
Is the Pinus massoniana Lamb. Tree-Ring Latewood Formation Influenced by the Diurnal Temperature Range in Humid Subtropical China?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
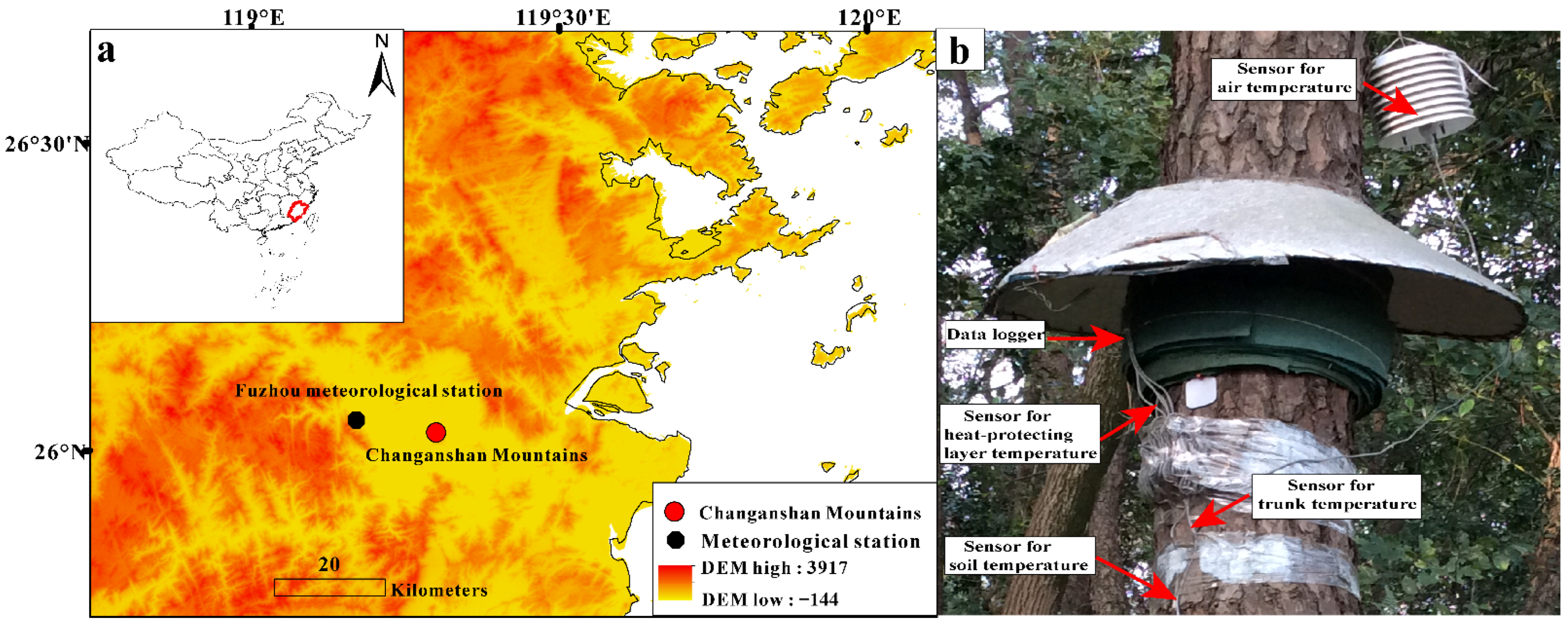
2.1. Study Region
2.2. Tree-Ring Width Chronology
2.3. Heat-Protecting Experiments on Tree Trunks
2.4. Tree-Ring Paraffin Section and Blue Intensity and Anatomical Structures of Tree-Ring Data
2.5. Climate Data
2.6. Analytical Methods
3. Results
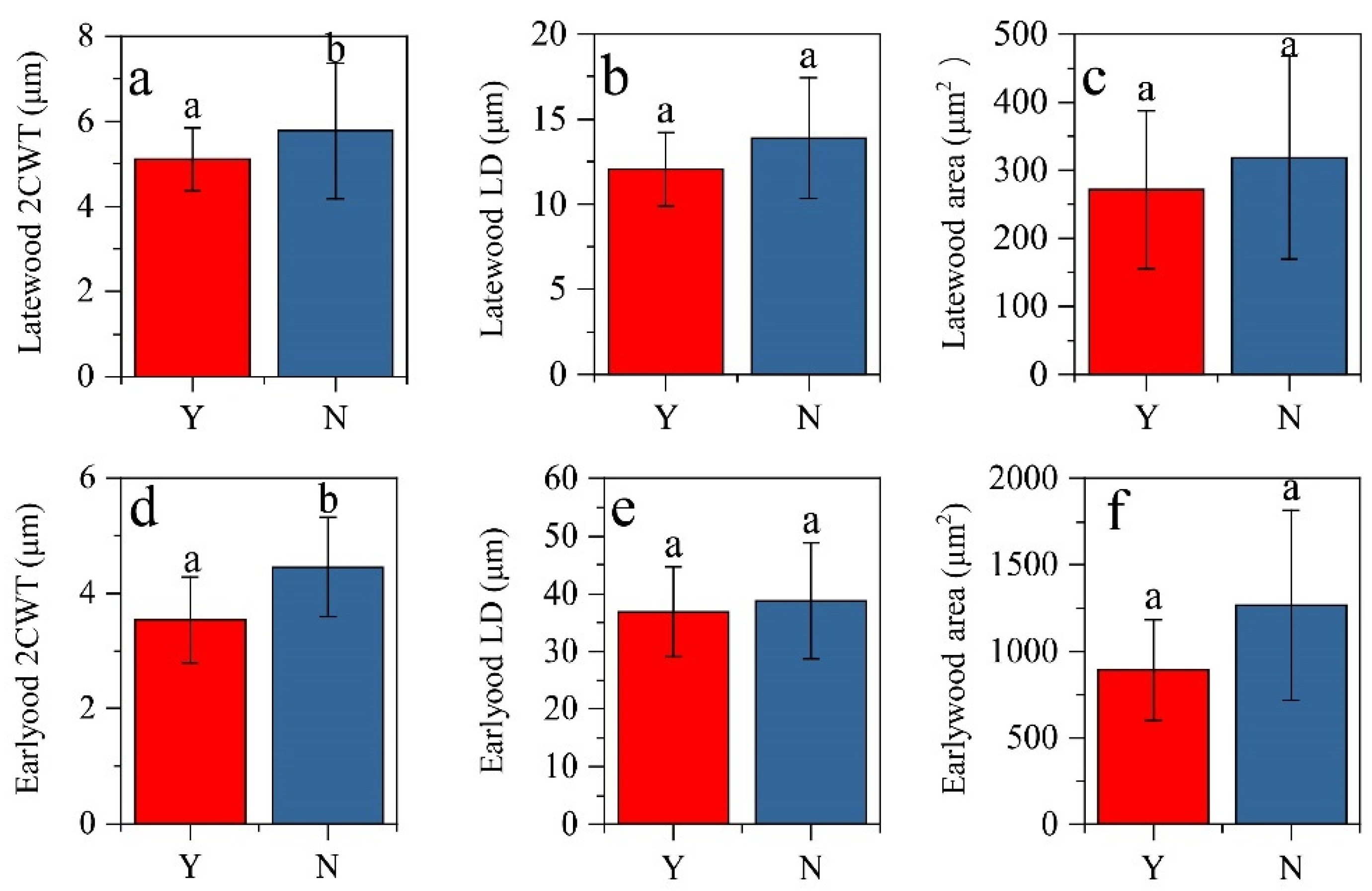
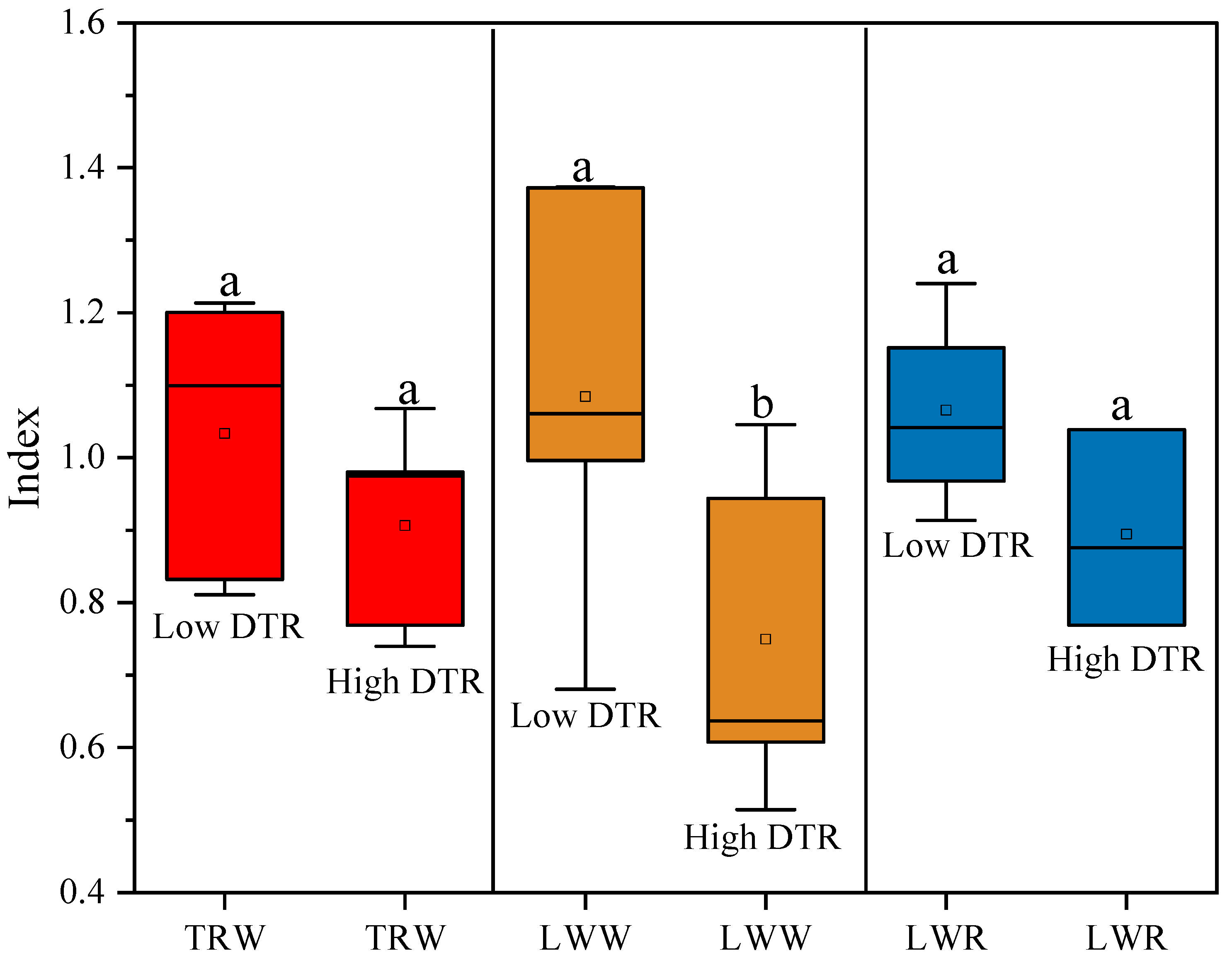
3.1. Wood Anatomy Changes after the Heat-Protecting
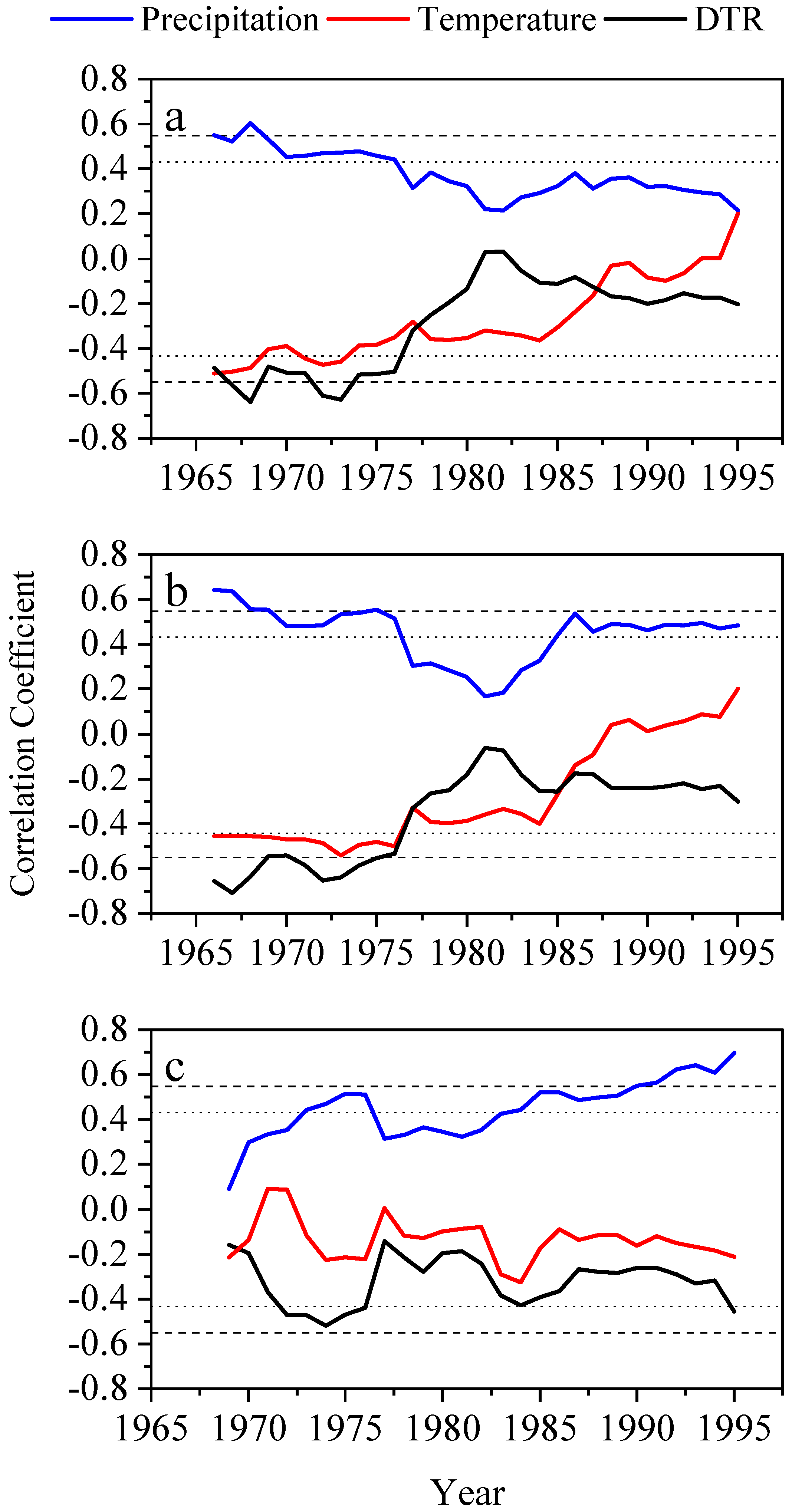
3.2. Tree-Ring Chronology and Climate–Growth Relationship
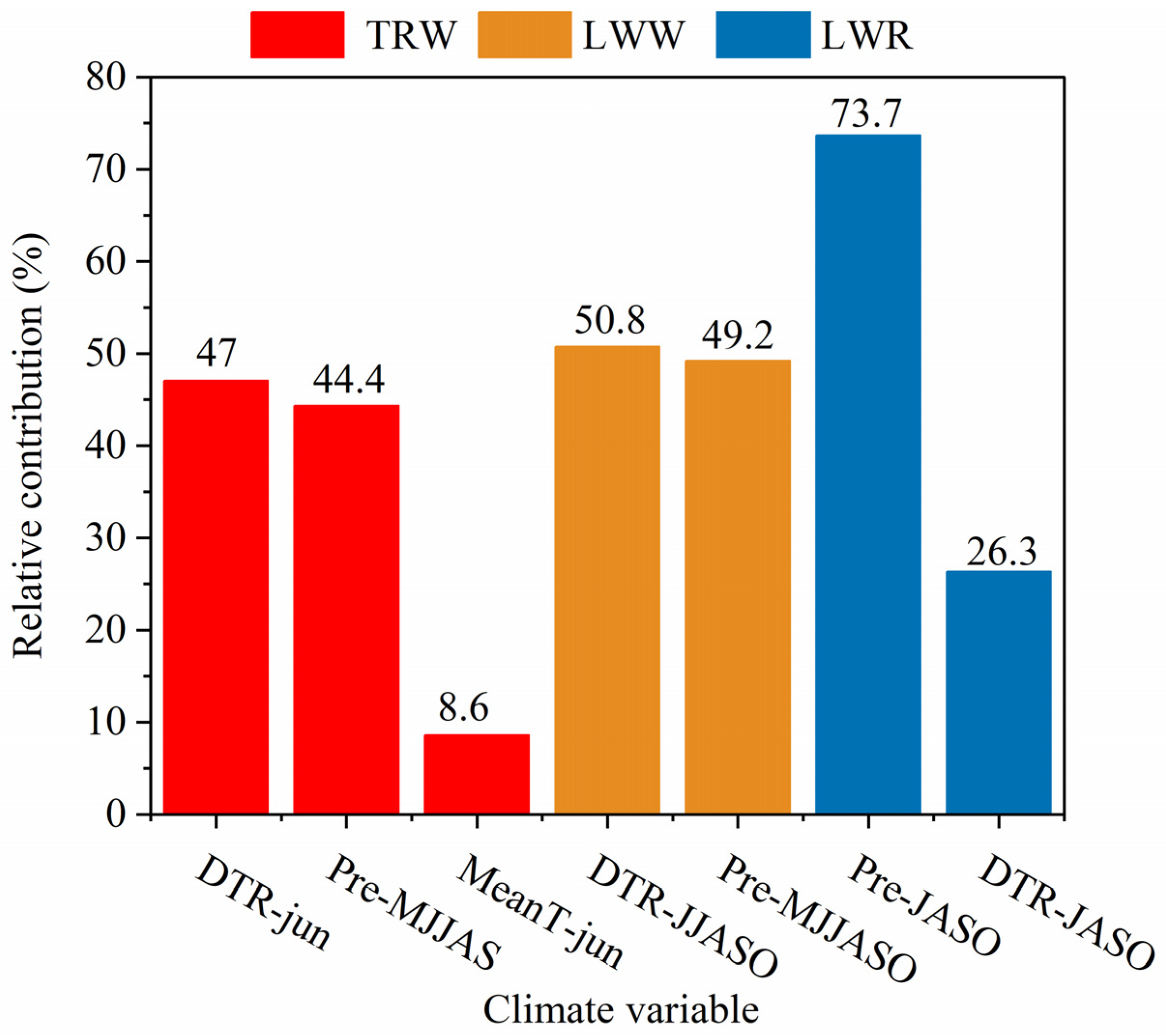
3.3. The Contribution of Climatic Factors in Climate–Growth Relationships
4. Discussion
4.1. The Effect of Heat-Protecting on Latewood Formation and Wood Anatomy
4.2. Tree Growth Relationship
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rutherford, S.; Mann, M.; Osborn, T.; Briffa, K.; Jones, P.D.; Bradley, R.; Hughes, M. Proxy-based Northern Hemisphere surface temperature reconstructions: Sensitivity to method, predictor network, target season, and target domain. J. Clim. 2005, 18, 2308–2329. [Google Scholar] [CrossRef]
- Fan, Z.-X.; Bräuning, A.; Yang, B.; Cao, K.-F. Tree ring density-based summer temperature reconstruction for the central Hengduan Mountains in southern China. Glob. Planet. Chang. 2009, 65, 1–11. [Google Scholar] [CrossRef]
- Anchukaitis, K.J.; Wilson, R.; Briffa, K.R.; Büntgen, U.; Cook, E.R.; D’Arrigo, R.; Davi, N.; Esper, J.; Frank, D.; Gunnarson, B.E. Last millennium Northern Hemisphere summer temperatures from tree rings: Part II, spatially resolved reconstructions. Quat. Sci. Rev. 2017, 163, 1–22. [Google Scholar] [CrossRef]
