
495-Year Wood Anatomical Record of Siberian Stone Pine (Pinus sibirica Du Tour) as Climatic Proxy on the Timberline

 , , and
, , and
Abstract
:1. Introduction
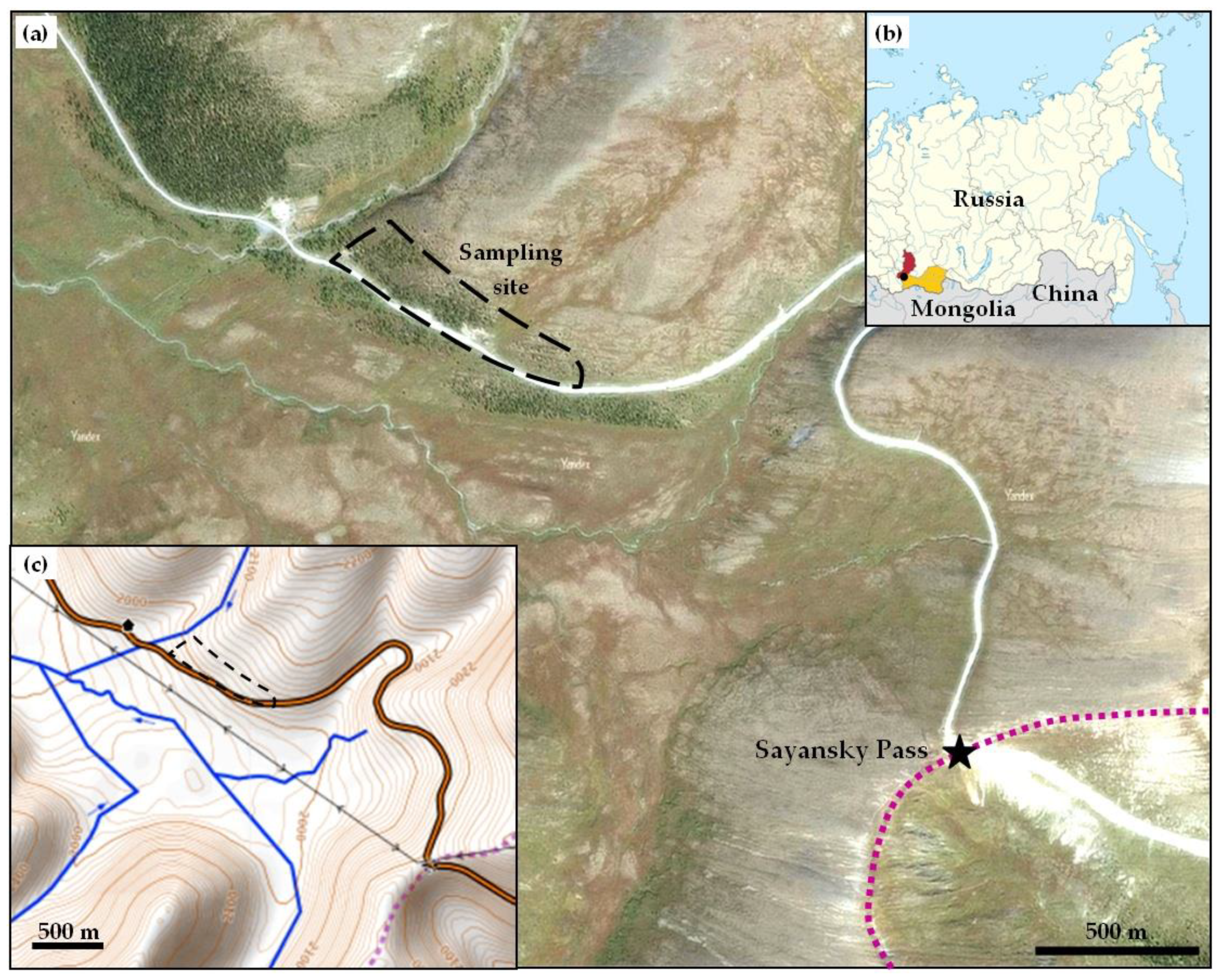
2. Materials and Methods
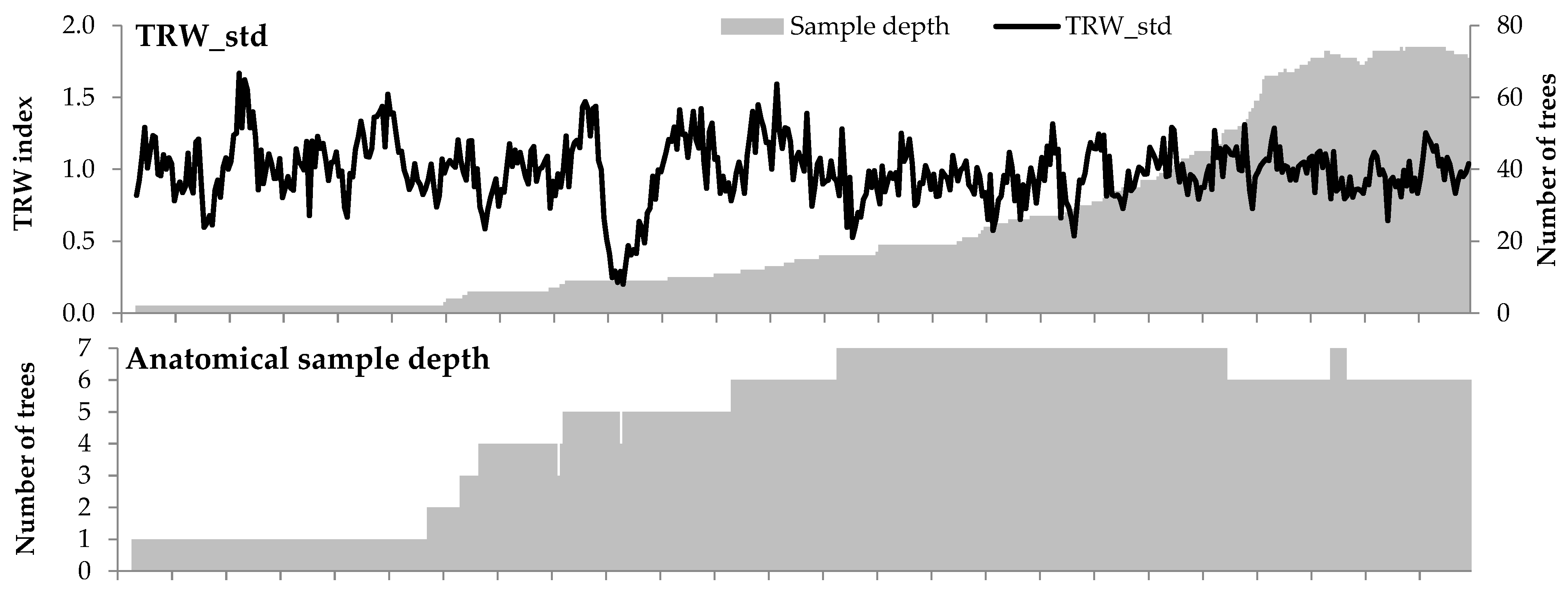
3. Results
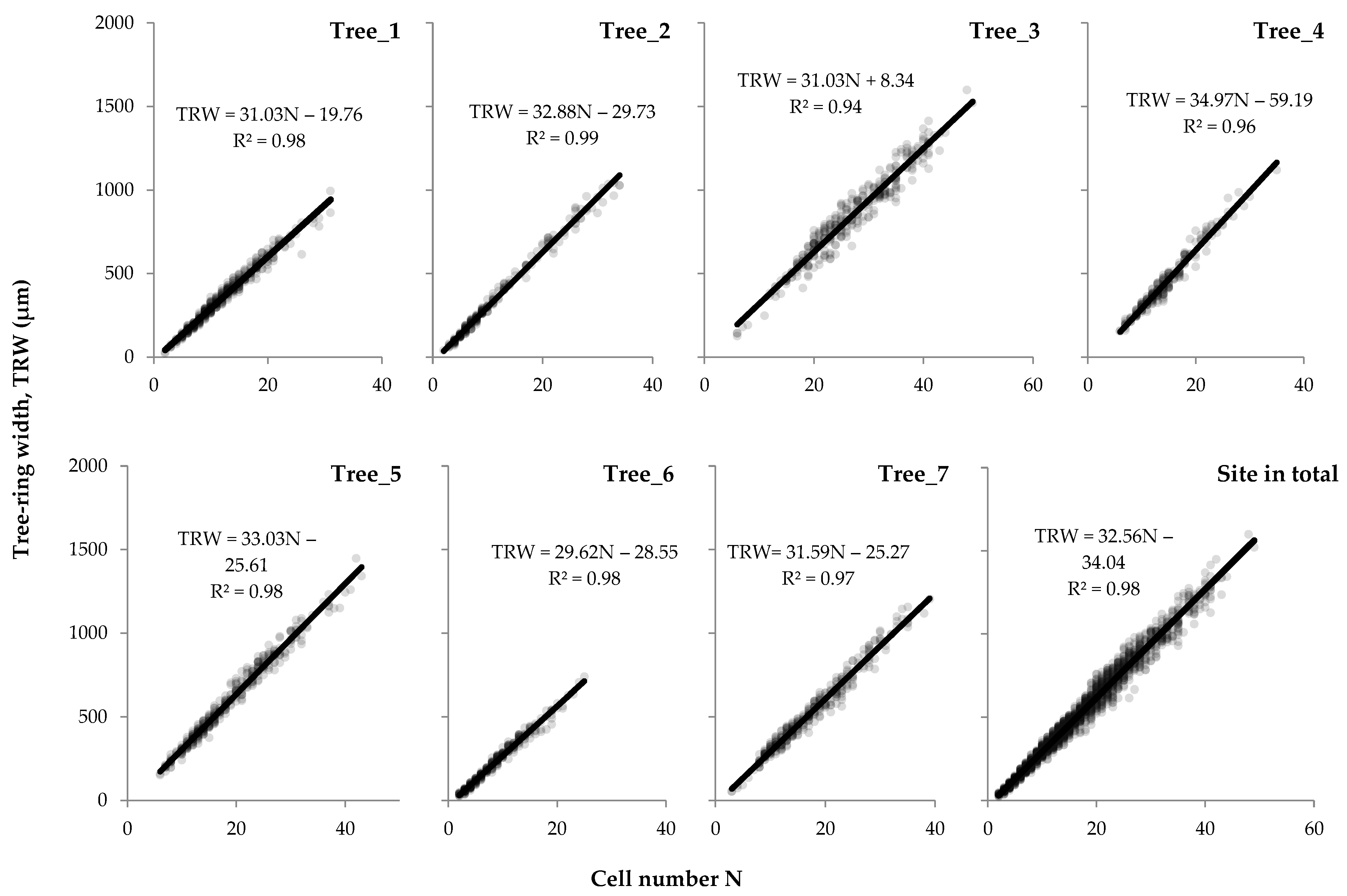
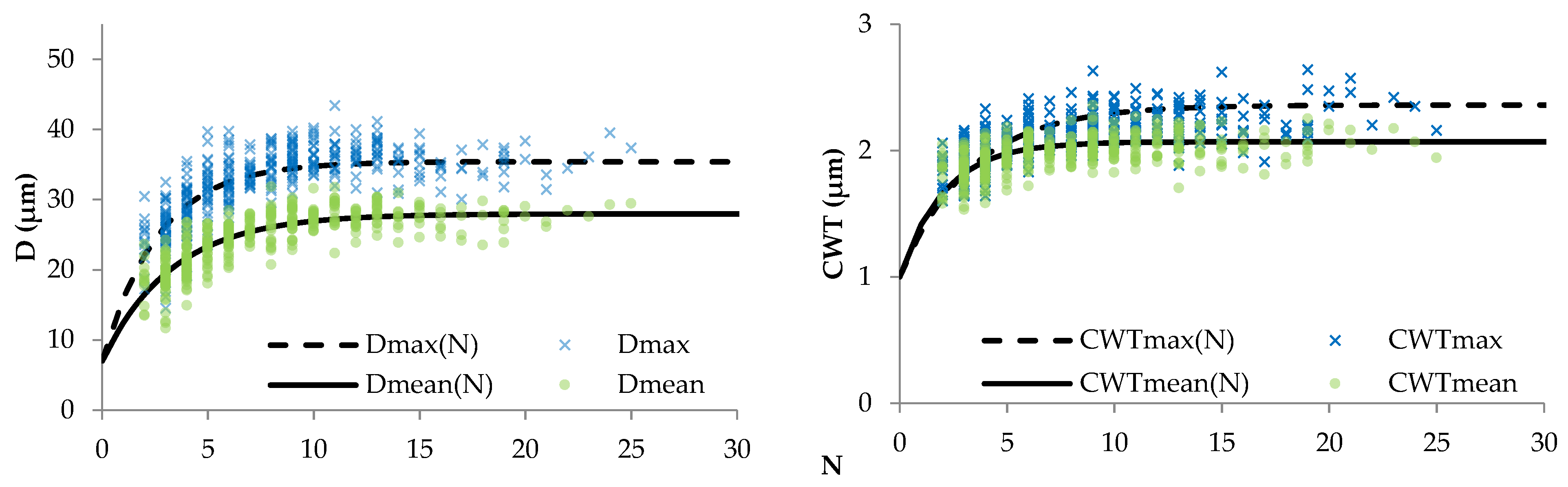
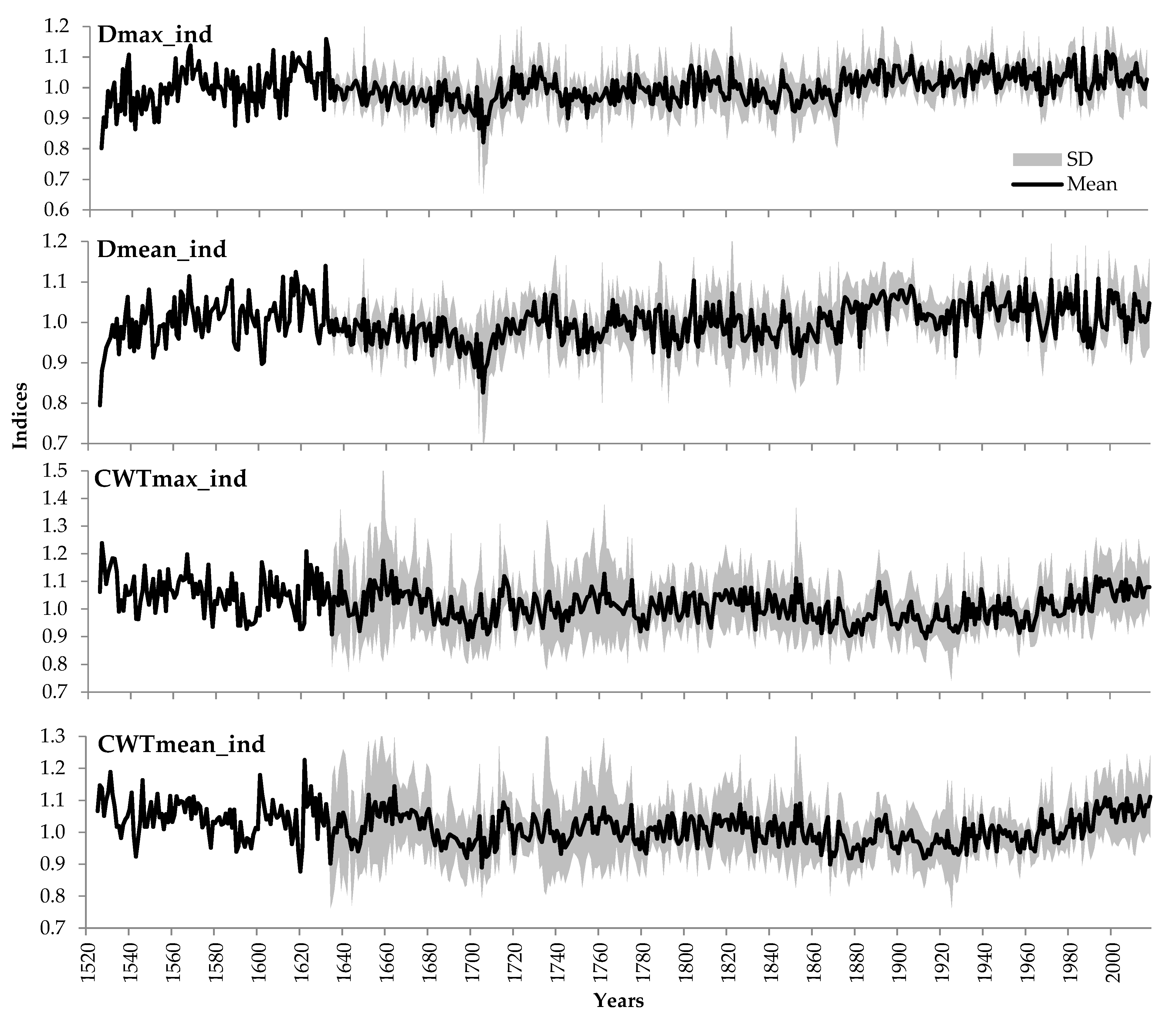
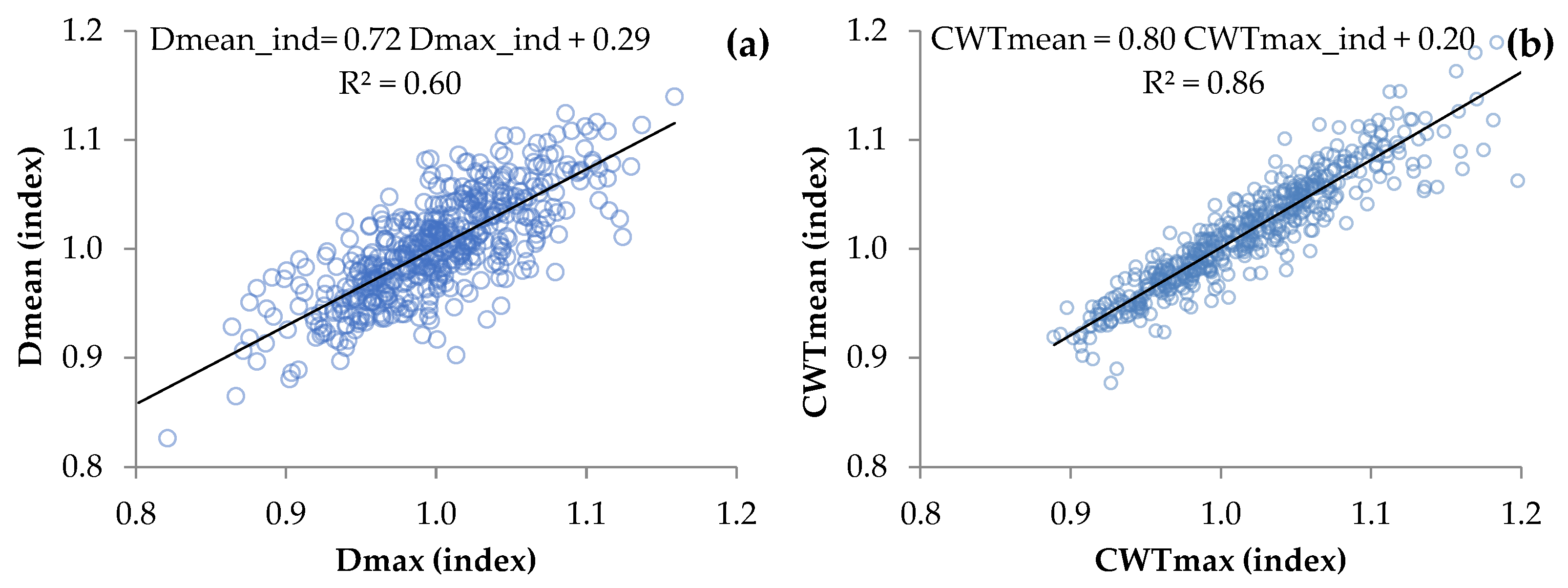
3.1. Relationships between Tree-Ring Traits
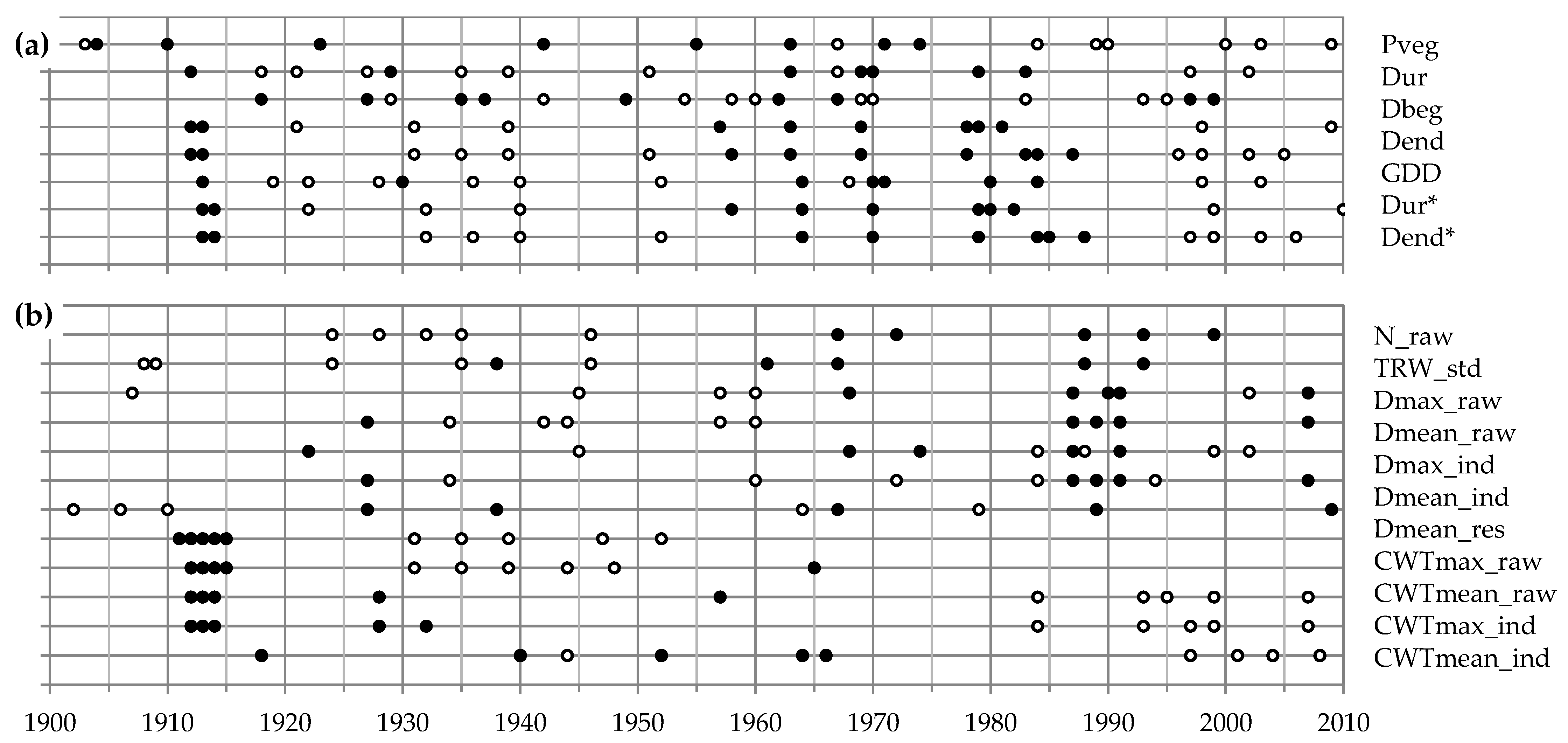
3.2. Climatic Response of Tree Ring Traits
4. Discussion
4.1. Quantitative Description of Relationships within Tree-Ring Structure
4.2. Climatic Response Patterns and Separation
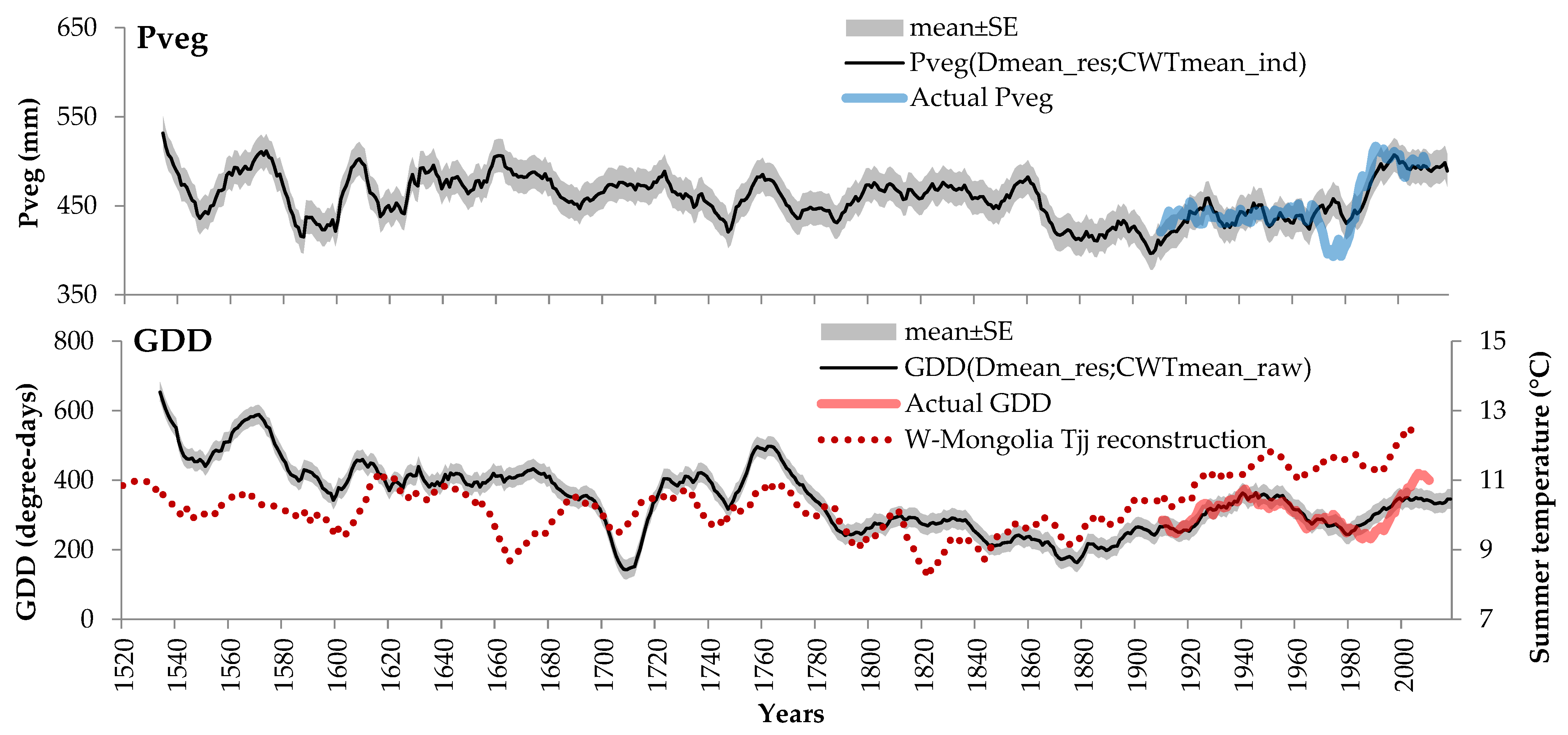
4.3. Reconstruction of the Climatic Characteristics of the Vegetative Season
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- White, A.; Cannell, M.G.; Friend, A.D. CO2 stabilization, climate change and the terrestrial carbon sink. Glob. Chang. Biol. 2000, 6, 817–833. [Google Scholar] [CrossRef]
- Parks, C.G.; Bernier, P. Adaptation of forests and forest management to changing climate with emphasis on forest health: A review of science, policies and practices. For. Ecol. Manag. 2010, 259, 657–659. [Google Scholar] [CrossRef]
- Pugh, T.A.; Lindeskog, M.; Smith, B.; Poulter, B.; Arneth, A.; Haverd, V.; Calle, L. Role of forest regrowth in global carbon sink dynamics. Proc. Natl. Acad. Sci. USA 2019, 116, 4382–4387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mérian, P.; Bontemps, J.D.; Bergés, L.; Lebourgeois, F. Spatial variation and temporal instability in climate-growth relationships of sessile oak (Quercus petraea [Matt.] Liebl.) under temperate conditions. Plant Ecol. 2011, 212, 1855–1871. [Google Scholar] [CrossRef]
- Crimmins, S.M.; Dobrowski, S.Z.; Greenberg, J.A.; Abatzoglou, J.T.; Mynsberge, A.R. Changes in climatic water balance drive downhill shifts in plant species’ optimum elevations. Science 2011, 331, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.U.; Shen, M.; Zhu, X. Plant phenology and global climate change: Current progresses and challenges. Glob. Chang. Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Tripathi, S.K. The need for establishing long-term ecological research stations network in India. Curr. Sci. 2010, 98, 21–22. [Google Scholar]
- Shiyatov, S.G. Dendrochronology, its principles and methods. In Problems of Botany in the Urals. Notes of the Soviet Union Botanical Society; Ural Scientific Center of the USSR Academy of Sciences: Sverdlovsk, Russia, 1973; Volume 6, pp. 53–81. (In Russian) [Google Scholar]
- Upadhyay, K.K.; Shah, S.K.; Roy, A.; Tripathi, S.K. Dendroclimatology of teak indicates prevailing climatic conditions of tropical moist forests in India. Ecol. Indic. 2021, 129, 107888:1–107888:11. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990. [Google Scholar]
- Brodersen, C.R.; Germino, M.J.; Johnson, D.M.; Reinhardt, K.; Smith, W.K.; Resler, L.M.; Bader, M.Y.; Sala, A.; Kueppers, L.M.; Broll, G.; et al. Seedling survival at timberline is critical to conifer mountain forest elevation and extent. Front. For. Glob. Chang. 2019, 2, 9. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin, Germany, 2003. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D. An updated list of species used in tree-ring research. Tree-Ring Bull. 1993, 53, 17–43. [Google Scholar]
- Vaganov, E.A.; Shiyatov, S.G.; Mazepa, V.S. Dendroclimatic Research in the Ural-Siberian Subarctic; Nauka: Novosibirsk, Russia, 1996. (In Russian) [Google Scholar]
- D’Arrigo, R.; Frank, D.; Pederson, N.; Cook, E.; Buckley, B.; Nachin, B.; Mijiddorj, R.; Dugarjav, C. 1738 years of Mongolian temperature variability inferred from a tree-ring width chronology of Siberian pine. Geophys. Res. Lett. 2001, 28, 543–546. [Google Scholar] [CrossRef] [Green Version]
