

Isolated Trees in Two Tibetan Plateau Treelines Reveal Growth Plasticity to Harsh Conditions of the Little Ice Age

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
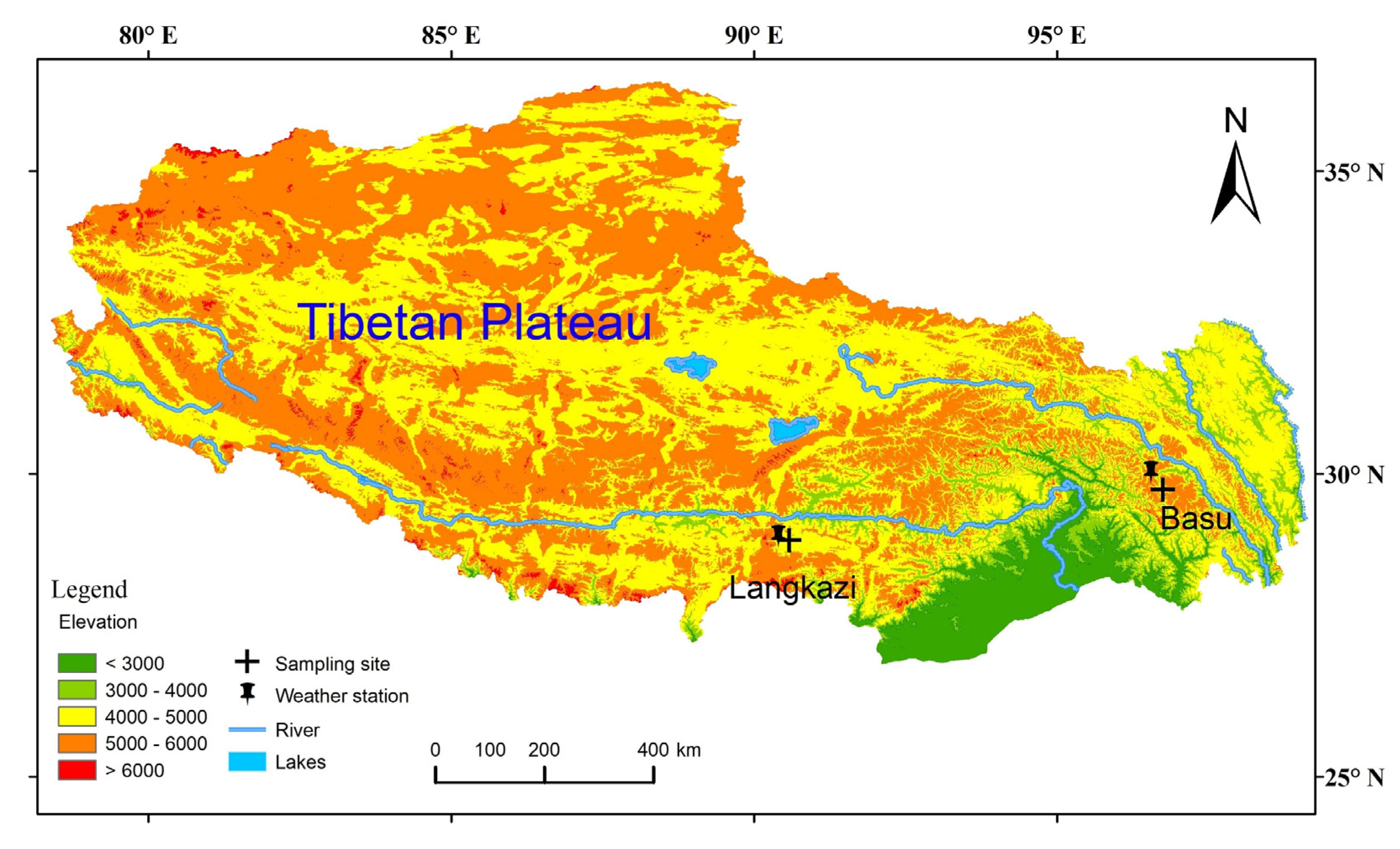
2.1. Study Sites and Tree-Ring Data
2.2. Comparison of Radial Growth Rates during the Little Ice Age and after
2.3. Comparison of Growth Synchronicity among Trees over Time
3. Results
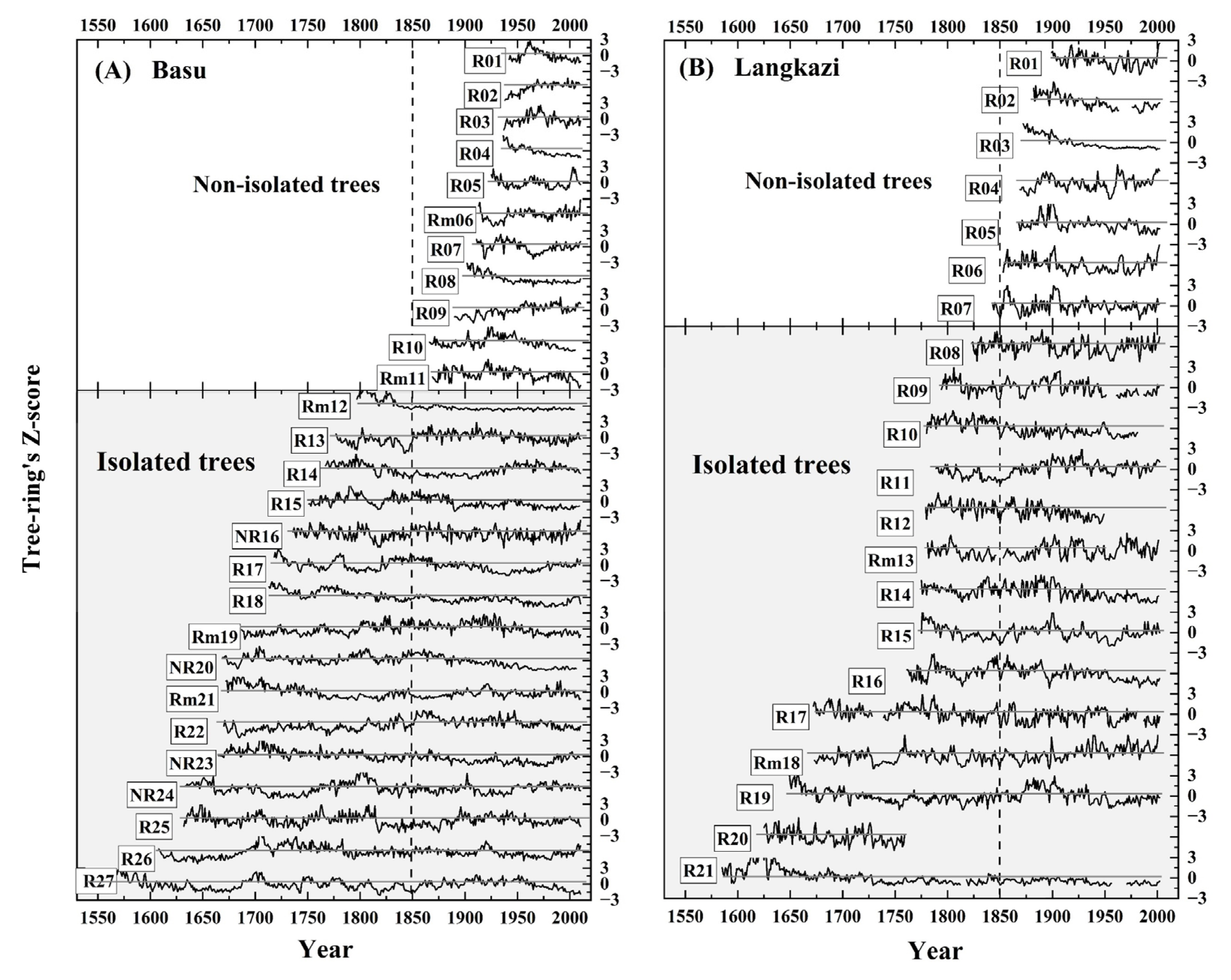
3.1. Age of the Tree-Ring Samples
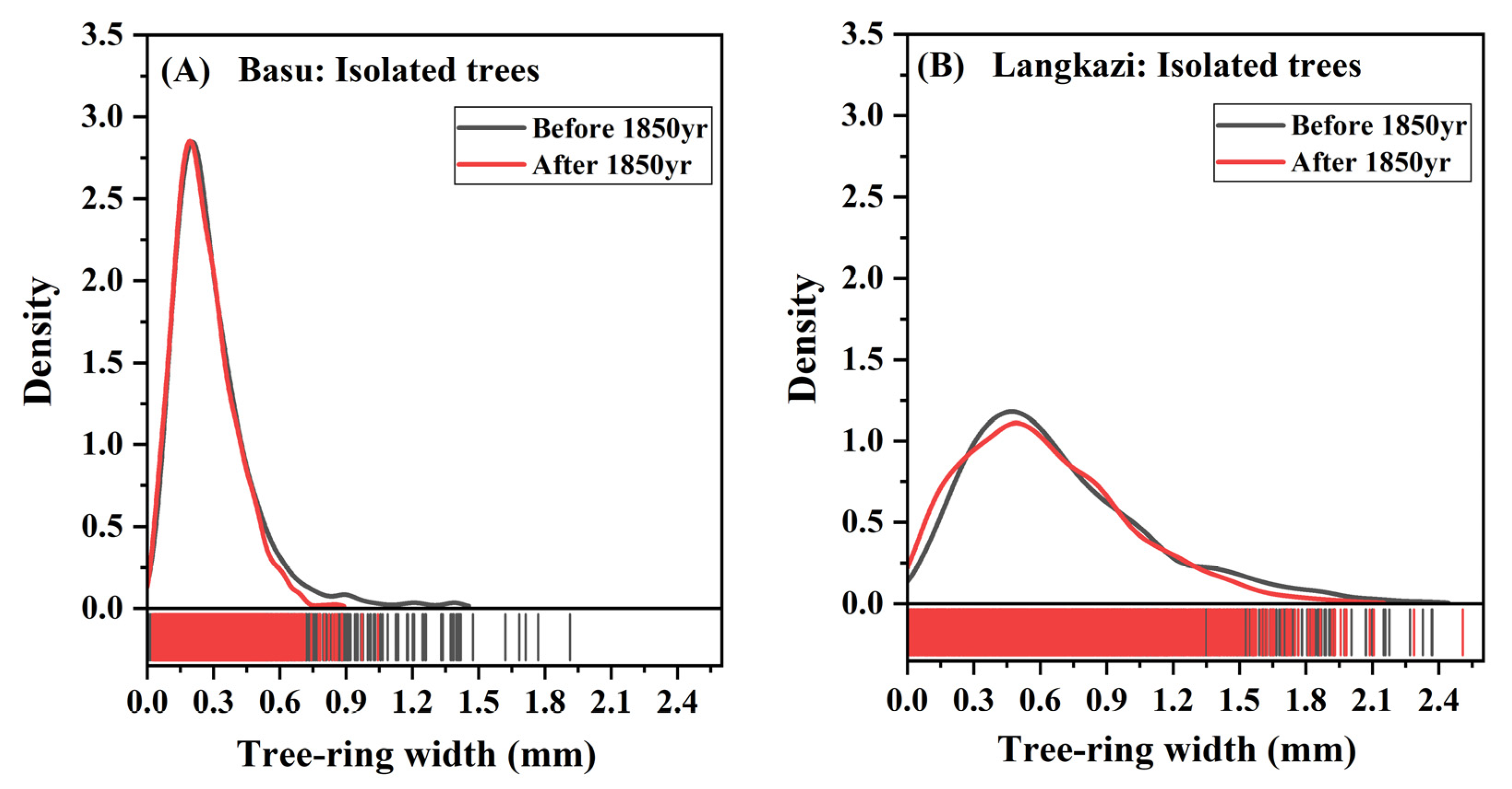
3.2. Radial Growth Rates during the Little Ice Age and after
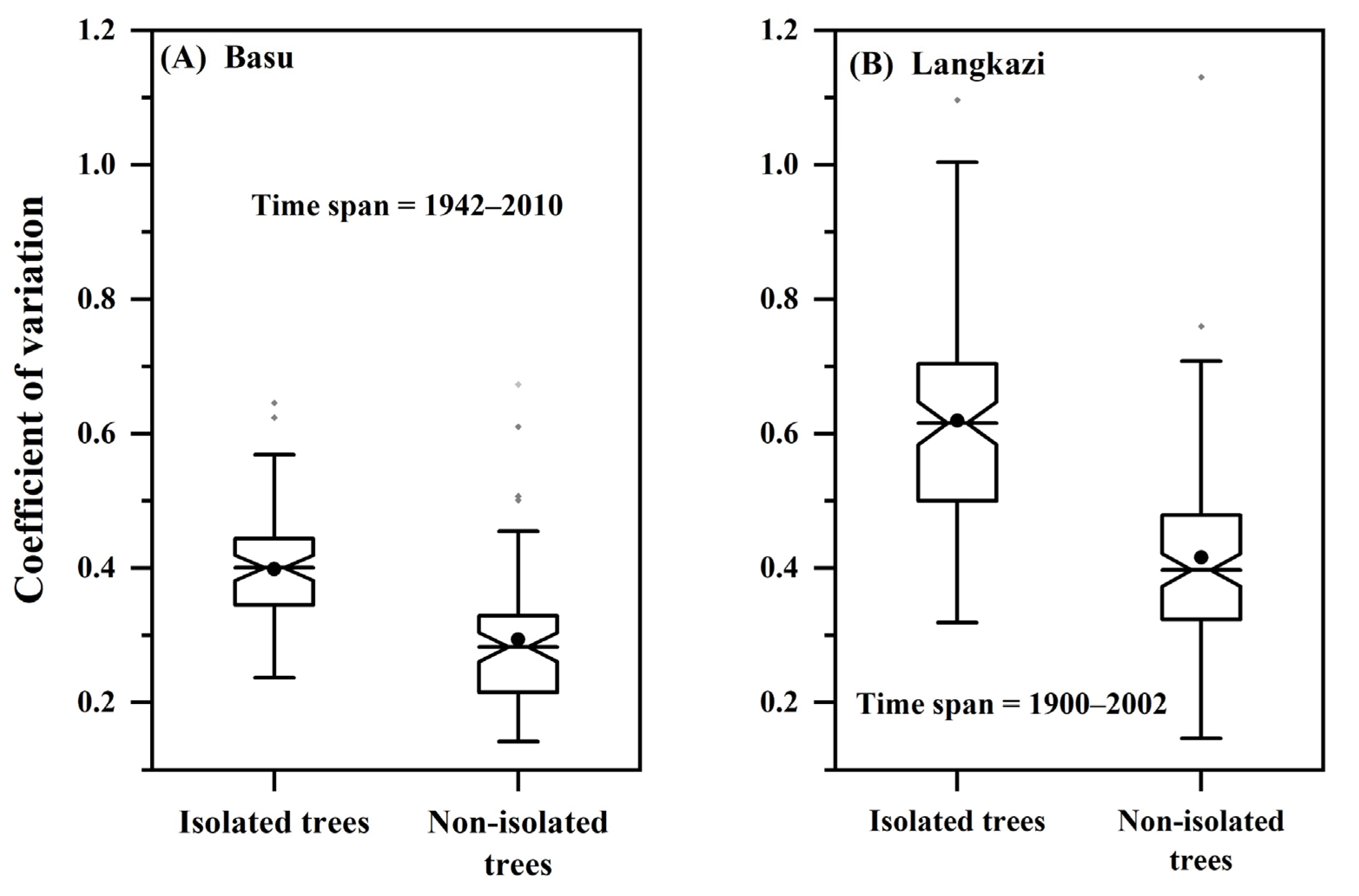
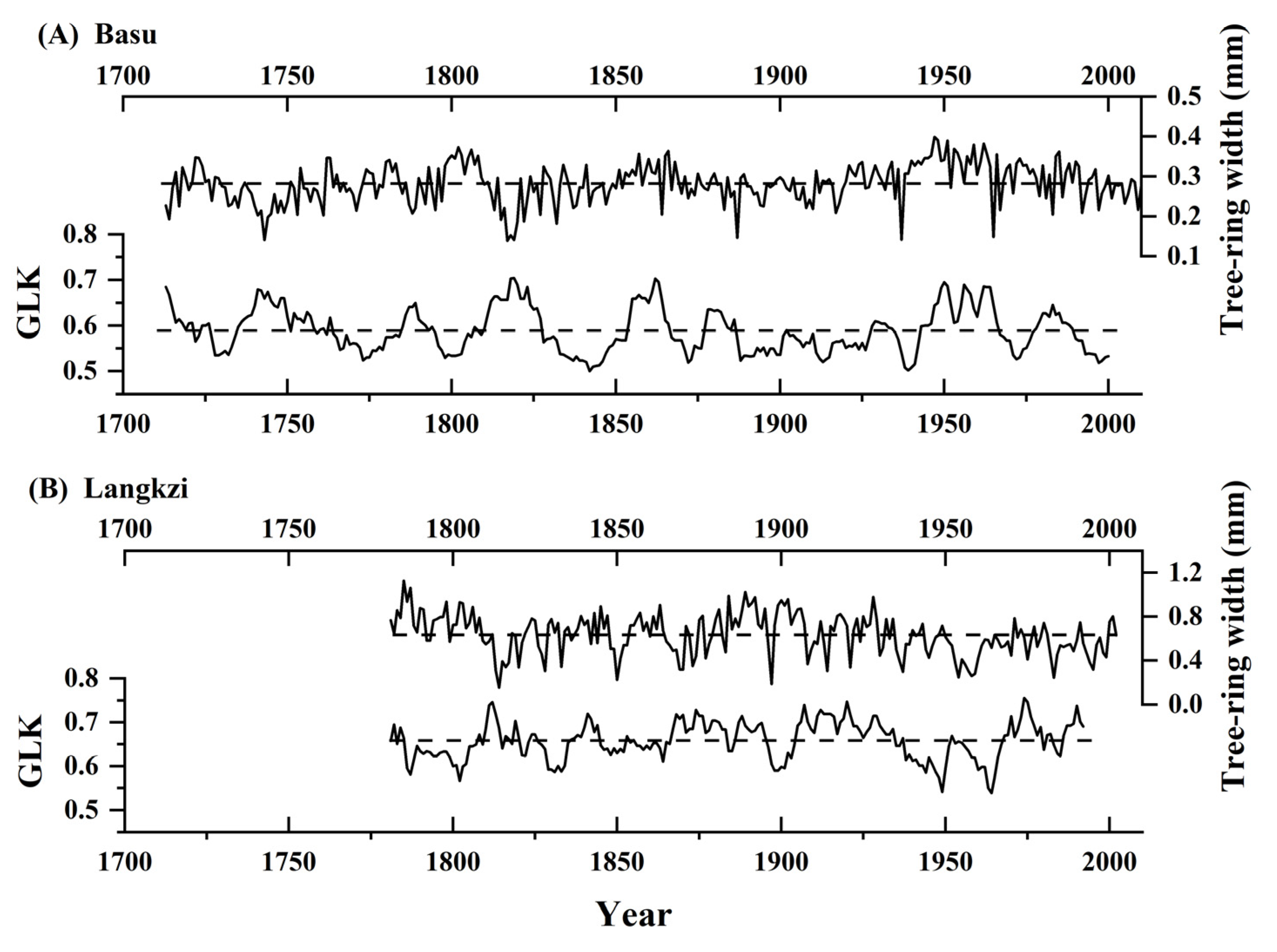
3.3. Growth Synchronicity among Trees over Time
4. Discussion
4.1. Absence of Age-Related Growth Trends in Treeline Trees
4.2. Comparison of Tree-Ring Widths in Isolated and Non-Isolated Trees in Their Common Growth Period
4.3. Fluctuations in the Growth Synchronicity among Trees
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Körner, C. A re-assessment of high elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [CrossRef]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Lyu, L.; Zhang, Q.-B.; Pellatt, M.G.; Büntgen, U.; Li, M.-H.; Cherubini, P. Drought limitation on tree growth at the Northern Hemisphere’s highest tree line. Dendrochronologia 2019, 53, 40–47. [Google Scholar]
- Kullman, L. Pine (Pinus sylvestris) treeline dynamics during the past millennium—A population study in west-central Sweden. Ann. Bot. Fenn. 2005, 42, 95–106. [Google Scholar]