- Deng, G.; Li, M.; Hao, Z.; Shao, X. Responses to Climate Change of Maximum Latewood Density from Larix speciosa Cheng et Law and Abies delavayi Franch. in the Northwest of Yunnan Province, China. Forests 2022, 13, 720. [Google Scholar] [CrossRef]
- Yin, H.; Sun, Y.; Li, M.-Y. Reconstructed temperature change in late summer over the eastern Tibetan Plateau since 1867 CE and the role of anthropogenic forcing. Glob. Planet. Chang. 2022, 208, 103715. [Google Scholar] [CrossRef]
- Esper, J.; Düthorn, E.; Krusic, P.J.; Timonen, M.; Büntgen, U. Northern European summer temperature variations over the Common Era from integrated tree-ring density records. J. Quat. Sci. 2014, 29, 487–494. [Google Scholar] [CrossRef]
- Wu, P.; Wang, L.; Shao, X. Reconstruction of summer temperature variation from maximum density of alpine pine during 1917–2002 for west Sichuan Plateau, China. J. Geogr. Sci. 2008, 18, 201–210. [Google Scholar] [CrossRef]
- Schneider, L.; Smerdon, J.E.; Büntgen, U.; Wilson, R.J.S.; Myglan, V.S.; Kirdyanov, A.V.; Esper, J. Revising midlatitude summer temperatures back to A.D. 600 based on a wood density network. Geophys. Res. Lett. 2015, 42, 4556–4562. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Wilson, R.; Liepert, B.; Cherubini, P. On the ‘divergence problem’ in northern forests: A review of the tree-ring evidence and possible causes. Glob. Planet. Chang. 2008, 60, 289–305. [Google Scholar] [CrossRef]
- Leonelli, G.; Coppola, A.; Baroni, C.; Salvatore, M.C.; Maugeri, M.; Brunetti, M.; Pelfini, M. Multispecies dendroclimatic reconstructions of summer temperature in the European Alps enhanced by trees highly sensitive to temperature. Clim. Chang. 2016, 137, 275–291. [Google Scholar] [CrossRef]