- Gerasimova, O.V.; Zharnikov, Z.Y.; Knorre, A.A.; Myglan, V.S. Climaticaly induced dynamic of radial growth of Siberian stone pine and Siberian fir in the mountaintaiga belt in “Ergaki” National Park. J. Sib. Fed. Univ. Biol. 2010, 3, 18–29. (In Russian) [Google Scholar]
- Nikolaeva, S.A.; Savchuk, D.A.; Bocharov, A.Y. Influence of Different Factors on Tree Growth of Pinus sibirica in the Highlands of the Central Altai Mountains. J. Sib. Fed. Univ. Biol. 2015, 8, 299–318. (In Russian) [Google Scholar] [CrossRef]
- Carrer, M.; Unterholzner, L.; Castagneri, D. Wood anatomical traits highlight complex temperature influence on Pinus cembra at high elevation in the Eastern Alps. Int. J. Biometeorol. 2018, 62, 1745–1753. [Google Scholar] [CrossRef]
- Wang, H.; Shao, X.M.; Jiang, Y.; Fang, X.Q.; Wu, S.H. The impacts of climate change on the radial growth of Pinus koraiensis along elevations of Changbai Mountain in northeastern China. For. Ecol. Manag. 2013, 289, 333–340. [Google Scholar] [CrossRef]
- Yu, D.; Liu, J.; Lewis, B.J.; Li, Z.; Wangming, Z.; Xiangmin, F.; Yawei, W.; Shengwei, J.; Limin, D. Spatial variation and temporal instability in the climate–growth relationship of Korean pine in the Changbai Mountain region of Northeast China. For. Ecol. Manag. 2013, 300, 96–105. [Google Scholar] [CrossRef]
- Cerrato, R.; Salvatore, M.C.; Gunnarson, B.E.; Linderholm, H.W.; Carturan, L.; Brunetti, M.; De Blasi, F.; Baroni, C. A Pinus cembra L. tree-ring record for late spring to late summer temperature in the Rhaetian Alps, Italy. Dendrochronologia 2019, 53, 22–31. [Google Scholar] [CrossRef]
- Hantemirov, R.M.; Shiyatov, S.G. A continuous multimillennial ring-width chronology in Yamal, northwestern Siberia. Holocene 2002, 12, 717–726. [Google Scholar] [CrossRef]
- Khansaritoreh, E.; Dulamsuren, C.; Klinge, M.; Ariunbaatar, T.; Bat-Enerel, B.; Batsaikhan, G.; Ganbaatar, K.; Saindovdon, D.; Yeruult, Y.; Tsogtbaatar, J.; et al. Higher climate warming sensitivity of Siberian larch in small than large forest islands in the fragmented Mongolian forest steppe. Glob. Chang. Biol. 2017, 23, 3675–3689. [Google Scholar] [CrossRef]
- Zhirnova, D.F.; Belokopytova, L.V.; Babushkina, E.A.; Crivellaro, A.; Vaganov, E.A. Earlywood structure of evergreen conifers near forest line is habitat driven but latewood depends on species and seasons. Trees 2021, 35, 479–492. [Google Scholar] [CrossRef]
- Polikarpov, N.P.; Chebakova, N.M.; Nazimova, D.I. Climate and Mountain Forests of Southern Siberia; Nauka: Novosibirsk, Russia, 1986. (In Russian) [Google Scholar]
- Chytrý, M.; Danihelka, J.; Kubešová, S.; Lustyk, P.; Ermakov, N.; Hájek, M.; Hájková, P.; Kočí, M.; Otýpková, Z.; Roleček, J.; et al. Diversity of forest vegetation across a strong gradient of climatic continentality: Western Sayan Mountains, southern Siberia. Plant Ecol. 2008, 196, 61–83. [Google Scholar] [CrossRef]
- Timoshok, E.E.; Skorokhodov, S.N.; Timoshok, E.N. Ecological and coenotic description of Siberian stone pine (Pinus sibirica Du Tour) at the highest line of its distribution in the Central Altai. Tomsk State Univ. J. Biol. 2012, 4, 171–184. (In Russian) [Google Scholar] [CrossRef]
- Petrova, E.A.; Goroshkevich, S.N.; Belokon, M.M.; Belokon, Y.S.; Politov, D.V. Distribution of the genetic diversity of the Siberian stone pine, Pinus sibirica Du Tour, along the latitudinal and longitudinal profiles. Russ. J. Genet. 2014, 50, 467–482. [Google Scholar] [CrossRef]
- Kulagin, A.Y.; Davydychev, A.N.; Zaitsev, G.A. Specific features of the growth of Siberian spruce (Picea obovata Ledeb.) at early stages of ontogeny in broadleaf-conifer forests of the Ufa plateau. Russ. J. Ecol. 2006, 37, 66–69. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Ranson, K.J.; Im, S.T.; Dvinskaya, M.L. Response of Pinus sibirica and Larix sibirica to climate change in southern Siberian alpine forest–tundra ecotone. Scand. J. For. Res. 2009, 24, 130–139. [Google Scholar] [CrossRef]