- Salzer, M.W.; Hughes, M.K.; Bunn, A.G.; Kipfmueller, K.F. Recent unprecedented tree-ring growth in bristlecone pine at the highest elevations and possible causes. Proc. Natl. Acad. Sci. USA 2009, 106, 20348–20353. [Google Scholar] [PubMed]
- Zhu, H.-F.; Shao, X.-M.; Yin, Z.-Y.; Xu, P.; Xu, Y.; Tian, H. August temperature variability in the southeastern Tibetan Plateau since AD 1385 inferred from tree rings. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 305, 84–92. [Google Scholar] [CrossRef]
- Bunn, A.G.; Salzer, M.W.; Anchukaitis, K.J.; Bruening, J.M.; Hughes, M.K. Spatiotemporal variability in the climate growth response of high elevation bristlecone pine in the White Mountains of California. Geophys. Res. Lett. 2018, 45, 13–312. [Google Scholar] [CrossRef]
- Vallée, S.; Payette, S. Contrasted growth of black spruce (Picea mariana) forest trees at treeline associated with climate change over the last 400 years. Arct. Antarct. Alp. Res. 2004, 36, 400–406. [Google Scholar] [CrossRef]
- Kullman, L. Short-term population trends of isolated tree-limit stands of Pinus sylvestris L. in central Sweden. Arct. Alp. Res. 1983, 15, 369–382. [Google Scholar] [CrossRef]
- Renard, S.M.; McIntire, E.J.B.; Fajardo, A. Winter conditions–not summer temperature–influence establishment of seedlings at white spruce alpine treeline in Eastern Quebec. J. Veg. Sci. 2016, 27, 29–39. [Google Scholar]