- Qiu, T.; Sharma, S.; Woodall, C.W.; Clark, J.S. Niche Shifts From Trees to Fecundity to Recruitment That Determine Species Response to Climate Change. Front. Ecol. Evol. 2021, 9, 719141. [Google Scholar] [CrossRef]
- Castagneri, D.; Fonti, P.; von Arx, G.; Carrer, M. How does climate influence xylem morphogenesis over the growing season? Insights from long-term intra-ring anatomy in Picea abies. Ann. Bot. 2017, 119, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Yuichiro, O.; Takafumi, K. Effect of heat on cambial reactivation during winter dormancy in evergreen and deciduous conifers. Tree Physiol. 1997, 17, 81–87. [Google Scholar]
- Giovannelli, A.; Mattana, S.; Emiliani, G.; Anichini, M.; Traversi, M.L.; Pavone, F.S.; Cicchi, R. Localized stem heating from the rest to growth phase induces latewood-like cells formation and slower stem radial growth in Norway spruce saplings. Tree Physiol. 2021, 42, 1149–1163. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.D.; Montagu, D.K.; Conroy, P.J. Temperature effects on wood anatomy, wood density, photosynthesis and biomass partitioning of Eucalyptus grandis seedlings. Tree Physiol. 2007, 27, 251–260. [Google Scholar] [CrossRef]
- Mellerowicz, E.; Coleman, W.; Riding, R.; Little, C. Periodicity of cambial activity in Abies balsamea. I. Effects of temperature and photoperiod on cambial dormancy and frost hardiness. Physiol. Plant. 1992, 85, 515–525. [Google Scholar] [CrossRef]
- Eklund, L.; Little, C.A.; Riding, R.T. Concentrations of oxygen and indole-3-acetic acid in the cambial region during latewood formation and dormancy development in Picea abies stems. J. Exp. Bot. 1998, 49, 205–211. [Google Scholar]