- Monnier, Y.; Prévosto, B.; Ripert, C.; Corbani, A.C.; Fernandez, C. Forest microhabitats differentially influence seedling phenology of two co-existing Mediterranean oak species. J. Veg. Sci. 2012, 23, 260–270. [Google Scholar] [CrossRef]
- Zhirnova, D.F.; Babushkina, E.A.; Belokopytova, L.V.; Yurin, D.O.; Vaganov, E.A. Sunshine as culprit: It induces early spring physiological drought in dark coniferous (Pinus sibirica and Abies sibirica) alpine forest. For. Ecol. Manag. 2019, 449, 117458. [Google Scholar] [CrossRef] [Green Version]
- Zhirnova, D.F.; Belokopytova, L.V.; Meko, D.M.; Babushkina, E.A.; Vaganov, E.A. Climate change and tree growth in the Khakass-Minusinsk Depression (South Siberia) impacted by large water reservoirs. Sci. Rep. 2021, 11, 14266. [Google Scholar] [CrossRef]
- Büntgen, U. Re-thinking the boundaries of dendrochronology. Dendrochronologia 2019, 53, 1–4. [Google Scholar] [CrossRef]
- Wang, H.; Shao, X.; Fang, X.; Jiang, Y.; Liu, C.; Qiao, Q. Relationships between tree-ring cell features of Pinus koraiensis and climate factors in the Changbai Mountains, Northeastern China. J. For. Res. 2017, 28, 105–114. [Google Scholar] [CrossRef]
- Fonti, P.; von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Panyushkina, I.P.; Hughes, M.K.; Vanganov, E.A.; Munro, M.A.R. Summer temperature in northeastern Siberia since 1642 reconstructed from tracheid dimensions and cell numbers of Larix cajanderi. Can. J. For. Res. 2003, 33, 1905–1914. [Google Scholar] [CrossRef]
- Wang, H.; Shao, X.M.; Fang, X.Q.; Yin, Z.Y.; Chen, L.; Zhao, D.S.; Wu, S.H. Responses of Pinus koraiensis tree ring cell scale parameters to climate elements in the Changbai Mountains. China J. Plant. Ecol. 2011, 22, 2643–2652. [Google Scholar]
- Ziaco, E.; Biondi, F.; Heinrich, I. Wood cellular dendroclimatology: Testing new proxies in Great Basin bristlecone pine. Front. Plant Sci. 2016, 7, 1602. [Google Scholar] [CrossRef] [Green Version]
- Balanzategui, D.; Nordhauß, H.; Heinrich, I.; Biondi, F.; Miley, N.; Hurley, A.G.; Ziaco, E. Wood Anatomy of Douglas-Fir in Eastern Arizona and Its Relationship with Pacific Basin Climate. Front. Plant Sci. 2021, 12, 702442:1–702442:14. [Google Scholar] [CrossRef]
- Frank, D.A.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob. Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Larson, P.R. The Vascular Cambium. Development and Structure; Springer: Berlin/Heidelberg, Germany, 1994. [Google Scholar]
- Vaganov, E.A.; Hughes, M.K.; Shashkin, A.V. Growth Dynamics of Conifer Tree Rings: Images of Past and Future Environments; Springer: Dordrecht, Germany, 2006. [Google Scholar]
- Compo, G.P.; Whitaker, J.S.; Sardeshmukh, P.D.; Matsui, N.; Allan, R.J.; Yin, X.; Gleason, B.E.; Vose, R.S.; Rutledge, G.; Bessemoulin, P.; et al. The Twentieth Century Reanalysis Project. Q. J. Roy. Meteorol. Soc. 2011, 137, 1–28. [Google Scholar] [CrossRef]
- Polikarpov, N.P.; Nazimova, D.I. The dark coniferous forests of the northern part of the west Siberian mountains. In Forestry Research in the Forests of Siberia; Institute for Forests and Wood: Krasnoyarsk, Russia, 1963; pp. 103–147. (In Russian) [Google Scholar]
- Monserud, R.A.; Tchebakova, N.M. A vegetation model for the Sayan Mountains, southern Siberia. Can. J. For. Res. 1996, 26, 1055–1068. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 68–78. [Google Scholar]
- Rinn, F. TSAP-Win: Time Series Analysis and Presentation for Dendrochronology and Related Applications: User Reference; RINNTECH: Heidelberg, Germany, 2003. [Google Scholar]