- Smith, W.K.; Germino, M.J.; Hancock, T.E.; Johnson, D.M. Another perspective on altitudinal limits of alpine timberlines. Tree Physiol. 2003, 23, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Maher, C.T.; Nelson, C.R.; Larson, A.J. Winter damage is more important than summer temperature for maintaining the krummholz growth form above alpine treeline. J. Ecol. 2020, 108, 1074–1087. [Google Scholar] [CrossRef]
- Kullman, L. Cold-induced dieback of montane spruce forests in the Swedish Scandes–a modern analogue of paleoenvironmental processes. New Phytol. 1989, 113, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Pereg, D.; Payette, S. Development of black spruce growth forms at treeline. Plant Ecol. 1998, 138, 137–147. [Google Scholar] [CrossRef]
- Daly, C.; Shankman, D. Seedling establishment by conifers above tree limit on Niwot Ridge, Front Range, Colorado, USA. Arct. Alp. Res. 1985, 17, 389–400. [Google Scholar] [CrossRef]
- Piovesan, G.; Biondi, F. On tree longevity. New Phytol. 2021, 231, 1318–1337. [Google Scholar]
- Sánchez-Salguero, R.; Camarero, J.J.; Rozas, V.; Génova, M.; Olano, J.M.; Arzac, A.; Gazol, A.; Caminero, L.; Tejedor, E.; de Luis, M.; et al. Resist, recover or both? Growth plasticity in response to drought is geographically structured and linked to intraspecific variability in Pinus pinaster. J. Biogeogr. 2018, 45, 1126–1139. [Google Scholar]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar]
- Wang, X.Y.; Wang, T.; Xu, J.F.; Shen, Z.H.; Yang, Y.P.; Chen, A.P.; Wang, S.P.; Liang, E.Y.; Piao, S.L. Enhanced habitat loss of the Himalayan endemic flora driven by warming-forced upslope tree expansion. Nat. Ecol. Evol. 2022, 6, 890–899. [Google Scholar] [CrossRef]