- Babushkina, E.A.; Belokopytova, L.V.; Zhirnova, D.F.; Vaganov, E.A. Siberian spruce tree ring anatomy: Imprint of development processes and their high-temporal environmental regulation. Dendrochronologia 2019, 53, 114–124. [Google Scholar] [CrossRef]
- Campelo, F.; Nabais, C.; Freitas, H.; Gutiérrez, E. Climatic significance of tree-ring width and intra-annual density fluctuations in Pinus pinea from a dry Mediterranean area in Portugal. Ann. For. Sci. 2007, 64, 229–238. [Google Scholar] [CrossRef]
- Lebourgeois, F. Climatic signals in earlywood, latewood and total ring width of Corsican pine from western France. Ann. For. Sci. 2000, 57, 155–164. [Google Scholar] [CrossRef]
- Zahner, R.; Lotan, J.E.; Baughman, W.D. Earlywood-latewood features of red pine grown under simulated drought and irrigation. For. Sci. 1964, 10, 361–370. [Google Scholar]
- Pharis, R.; Jenkins, P.; Aoki, H.; Sassa, T. Hormonal physiology of wood growth in Pinus radiata D. Don: Effects of gibberellin A4 and the influence of abscisic acid upon (3H) gibberellin A4 metabolism. Funct. Plant Biol. 1981, 8, 559–570. [Google Scholar] [CrossRef]
- Davies, P.J. Plant Hormones: Physiology, Biochemistry and Molecular Biology; Springer Science & Business Media: New York, NY, USA, 2013. [Google Scholar]
- Funada, R.; Kubo, T.; Tabuchi, M.; Sugiyama, T.; Fushitani, M. Seasonal variations in endogenous indole-3-acetic acid and abscisic acid in the cambial region of Pinus densiflora Sieb. et Zucc. stems in relation to earlywood-latewood transition and cessation of tracheid production. Holzforschung 2001, 55, 128–134. [Google Scholar]
- Renninger, H.J.; Gartner, B.L.; Grotta, A.T. No correlation between latewood formation and leader growth in Douglas-fir saplings. Iawa J. 2006, 27, 183–191. [Google Scholar] [CrossRef]