- Cook, E.R.; Krusic, P.J. Program ARSTAN: A Tree-Ring Standardization Program Based on Detrending and Autoregressive Time Series Modeling, with Interactive Graphics; Lamont-Doherty Earth Observatory, Columbia University: Palisades, NY, USA, 2005. [Google Scholar]
- Silkin, P.P. Methods of Multiparameter Analysis of Conifers Tree-Rings Structure; Siberian Federal University: Krasnoyarsk, Russia, 2010. (In Russian) [Google Scholar]
- Seo, J.W.; Smiljanić, M.; Wilmking, M. Optimizing cell-anatomical chronologies of Scots pine by stepwise increasing the number of radial tracheid rows included—Case study based on three Scandinavian sites. Dendrochronologia 2014, 32, 205–209. [Google Scholar] [CrossRef]
- Vaganov, E.A. The tracheidogram method in tree-ring analysis and its application. In Methods of Dendrochronology. Application in Environmental Sciences; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, Germany, 1990; pp. 63–75. [Google Scholar]
- Wilks, D.S. Statistical Methods in the Atmospheric Sciences; Elsevier: Cambridge, UK, 2019. [Google Scholar]
- Methodological Guidelines for Compiling an Agrometeorological Yearbook for the Agricultural Zone of the Russian Federation; Guiding Document 52.33.725-2010.; Roshydromet: Obninsk, Russia, 2010. (In Russian)
- Sadokov, V.P.; Kozeltseva, V.F.; Kuznetsova, N.N. Determination of spring dates for a stable transition of the average daily air temperature through 0, +5 °C, their forecast and assessment. Proc. Hydrometeorol. Res. Cent. Russ. Fed. 2012, 348, 162–172. (In Russian) [Google Scholar]
- Schweingruber, F.H.; Eckstein, D.; Serre-Bachet, F.; Braker, O.U. Identification, presentation and interpretation of event years and pointer years in dendrochronology. Dendrochronologia 1990, 8, 9–38. [Google Scholar]
- Meko, D.M.; Baisan, C.H. Pilot study of latewood width of conifers as an indicator of variability of summer rainfall in the North American monsoon region. Int. J. Climatol. J. R. Meteorol. Soc. 2001, 21, 697–708. [Google Scholar] [CrossRef]
- Davi, N.K.; Rao, M.P.; Wilson, R.; Andreu-Hayles, L.; Oelkers, R.; D’Arrigo, R.; Nachin, B.; Buckley, B.; Pederson, N.; Leland, C.; et al. Accelerated recent warming and temperature variability over the past eight centuries in the Central Asian Altai from blue intensity in tree rings. Geophys. Res. Lett. 2021, 48, e2021GL092933. [Google Scholar] [CrossRef]
- Darikova, Y.A.; Vaganov, E.A.; Kuznetsova, G.V.; Grachev, A.M. Changes in the anatomical structure of tree rings of the rootstock and scion in the heterografts of Siberian pine. Trees 2013, 27, 1621–1631. [Google Scholar] [CrossRef]
- Matisons, R.; Krišāns, O.; Kārkliņa, A.; Adamovičs, A.; Jansons, Ā.; Gärtner, H. Plasticity and climatic sensitivity of wood anatomy contribute to performance of eastern Baltic provenances of Scots pine. For. Ecol. Manag. 2019, 452, 117568. [Google Scholar] [CrossRef]
- Mencuccini, M.; Grace, J.; Fioravanti, M. Biomechanical and hydraulic determinants of tree structure in Scots pine: Anatomical characteristics. Tree Physiol. 1997, 17, 105–113. [Google Scholar] [CrossRef]
- Kašpar, J.; Anfodillo, T.; Treml, V. Tree size mostly drives the variation of xylem traits at the treeline ecotone. Trees 2019, 33, 1657–1665. [Google Scholar] [CrossRef]
- Van Do, T.; Sato, T.; Saito, S.; Kozan, O.; Yamagawa, H.; Nagamatsu, D.; Nishimura, N.; Manabe, T. Effects of micro-topographies on stand structure and tree species diversity in an old-growth evergreen broad-leaved forest, southwestern Japan. Glob. Ecol. Conserv. 2015, 4, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Pritzkow, C.; Heinrich, I.; Grudd, H.; Helle, G. Relationship between wood anatomy, tree-ring widths and wood density of Pinus sylvestris L. and climate at high latitudes in northern Sweden. Dendrochronologia 2014, 32, 295–302. [Google Scholar] [CrossRef]