- Lu, X.M.; Liang, E.Y.; Babst, F.; Camarero, J.J.; Buntgen, U. Warming-induced tipping points of Arctic and alpine shrub recruitment. Proc. Natl. Acad. Sci. USA. 2022, 119, e2118120119. [Google Scholar] [CrossRef]
- Chen, D.L.; Xu, B.Q.; Yao, T.D.; Guo, Z.T.; Cui, P.; Chen, F.H.; Zhang, R.H.; Zhang, X.Z.; Zhang, Y.L.; Fan, J.; et al. Assessment of past, present and future environmental changes on the Tibetan Plateau. China Sci. Bull. 2015, 60, 3025–3035. (In Chinese) [Google Scholar] [CrossRef]
- Yu, W.; Wei, F.; Ma, Y.; Liu, W.; Zhang, Y.; Luo, L.; Tian, L.; Xu, B.; Qu, D. Stable isotope variations in precipitation over Deqin on the southeastern margin of the Tibetan Plateau during different seasons related to various meteorological factors and moisture sources. Atmos. Res. 2016, 170, 123–130. [Google Scholar]
- Von Storch, H.; Zwiers, F.W. Statistical Analysis in Climate Research; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Free, M.; Robock, A. Global warming in the context of the Little Ice Age. J. Geophys. Res.-Biogeosciences 1999, 104, 19057–19070. [Google Scholar]
- Terrell, G.R. The maximal smoothing principle in density estimation. J. Am. Stat. Assoc. 1990, 85, 470–477. [Google Scholar] [CrossRef]
- Krzywinski, M.; Altman, N. Visualizing samples with box plots. Nat. Methods 2014, 11, 119–120. [Google Scholar] [CrossRef]
- Huber, B. Über die Sicherheit jahrringschronologischer Datierung. Holz Als Roh-Und Werkst. 1943, 6, 263–268. [Google Scholar] [CrossRef]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar]