- Funada, R.; Kubo, T.; Fushitani, M. Early-and latewood formation in Pinus densiflora trees with different amounts of crown. Iawa J. 1990, 11, 281–288. [Google Scholar] [CrossRef]
- Braganza, K.; Karoly, D.J.; Arblaster, J.M. Diurnal temperature range as an index of global climate change during the twentieth century. Geophys. Res. Lett. 2004, 31, L13217. [Google Scholar] [CrossRef]
- Zhang, X.; Manzanedo, R.D.; Lv, P.; Xu, C.; Hou, M.; Huang, X.; Rademacher, T. Reduced diurnal temperature range mitigates drought impacts on larch tree growth in North China. Sci. Total Environ. 2022, 848, 157808. [Google Scholar] [CrossRef]
- Büntgen, U.; Martínez-Peña, F.; Aldea, J.; Rigling, A.; Fischer, E.M.; Camarero, J.J.; Hayes, M.J.; Fatton, V.; Egli, S. Declining pine growth in Central Spain coincides with increasing diurnal temperature range since the 1970s. Glob. Planet. Chang. 2013, 107, 177–185. [Google Scholar] [CrossRef]
- Stoffel, M.; Corona, C.; Ballesteros-Cánovas, J.A.; Bodoque, J.M. Dating and quantification of erosion processes based on exposed roots. Earth-Sci. Rev. 2013, 123, 18–34. [Google Scholar] [CrossRef]
- Zhou, F.; Gou, X.; Zhang, J.; Zhao, Z.; Chen, Q.; Cao, Z. Application of Picea wilsonii roots to determine erosion rates in eastern Qilian Mountains, Northwest China. Trees 2013, 27, 371–378. [Google Scholar] [CrossRef]
- Friedman, J.M.; Vincent, K.R.; Shafroth, P.B. Dating floodplain sediments using tree-ring response to burial. Earth Surf. Process. Landf. 2010, 30, 1077–1091. [Google Scholar] [CrossRef]
- Thorne, P.; Donat, M.; Dunn, R.; Williams, C.; Alexander, L.; Caesar, J.; Durre, I.; Harris, I.; Hausfather, Z.; Jones, P. Reassessing changes in diurnal temperature range: Intercomparison and evaluation of existing global data set estimates. J. Geophys. Res. Atmos. 2016, 121, 5138–5158. [Google Scholar] [CrossRef]