- Kirdyanov, A.V.; Vaganov, E.A.; Hughes, M.K. Separating the climatic signal from tree-ring width and maximum latewood density records. Trees 2007, 21, 37–44. [Google Scholar] [CrossRef]
- Björklund, J.; Seftigen, K.; Schweingruber, F.; Fonti, P.; von Arx, G.; Bryukhanova, M.V.; Cuny, H.E.; Carrer, M.; Castagneri, D.; Frank, D.C. Cell size and wall dimensions drive distinct variability of earlywood and latewood density in Northern Hemisphere conifers. New Phytol. 2017, 216, 728–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathgeber, C.B. Conifer tree-ring density interannual variability–anatomical, physiological and environmental determinants. New Phytol. 2017, 216, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Babushkina, E.A.; Dergunov, D.R.; Belokopytova, L.V.; Zhirnova, D.F.; Upadhyay, K.K.; Tripathi, S.K.; Zharkov, M.S.; Vaganov, E.A. Non-linear response to cell number revealed and eliminated from long-term tracheid measurements of Scots pine in Southern Siberia. Front. Plant Sci. 2021, 12, 719796. [Google Scholar] [CrossRef]
- Mork, E. Die Qualitat des Fichtenhozes unter besonder Rucksichtnahme auf Schleif und Papierholz. Der Pap. Fabr. 1928, 26, 741–747. (In German) [Google Scholar]
- Denne, M. Definition of latewood according to Mork. IAWA Bull. 1989, 10, 59–62. [Google Scholar] [CrossRef]
- Naurzbaev, M.M.; Vaganov, E.A. Variation of early summer and annual temperature in east Taymir and Putoran (Siberia) over the last two millennia inferred from tree rings. J. Geophys. Res. Atmos. 2000, 105, 7317–7326. [Google Scholar] [CrossRef]
- Oberhuber, W.; Kofler, W.; Pfeifer, K.; Seeber, A.; Gruber, A.; Wieser, G. Long-term changes in tree-ring–climate relationships at Mt. Patscherkofel (Tyrol, Austria) since the mid-1980s. Trees 2008, 22, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Jochner, M.; Bugmann, H.; Nötzli, M.; Bigler, C. Tree growth responses to changing temperatures across space and time: A fine-scale analysis at the treeline in the Swiss Alps. Trees 2018, 32, 645–660. [Google Scholar] [CrossRef]
- King, G.; Fonti, P.; Nievergelt, D.; Büntgen, U.; Frank, D. Climatic drivers of hourly to yearly tree radius variations along a 6 degrees C natural warming gradient. Agric. For. Meteorol. 2013, 168, 36–46. [Google Scholar] [CrossRef]
- Gindl, W.; Grabner, M.; Wimmer, R. Effects of altitude on tracheid differentiation and lignification of Norway spruce. Can. J. Bot. 2001, 79, 815–821. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; King, G.M.; et al. A meta-analysis of cambium phenology and growth: Linear and non-linear patterns in conifers of the northern hemisphere. Ann. Bot. 2013, 112, 1911–1920. [Google Scholar] [CrossRef]
- Begum, S.; Nakaba, S.; Yamagishi, Y.; Oribe, Y.; Funada, R. Regulation of cambial activity in relation to environmental conditions: Understanding the role of temperature in wood formation of trees. Physiol. Plant. 2013, 147, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Moreau, G.; Chagnon, C.; Auty, D.; Caspersen, J.; Achim, A. Impacts of climatic variation on the growth of black spruce across the forest-tundra ecotone: Positive effects of warm growing seasons and heat waves are offset by late spring frosts. Front. For. Glob. Chang. 2020, 3, 145. [Google Scholar] [CrossRef]
- Searls, T.; Zhu, X.; McKenney, D.W.; Mazumder, R.; Steenberg, J.; Yan, G.; Meng, F.R. Assessing the influence of climate on the growth rate of boreal tree species in northeastern Canada through long-term permanent sample plot data sets. Can. J. For. Res. 2021, 51, 1039–1049. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Carbohydrate sources and sinks in woody plants. Bot. Rev. 1992, 58, 107–222. [Google Scholar] [CrossRef]
- Pallardy, S.G. Physiology of Woody Plant, 3rd ed.; Academic Press: San Diego, CA, USA, 2010. [Google Scholar]
