 0.346 | 0.653 | 0.487 | 0.099 |
| Measured Value (μm) | A (μm) | rmean (μm/d) | rmax (μm/d) | xc (DOY) | ||
|---|---|---|---|---|---|---|
| Salix babylonica | 1 | 7870 | 7154 | 45.59 | 74.54 | 150 |
| 2 | 9106 | 47.48 | 77.64 | 161 | ||
| 3 | 7634 | 8342 | 53.25 | 87.07 | 153 | |
| Salix matsudana | 1 | 7742 | 8636 | 34.53 | 56.45 | 153 |
| 2 | 6778 | 7560 | 37.98 | 62.10 | 145 | |
| 3 | 5329 | 6614 | 49.05 | 80.20 | 140 |
| Model Parameters | Xylem Growth Increment | Xylem Growth Rate | ||||
|---|---|---|---|---|---|---|
| Estimate | SE | t-Value | Estimate | SE | t-Value | |
| Air temperature | ||||||
| Intercept | −153.31 | 40.04 | −3.83 *** | −12.15 | 5.70 | −2.13 * |
| Ta | 21.65 | 2.02 | 10.74 *** | 2.22 | 0.29 | 7.75 *** |
| S. matsudana | −119.31 | 56.62 | −2.11 * | −12.46 | 8.06 | −1.55 |
| Ta × S. matsudana | 8.13 | 2.85 | 2.85 ** | 0.82 | 0.41 | 2.03 * |
| Ground surface temperature | ||||||
| Intercept | −124.81 | 41.12 | −3.04 ** | −10.68 | 5.52 | −1.94 |
| GST | 17.87 | 1.83 | 9.77 *** | 1.91 | 0.25 | 7.77 *** |
| S. matsudana | −105.14 | 58.16 | −1.81 | −11.32 | 7.80 | −1.45 |
| GST × S. matsudana | 6.55 | 2.59 | 2.53 * | 0.68 | 0.35 | 1.95 |
| Precipitation | ||||||
| Intercept | 158.99 | 28.14 | 5.65 *** | 23.42 | 3.70 | 6.32 *** |
| PRE | 3.41 | 0.73 | 4.66 *** | 0.22 | 0.10 | 2.26 * |
| S. matsudana | −6.04 | 39.80 | −0.15 | −0.33 | 5.24 | −0.06 |
| PRE × S. matsudana | 1.44 | 1.04 | 1.39 | 0.12 | 0.14 | 0.87 |
| Relative humidity | ||||||
| Intercept | 4.33 | 102.01 | 0.04 | 23.40 | 12.60 | 1.86 |
| RH | 398.63 | 161.96 | 2.46 * | 9.40 | 20.00 | 0.47 |
| S. matsudana | −77.97 | 144.29 | −0.54 | −4.44 | 17.82 | −0.25 |
| RH × S. matsudana | 178.89 | 229.05 | 0.78 | 11.81 | 28.29 | 0.42 |
| Vapor pressure deficit | ||||||
| Intercept | 110.24 | 50.28 | 2.19 * | 5.84 | 5.48 | 1.06 |
| VPD | 151.74 | 62.57 | 3.08 ** | 25.51 | 5.38 | 4.74 *** |
| S. matsudana | −17.66 | 71.11 | −0.25 | −4.67 | 7.76 | −0.60 |
| VPD × S. matsudana | 54.12 | 69.67 | 0.78 | 8.18 | 7.60 | 1.08 |
| Sunshine duration | ||||||
| Intercept | 132.77 | 65.75 | 2.02 * | 7.34 | 7.51 | 0.98 |
| SSD | 17.90 | 9.46 | 1.89 | 3.36 | 1.08 | 3.11 ** |
| S. matsudana | 14.12 | 92.98 | 0.15 | 0.91 | 10.61 | 0.09 |
| SSD × S. matsudana | 2.73 | 13.38 | 0.20 | 0.29 | 1.53 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Luo, P.; Xu, J.; Hou, J.; Zhai, L. Intra-Annual Growth and Its Response to Climatic Factors of Two Salix Species under Warm Temperate Environment. Forests 2022, 13, 1441. https://doi.org/10.3390/f13091441
Zhang Y, Luo P, Xu J, Hou J, Zhai L. Intra-Annual Growth and Its Response to Climatic Factors of Two Salix Species under Warm Temperate Environment. Forests. 2022; 13(9):1441. https://doi.org/10.3390/f13091441
Chicago/Turabian StyleZhang, Yiping, Pengfei Luo, Junliang Xu, Jiayu Hou, and Lexin Zhai. 2022. "Intra-Annual Growth and Its Response to Climatic Factors of Two Salix Species under Warm Temperate Environment" Forests 13, no. 9: 1441. https://doi.org/10.3390/f13091441