- Huang, J.-G.; Guo, X.; Rossi, S.; Zhai, L.; Yu, B.; Zhan, S.; Zhang, M. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season. Tree Physiol. 2018, 38, 1225–1236. [Google Scholar] [CrossRef]
- Li, D.; Fang, K.; Li, Y.; Chen, D.; Liu, X.; Dong, Z.; Zhou, F.; Guo, G.; Shi, F.; Xu, C. Climate, intrinsic water-use efficiency and tree growth over the past 150 years in humid subtropical China. PLoS ONE 2017, 12, e0172045. [Google Scholar]
- Li, Y.; Fang, K.; Cao, C.; Li, D.; Zhou, F.; Dong, Z.; Zhang, Y.; Gan, Z. A tree-ring chronology spanning 210 years in the coastal area of southeastern China, and its relationship with climate change. Clim. Res. 2016, 67, 209–220. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Stokes, M.A. An Introduction to Tree-Ring Dating; University of Arizona Press: Arizona, AZ, USA, 1996. [Google Scholar]
- Schweingruber, F.H. Tree Rings: Basics and Applications of Dendrochronology; Springer Science & Business Media: New York, NY, USA, 1988. [Google Scholar]
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree-Ring Bull. 1983, 43, 52–67. [Google Scholar]
- Cook, E.R. A Time Series Analysis Approach to Tree Ring Standardization. Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 1985. [Google Scholar]
- Wigley, T.M.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Buras, A. A comment on the expressed population signal. Dendrochronologia 2017, 44, 130–132. [Google Scholar] [CrossRef]
- Larsson, L.-Å. CooRecorder and CDendro Programs of the CooRecorder/CDendro Package Version 9.3. Available online: http://www.cybis.se/forfun/dendro/ (accessed on 3 March 2018).