- Bazzoffi, P.; Nieddu, S. Effects of water logging on the soil structure of some Italian soils in relation to the GAEC cross-compliance standard maintenance of farm channel networks and field convexity. Ital. J. Agron. 2011, 6 (Suppl. S1), 63. [Google Scholar] [CrossRef] [Green Version]
- Yin, D.; Xu, D.; Tian, K.; Xiao, D.; Zhang, W.; Sun, D.; Sun, H.; Zhang, Y. Radial growth response of Abies georgei to climate at the upper timberlines in central Hengduan Mountains, Southwestern China. Forests 2018, 9, 606. [Google Scholar] [CrossRef] [Green Version]
- Mayr, S.; Wolfschwenger, M.; Bauer, H. Winter-drought induced embolism in Norway spruce (Picea abies) at the Alpine timberline. Physiol. Plant. 2002, 115, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Mayr, S.; Schmid, P.; Rosner, S. Winter embolism and recovery in the conifer shrub Pinus mugo L. Forests 2019, 10, 941. [Google Scholar] [CrossRef] [Green Version]
- D’Arrigo, R.D.; Jacoby, G.C.; Free, R.M. Tree-ring width and maximum latewood density at the North American tree line: Parameters of climatic change. Can. J. For. Res. 1992, 22, 1290–1296. [Google Scholar] [CrossRef]
- Zheng, L.; Gaire, N.P.; Shi, P. High-altitude tree growth responses to climate change across the Hindu Kush Himalaya. J. Plant Ecol. 2021, 14, 829–842. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B.; Frank, D.; Fonti, P.; Fournier, M. Kinetics of tracheid development explain conifer tree-ring structure. New Phytol. 2014, 203, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Cuny, H.E.; Rathgeber, C.B.; Frank, D.; Fonti, P.; Mäkinen, H.; Prislan, P.; Rossi, S.; del Castillo, E.M.; Campelo, F.; Vavrčík, H.; et al. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 2015, 1, 15160:1–15160:6. [Google Scholar] [CrossRef] [PubMed]
- Rathgeber, C.B.; Cuny, H.E.; Fonti, P. Biological basis of tree-ring formation: A crash course. Front. Plant Sci. 2016, 7, 734:1–734:7. [Google Scholar] [CrossRef] [Green Version]
- Blasing, T.J.; Duvick, D.N.; Cook, E.R. Filtering the effect of competition from ring-width series. Tree-Ring Bull. 1983, 43, 19–30. [Google Scholar]
- Camarero, J.J.; Colangelo, M.; Gazol, A.; Pizarro, M.; Valeriano, C.; Igual, J.M. Effects of windthrows on forest cover, tree growth and soil characteristics in drought-prone pine plantations. Forests 2021, 12, 817. [Google Scholar] [CrossRef]
- Yuan, N.; Xiong, F.; Xoplaki, E.; He, W.; Luterbacher, J. A new approach to correct the overestimated persistence in tree-ring width based precipitation reconstructions. Clim. Dyn. 2021, 1–12. [Google Scholar] [CrossRef]
- García-Cervigón, A.I.; Mercado, L.N.; Mendivelso, H.A.; Toledo, M.; Camarero, J.J. Adjusting xylem anatomy and growth to inter-annual climate variability in two Fabaceae species (Centrolobium microchaete, Cenostigma pluviosum) from Bolivian dry tropical forests. Dendrochronologia 2021, 67, 125840. [Google Scholar] [CrossRef]
- Pandey, S. Climatic influence on tree wood anatomy: A review. J. Wood. Sci. 2021, 67, 24. [Google Scholar] [CrossRef]
- Obeltsev, S.V.; Gustova, A.I.; Terekhina, D.K. Hydro- and thermophysical characteristics of wood of the main and shrub species of protective forest plantations of the steppe zone. Bull. Nizhnevolzhsky Agro-Univ. Complex Sci. High. Prof. Educ. 2011, 4, 91–97. (In Russian) [Google Scholar]
- Körner, C. Plant adaptation to cold climates. F1000Research 2016, 5 (F1000 Faculty Rev), 2769:1–2769:5. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B. The world-wide ‘fast–slow’plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Ning, Q.R.; Gong, X.W.; Li, M.Y.; Hao, G.Y. Differences in growth pattern and response to climate warming between Larix olgensis and Pinus koraiensis in Northeast China are related to their distinctions in xylem hydraulics. Agric. For. Meteorol. 2022, 312, 108724. [Google Scholar] [CrossRef]