- Cook, E.R.; Briffa, K.R.; Shiyatov, S.; Mazepa, V. Tree-ring Standardization and Growth-Trend Estimation. In Methods of Dendrochronology: Applications in the Environmental Sciences; Springer Science & Business Media: New York, Ny, USA; Boston, MA, USA, 1990; pp. 104–123. [Google Scholar]
- Biondi, F.; Qeadan, F. A theory-driven approach to tree-ring standardization: Defining the biological trend from expected basal area increment. Tree-Ring Res. 2008, 64, 81–96. [Google Scholar]
- Thompson, L.G.; Tandong, Y.; Davis, M.E.; Mosley-Thompson, E.; Mashiotta, T.A.; Lin, P.-N.; Mikhalenko, V.N.; Zagorodnov, V.S. Holocene climate variability archived in the Puruogangri ice cap on the central Tibetan Plateau. Ann. Glaciol. 2006, 43, 61–69. [Google Scholar]
- Piovesan, G.; Biondi, F.; Baliva, M.; Dinella, A.; Di Fiore, L.; Marchiano, V.; Saba, E.P.; De Vivo, G.; Schettino, A.; Di Filippo, A. Tree growth patterns associated with extreme longevity: Implications for the ecology and conservation of primeval trees in Mediterranean mountains. Anthropocene 2019, 26, 100199. [Google Scholar]
- Piovesan, G.; Biondi, F.; Baliva, M.; De Vivo, G.; Marchianò, V.; Schettino, A.; Di Filippo, A. Lessons from the wild. Ecology 2019, 100, 1–4. [Google Scholar]
- Black, B.A.; Colbert, J.J.; Pederson, N. Relationships between radial growth rates and lifespan within North American tree species. Ecoscience 2008, 15, 349–357. [Google Scholar]