- Rydval, M.; Larsson, L.-Å.; McGlynn, L.; Gunnarson, B.E.; Loader, N.J.; Young, G.H.; Wilson, R. Blue intensity for dendroclimatology: Should we have the blues? Experiments from Scotland. Dendrochronologia 2014, 32, 191–204. [Google Scholar] [CrossRef]
- Denne, M. Definition of latewood according to Mork (1928). Iawa J. 1989, 10, 59–62. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- Robert, H.; Steven, P.; John, L.; Jane, E. Dismo: Species Distribution Modeling. R Package Version 2020. Available online: https://cran.r-project.org/web/packages/dismo/dismo.pdf (accessed on 10 August 2022).
- Zhang, Y.; Fang, K.; ZHou, F.; Dong, Z.; Gan, Z.; Li, D. A Study on the Inter-annual Latewood Growth of pinus massoniana in Fuzhou. J. Subtrop. Resour. Environ. 2016, 11, 59–64. [Google Scholar]
- De Luis, M.; Novak, K.; Raventós, J.; Gričar, J.; Prislan, P.; Čufar, K. Climate factors promoting intra-annual density fluctuations in Aleppo pine (Pinus halepensis) from semiarid sites. Dendrochronologia 2011, 29, 163–169. [Google Scholar] [CrossRef]
- Vieira, J.; Rossi, S.; Campelo, F.; Freitas, H.; Nabais, C. Xylogenesis of Pinus pinaster under a Mediterranean climate. Ann. For. Sci. 2014, 71, 71–80. [Google Scholar] [CrossRef]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Fonti, P.; Rigling, A. Drought-induced adaptation of the xylem in Scots pine and pubescent oak. Tree Physiol. 2009, 29, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Domec, J.C.; Gartner, B.L. How do water transport and water storage differ in coniferous earlywood and latewood? J. Exp. Bot. 2002, 53, 2369–2379. [Google Scholar] [CrossRef]
- Martinez-Meier, A.; Sanchez, L.; Pastorino, M.; Gallo, L.; Rozenberg, P. What is hot in tree rings? The wood density of surviving Douglas-firs to the 2003 drought and heat wave. For. Ecol. Manag. 2008, 256, 837–843. [Google Scholar] [CrossRef]
- Gindl, W.; Grabner, M.; Wimmer, R. The influence of temperature on latewood lignin content in treeline Norway spruce compared with maximum density and ring width. Trees 2000, 14, 409–414. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B. Xylogenesis: Coniferous Trees of Temperate Forests Are Listening to the Climate Tale during the Growing Season But Only Remember the Last Words! Plant Physiol. 2016, 171, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Demura, T.; Ye, Z.-H. Regulation of plant biomass production. Curr. Opin. Plant Biol. 2010, 13, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Cuny, H.E.; Rathgeber, C.B.; Frank, D.; Fonti, P.; Mäkinen, H.; Prislan, P.; Rossi, S.; Del Castillo, E.M.; Campelo, F.; Vavrčík, H. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 2015, 1, 15160. [Google Scholar] [CrossRef]
- Larson, P.R. Some indirect effects of environment on wood formation. In The Formation of Wood in Forest Tress; Zimmermann, M.H., Ed.; Academic Press: New York, NY, USA, 1964; pp. 345–365. [Google Scholar]
- Yang, Q.; Xu, M.; Chi, Y.; Zheng, Y. Relationship between stem CO2 efflux and stem temperature at different measuring depths in Pinus massoniana trees. Acta Ecol. Sin. 2016, 36, 229–235. [Google Scholar] [CrossRef]
- Yokota, T.; Hagihara, A. Seasonal change in the temperature coefficient Q 10 for respiration of field-grown hinoki cypress (Chamaecyparis obtusa) trees. J. For. Res. 1996, 1, 165–168. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, M.; Xu, X.; Sun, Z.; Yin, G.; Piao, S. Diurnal and seasonal change in stem respiration of Larix principis-rupprechtii trees, northern China. PLoS ONE 2014, 9, e89294. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.B.; Chen, A.; Chevallier, F.; Dolman, A.J.; Janssens, I.A.; Penuelas, J.; Zhang, G. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 2013, 501, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.F.; Mulvey, R.L.; Brownlee, A.H.; Barrett, T.M.; Pattison, R.R. Warm summer nights and the growth decline of shore pine in Southeast Alaska. Environ. Res. Lett. 2015, 10, 124007. [Google Scholar] [CrossRef]