- Neukom, R.; Barboza, L.A.; Erb, M.P.; Shi, F.; Emile-Geay, J.; Evans, M.N.; Franke, J.; Kaufman, D.S.; Lücke, L.; Rehfeld, K.; et al. Consistent multi-decadal variability in global temperature reconstructions and simulations over the Common Era. Nat. Geosci. 2019, 12, 643–649. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree | Diameter (cm) | Core Cover Period (Years) | Cell Number, N | TRW (mm) | |||
|---|---|---|---|---|---|---|---|
| Length | Calendar Years | Max | Mean ± 1SD | Max | Mean ± 1SD | ||
| Tree_1 | 46 | 493 | 1525–2018 | 31 | 12.0 ± 5.2 | 0.99 | 0.35 ± 0.16 |
| Tree_2 | 39 | 273 | 1746–2018 | 34 | 10.8 ± 7.5 | 1.04 | 0.33 ± 0.25 |
| Tree_3 | 61 | 327 | 1646–1972 | 49 | 26.8 ± 8.2 | 1.60 | 0.84 ± 0.26 |
| Tree_4 | 42 | 234 | 1785–2018 | 35 | 14.3 ± 5.0 | 1.12 | 0.44 ± 0.18 |
| Tree_5 | 47 | 387 | 1634–2018 | 43 | 18.5 ± 7.5 | 1.45 | 0.58 ± 0.25 |
| Tree_6 | 37 | 366 | 1653–2018 | 25 | 8.1 ± 4.7 | 0.74 | 0.21 ± 0.14 |
| Tree_7 | 38 | 336 | 1683–2018 | 39 | 17.2 ± 7.1 | 1.20 | 0.52 ± 0.23 |
| Tree | Dmax | Dmean | CWTmax | CWTmean | ||||
|---|---|---|---|---|---|---|---|---|
| Das (μm) | a | Das (μm) | a | CWTas (μm) | a | CWTas (μm) | a | |
| Tree_1 | 39.3 | 0.34 | 29.8 | 0.34 | 2.79 | 0.13 | 2.48 | 0.14 |
| Tree_2 | 41.8 | 0.34 | 31.7 | 0.29 | 3.55 | 0.09 | 2.92 | 0.11 |
| Tree_3 | 40.7 | 0.23 | 31.9 | 0.21 | 2.83 | 0.09 | 2.41 | 0.09 |
| Tree_4 | 42.5 | 0.20 | 33.0 | 0.19 | 2.24 | 0.33 | 2.00 | 0.44 |
| Tree_5 | 41.0 | 0.24 | 32.2 | 0.24 | 2.84 | 0.20 | 2.50 | 0.24 |
| Tree_6 | 35.4 | 0.38 | 28.0 | 0.30 | 2.36 | 0.30 | 2.07 | 0.49 |
| Tree_7 | 39.6 | 0.28 | 30.4 | 0.29 | 2.76 | 0.15 | 2.41 | 0.16 |
| Total sample | 40.0 | 0.30 | 31.0 | 0.26 | 2.77 | 0.14 | 2.40 | 0.18 |
| Statistics | Dmax | Dmean | CWTmax | CWTmean | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Indiv. | Chron. | Total | Indiv. | Chron. | Total | Indiv. | Chron. | Total | Indiv. | Chron. | Total | |
| Measurements | ||||||||||||
| Min (μm) | 11.3–29.7 | 27.4 | 11.3 | 10.8–24.0 | 21.5 | 10.8 | 1.51–1.90 | 1.90 | 1.51 | 0.49–1.76 | 1.76 | 1.45 |
| Max (μm) | 43.4–52.0 | 44.6 | 52.0 | 31.8–37.9 | 33.6 | 37.9 | 2.64–4.45 | 3.28 | 4.45 | 1.24–3.65 | 2.92 | 3.65 |
| Median (μm) | 33.5–40.5 | 38.3 | 38.5 | 25.6–31.8 | 29.5 | 29.8 | 2.11–2.73 | 2.48 | 2.37 | 1.00–2.25 | 2.24 | 2.15 |
| Mean (μm) | 32.4–40.5 | 38.2 | 38.0 | 24.5–31.7 | 29.3 | 29.2 | 2.11–2.73 | 2.50 | 2.44 | 1.00–2.27 | 2.25 | 2.20 |
| SD (μm) | 3.1–4.9 | 2.5 | 4.6 | 2.1–4.0 | 1.9 | 3.7 | 0.18–0.57 | 0.23 | 0.42 | 0.07–0.30 | 0.18 | 0.30 |
| Skewness | −1.12–0.24 | −0.53 | −0.97 | −1.30–−0.04 | −0.63 | −1.12 | −0.08–1.70 | 0.51 | 1.00 | −0.27–0.75 | 0.44 | 0.75 |
| Kurtosis | −0.44–5.53 | 1.22 | 2.23 | −0.24–5.05 | 1.18 | 1.99 | −0.48–2.55 | 0.46 | 1.69 | 0.23–2.89 | 0.42 | 0.94 |
| Indices | ||||||||||||
| Min (μm) | 0.49–0.82 | 0.80 | 0.49 | 0.59–0.79 | 0.79 | 0.59 | 0.72–0.81 | 0.89 | 0.72 | 0.71–0.82 | 0.88 | 0.71 |
| Max (μm) | 1.24–1.37 | 1.16 | 1.37 | 1.20–1.44 | 1.14 | 1.44 | 1.27–1.67 | 1.24 | 1.67 | 1.23–1.67 | 1.23 | 1.67 |
| Median (μm) | 0.99–1.01 | 1.00 | 1.00 | 1.00–1.01 | 1.00 | 1.00 | 0.98–1.02 | 1.02 | 1.00 | 0.99–1.01 | 1.01 | 1.00 |
| Mean (μm) | 1.00–1.01 | 1.00 | 1.00 | 1.00–1.01 | 1.00 | 1.00 | 0.99–1.03 | 1.02 | 1.01 | 0.99–1.02 | 1.01 | 1.01 |
| SD (μm) | 0.07–0.10 | 0.05 | 0.08 | 0.06–0.10 | 0.05 | 0.08 | 0.08–0.16 | 0.06 | 0.11 | 0.06–0.14 | 0.05 | 0.10 |
| Skewness | −0.59–0.35 | −0.06 | −0.10 | −0.36–0.02 | −0.11 | −0.25 | 0.19–0.78 | 0.44 | 0.68 | −0.02–0.62 | 0.38 | 0.60 |
| Kurtosis | 0.21–2.89 | 0.46 | 1.76 | 0.10–2.25 | 0.50 | 1.78 | −0.01–0.89 | 0.05 | 1.29 | 0.06–1.10 | 0.20 | 1.78 |
| Correlations | Dmax | Dmean | CWTmax | CWTmean | N | TRW * |
|---|---|---|---|---|---|---|
| Measurements | ||||||