- Lauder, J.D.; Moran, E.V.; Hart, S.C. Fight or flight? Potential tradeoffs between drought defense and reproduction in coni-fers. Tree Physiol. 2019, 39, 1071–1085. [Google Scholar] [PubMed]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef]
- Endara, M.J.; Coley, P.D. The resource availability hypothesis revisited: A meta-analysis. Funct. Ecol. 2011, 25, 389–398. [Google Scholar]
- Bigler, C.; Veblen, T.T. Increased early growth rates decrease longevities of conifers in subalpine forests. Oikos 2009, 118, 1130–1138. [Google Scholar]
- Schulman, E. Longevity under adversity in conifers. Science 1954, 119, 396–399. [Google Scholar]
- Di Filippo, A.; Biondi, F.; Maugeri, M.; Schirone, B.; Piovesan, G. Bioclimate and growth history affect beech lifespan in the I talian A lps and A pennines. Glob. Chang. Biol. 2012, 18, 960–972. [Google Scholar]
- Pu, X.; Wang, X.; Lyu, L. Recent Warming-Induced Tree Growth Enhancement at the Tibetan Treeline and the Link to Improved Water-Use Efficiency. Forests 2021, 12, 1702. [Google Scholar] [CrossRef]
- Gleason, S.M.; Westoby, M.; Jansen, S.; Choat, B.; Hacke, U.G.; Pratt, R.B.; Bhaskar, R.; Brodribb, T.J.; Bucci, S.J.; Cao, K.F. Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species. New Phytol. 2016, 209, 123–136. [Google Scholar]
- Roskilly, B.; Keeling, E.; Hood, S.; Giuggiola, A.; Sala, A. Conflicting functional effects of xylem pit structure relate to the growth-longevity trade-off in a conifer species. Proc. Natl. Acad. Sci. USA 2019, 116, 15282–15287. [Google Scholar] [CrossRef] [PubMed]
- Liang, E.; Dawadi, B.; Pederson, N.; Eckstein, D. Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature? Ecology 2014, 95, 2453–2465. [Google Scholar] [CrossRef]
- Rüger, N.; Condit, R.; Dent, D.H.; DeWalt, S.J.; Hubbell, S.P.; Lichstein, J.W.; Lopez, O.R.; Wirth, C.; Farrior, C.E. Demographic trade-offs predict tropical forest dynamics. Science 2020, 368, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Waring, R.H. Characteristics of trees predisposed to die. Bioscience 1987, 37, 569–574. [Google Scholar] [CrossRef]
- Sousa, W.P. The role of disturbance in natural communities. Annu. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Galiano, L.; Martínez-Vilalta, J.; Lloret, F. Carbon reserves and canopy defoliation determine the recovery of Scots pine 4 yr after a drought episode. New Phytol. 2011, 190, 750–759. [Google Scholar] [CrossRef]
- Fang, O.; Alfaro, R.I.; Zhang, Q.-B. Tree rings reveal a major episode of forest mortality in the late 18th century on the Tibetan Plateau. Glob. Planet. Chang. 2018, 163, 44–50. [Google Scholar] [CrossRef]
- Dobbertin, M. Tree growth as indicator of tree vitality and of tree reaction to environmental stress: A review. Eur. J. For. Res. 2005, 124, 319–333. [Google Scholar] [CrossRef]
- Fajardo, A.; Mcintire, F.J.B. Reversal of multicentury tree growth improvements and loss of synchrony at mountain tree lines point to changes in key drivers. J. Ecol. 2012, 100, 782–794. [Google Scholar] [CrossRef]