| between trees | −0.08–0.62 0.31 | −0.07–0.66 0.35 | −0.26–0.53 0.13 | −0.27–0.49 0.11 | −0.41–0.70 0.32 | −0.33–0.76 0.30 |
| with site chronology | 0.36–0.83 0.64 | 0.31–0.83 0.67 | 0.25–0.78 0.52 | 0.23–0.75 0.52 | 0.39–0.86 0.68 | −0.05–0.76 0.41 |
| Indices | ||||||
| between trees | 0.00–0.29 0.19 | 0.05–0.39 0.23 | −0.19–0.36 0.07 | −0.20–0.38 0.06 | – | 0.20–0.58 0.42 |
| with site chronology | 0.48–0.72 0.57 | 0.49–0.73 0.60 | 0.23–0.68 0.47 | 0.23–0.69 0.47 | – | 0.55–0.75 0.65 |
| Climatic Variable | Tree-Ring Chronology | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N_raw | TRW_std | Dmax_raw | Dmean_raw | CWTmax_raw | CWTmean_raw | Dmax_ind | Dmean_ind | CWTmax_ind | CWTmean_ind | Dmean_res | CWTmean_res | |
| Time series per se | ||||||||||||
| Pveg | −0.23 | −0.16 | −0.12 | −0.32 | −0.06 | −0.03 | 0.08 | −0.18 | 0.25 | 0.27 | −0.35 | 0.13 |
| Dur | 0.06 | 0.05 | 0.04 | −0.12 | 0.25 | 0.30 | 0.04 | −0.14 | 0.17 | 0.21 | −0.25 | 0.18 |
| Dbeg | −0.05 | 0.00 | −0.02 | 0.13 | −0.09 | −0.14 | −0.04 | 0.14 | −0.08 | −0.13 | 0.24 | −0.19 |
| Dend | 0.04 | 0.07 | 0.03 | −0.06 | 0.27 | 0.30 | 0.03 | −0.08 | 0.17 | 0.18 | −0.15 | 0.08 |
| GDD | 0.12 | 0.19 | 0.04 | −0.04 | 0.41 | 0.42 | 0.03 | −0.07 | 0.29 | 0.29 | −0.13 | 0.09 |
| Dur * | 0.21 | 0.21 | 0.00 | −0.07 | 0.21 | 0.18 | −0.05 | −0.15 | 0.07 | 0.05 | −0.17 | −0.05 |
| Dend * | 0.21 | 0.23 | 0.03 | −0.05 | 0.21 | 0.20 | −0.03 | −0.14 | 0.07 | 0.06 | −0.17 | −0.01 |
| GDD * | 0.17 | 0.21 | 0.02 | −0.09 | 0.32 | 0.29 | −0.01 | −0.15 | 0.22 | 0.20 | −0.21 | −0.01 |
| Time series smoothed by 11-year moving average | ||||||||||||
| Pveg | −0.44 | −0.08 | −0.37 | −0.57 | 0.03 | 0.19 | 0.32 | −0.22 | 0.67 | 0.71 | −0.69 | 0.62 |
| Dur | 0.30 | 0.44 | 0.23 | 0.04 | 0.65 | 0.71 | 0.40 | 0.05 | 0.32 | 0.33 | −0.33 | 0.16 |
| Dbeg | 0.01 | −0.20 | 0.01 | 0.17 | −0.37 | −0.44 | −0.34 | 0.02 | −0.40 | −0.41 | 0.36 | −0.31 |
| Dend | 0.45 | 0.53 | 0.34 | 0.17 | 0.73 | 0.78 | 0.38 | 0.08 | 0.22 | 0.23 | −0.25 | 0.03 |
| GDD | 0.30 | 0.43 | 0.23 | 0.08 | 0.69 | 0.74 | 0.39 | 0.09 | 0.41 | 0.41 | −0.24 | 0.10 |
| Dur * | 0.50 | 0.60 | 0.41 | 0.25 | 0.75 | 0.79 | 0.41 | 0.14 | 0.17 | 0.18 | −0.20 | 0.00 |
| Dend * | 0.35 | 0.52 | 0.30 | 0.11 | 0.68 | 0.73 | 0.44 | 0.10 | 0.28 | 0.30 | −0.29 | 0.13 |
| GDD * | 0.35 | 0.48 | 0.30 | 0.16 | 0.71 | 0.75 | 0.43 | 0.17 | 0.37 | 0.37 | −0.17 | 0.03 |
| Dependent Climatic Variable | Explanatory Anatomical Variables | R | R2 | R2adj | F(2.98) | p | SE |
|---|---|---|---|---|---|---|---|
| Pveg | Dmean_res, CWTmean_ind | 0.806 | 0.650 | 0.643 | 91.1 | <0.0001 | 18.9 |
| Dbeg | Dmean_res, CWTmean_raw | 0.579 | 0.335 | 0.321 | 24.7 | <0.0001 | 2.15 |
| Dur | Dmean_res, CWTmean_raw | 0.794 | 0.631 | 0.623 | 83.8 | <0.0001 | 3.69 |
| Dend | Dmean_res, CWTmax_raw | 0.813 | 0.662 | 0.655 | 95.8 | <0.0001 | 2.38 |
| GDD | Dmean_res, CWTmean_raw | 0.790 | 0.625 | 0.617 | 81.5 | <0.0001 | 28.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhirnova, D.F.; Belokopytova, L.V.; Upadhyay, K.K.; Tripathi, S.K.; Babushkina, E.A.; Vaganov, E.A. 495-Year Wood Anatomical Record of Siberian Stone Pine (Pinus sibirica Du Tour) as Climatic Proxy on the Timberline. Forests 2022, 13, 247. https://doi.org/10.3390/f13020247
Zhirnova DF, Belokopytova LV, Upadhyay KK, Tripathi SK, Babushkina EA, Vaganov EA. 495-Year Wood Anatomical Record of Siberian Stone Pine (Pinus sibirica Du Tour) as Climatic Proxy on the Timberline. Forests. 2022; 13(2):247. https://doi.org/10.3390/f13020247
Chicago/Turabian StyleZhirnova, Dina F., Liliana V. Belokopytova, Keshav K. Upadhyay, Shri K. Tripathi, Elena A. Babushkina, and Eugene A. Vaganov. 2022. "495-Year Wood Anatomical Record of Siberian Stone Pine (Pinus sibirica Du Tour) as Climatic Proxy on the Timberline" Forests 13, no. 2: 247. https://doi.org/10.3390/f13020247