- Camarerro, J.J.; Garol, A.; Sánchez-Salguero, R.; Fajardo, A.; McIntire, E.J.; Gutiérrez, E.; Batllori, E.; Boudreau, S.; Carrer, M.; Diez, J.; et al. Global fading of the temperature–growth coupling at alpine and polar treelines. Glob. Chang. Biol. 2021, 27, 1879–1889. [Google Scholar] [CrossRef]
- Shi, C.M.; Massion-Delmotte, V.; Daux, V.; Li, Z.S.; Zhang, Q.-B. An unstable tree-growth response to climate in two 500 year chronologies, North Eastern Qinghai-Tibetan Plateau. Dendrochronologia 2010, 28, 225–237. [Google Scholar] [CrossRef]
- Liu, J.; Deng, X.; Zhang, Q.-B. Relationship of tree growth and climatic factors at treeline of Picea likiangensis var. balfouriana forest in Basu County, Xizang (In Chinese with English abstract). Chin. J. Plant Ecol. 2015, 39, 442–452. [Google Scholar]
- Lyu, L.; Zhang, Q.-B.; Deng, X.; Mäkinen, H. Fine-scale distribution of treeline trees and the nurse plant facilitation on the eastern Tibetan Plateau. Ecol. Indic. 2016, 66, 251–258. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, J.; Lyu, L.; Zhang, Q. Isolated Trees in Two Tibetan Plateau Treelines Reveal Growth Plasticity to Harsh Conditions of the Little Ice Age. Forests 2022, 13, 1371. https://doi.org/10.3390/f13091371
Zheng J, Lyu L, Zhang Q. Isolated Trees in Two Tibetan Plateau Treelines Reveal Growth Plasticity to Harsh Conditions of the Little Ice Age. Forests. 2022; 13(9):1371. https://doi.org/10.3390/f13091371
Chicago/Turabian StyleZheng, Jiacheng, Lixin Lyu, and Qibin Zhang. 2022. "Isolated Trees in Two Tibetan Plateau Treelines Reveal Growth Plasticity to Harsh Conditions of the Little Ice Age" Forests 13, no. 9: 1371. https://doi.org/10.3390/f13091371