- Holtta, T.; Makinen, H.; Nojd, P.; Makela, A.; Nikinmaa, E. A physiological model of softwood cambial growth. Tree Physiol. 2010, 30, 1235–1252. [Google Scholar] [CrossRef]
- Rahman, M.H.; Begum, S.; Nakaba, S.; Yamagishi, Y.; Kudo, K.; Nabeshima, E.; Nugroho, W.D.; Oribe, Y.; Funada, R. Relationship between the earlywood-to-latewood transition and changes in levels of stored starch around the cambium in locally heated stems of the evergreen conifer Chamaecyparis pisifera. Trees-Struct. Funct. 2016, 30, 1619–1631. [Google Scholar] [CrossRef]
- Cao, X.; Fang, K.; Chen, P.; Zhang, P.; Björklund, J.; Pumijumnong, N.; Guo, Z. Microdensitometric records from humid subtropical China show distinct climate signals in earlywood and latewood. Dendrochronologia 2020, 64, 125764. [Google Scholar] [CrossRef]
- Cao, X.; Hu, H.; Kao, P.-k.; Buckley, B.M.; Dong, Z.; Chen, X.; Zhou, F.; Fang, K. Improved spring temperature reconstruction using earlywood blue intensity in southeastern China. Int. J. Climatol. 2022, 1–17. [Google Scholar] [CrossRef]
- Schweingruber, F.; Fritts, H.; Bräker, O.; Drew, L.; Schär, E. The X-ray Technique as Applied to Dendroclimatology. Tree-Ring Bull. 1978, 38, 61–91. [Google Scholar]
- Rathgeber, C. Conifer tree-ring density inter-annual variability—Anatomical, physiological and environmental determinants. New Phytol. 2017, 216, 621–625. [Google Scholar] [CrossRef]
- Dunisch. Low night temperatures cause reduced tracheid expansion in podocarpus latifolius. Iawa J. 2010, 31, 245–255. [Google Scholar] [CrossRef]
- Antonova, G.F.; Stasova, V.V. Effects of environmental factors on wood formation in larch ( Larix sibirica Ldb.) stems. Trees 1997, 11, 462–468. [Google Scholar] [CrossRef]
- Song, X.; Lyu, S.; Wen, X. Limitation of soil moisture on the response of transpiration to vapor pressure deficit in a subtropical coniferous plantation subjected to seasonal drought. J. Hydrol. 2020, 591, 125301. [Google Scholar] [CrossRef]
- Larysch, E.; Stangler, D.F.; Puhlmann, H.; Rathgeber, C.B.K.; Seifert, T.; Kahle, H.P. The 2018 hot drought pushed conifer wood formation to the limit of its plasticity: Consequences for woody biomass production and tree ring structure. Plant Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Oberhuber, W.; Gruber, A. Climatic influences on intra-annual stem radial increment of Pinus sylvestris (L.) exposed to drought. Trees 2010, 24, 887–898. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhou, F.; Fonti, P.; Ren, P.; Li, X.; Miao, G.; Dong, Z.; Fang, K. Intra-Annual Wood Formation of Cryptomeria fortunei and Cunninghamia lanceolata in Humid Subtropical China. Front. Ecol. Evol. 2021, 9, 733974. [Google Scholar] [CrossRef]
- Dong, Z.; Chen, D.; Du, J.; Yang, G.; Bai, M.; Zhou, F.; Zheng, Z.; Ruan, C.; Fang, K. A 241-Year Cryptomeria fortune Tree-Ring Chronology in Humid Subtropical China and Its Linkages with the Pacific Decadal Oscillation. Atmosphere 2020, 11, 247. [Google Scholar] [CrossRef]
- Bonfils, C. Extending the record of photosynthetic activity in the eastern United States into the presatellite period using surface diurnal temperature range. Geophys. Res. Lett. 2005, 32, L08405. [Google Scholar] [CrossRef]
- Cox, D.T.C.; Maclean, I.M.D.; Gardner, A.S.; Gaston, K.J. Global variation in diurnal asymmetry in temperature, cloud cover, specific humidity and precipitation and its association with leaf area index. Glob. Chang. Biol. 2020, 26, 7099–7111. [Google Scholar] [CrossRef]
- Lindvall, J.; Svensson, G. The diurnal temperature range in the CMIP5 models. Clim. Dyn. 2015, 44, 405–421. [Google Scholar] [CrossRef]
- Dai, A.; Trenberth, K.E.; Karl, T.R. Effects of clouds, soil moisture, precipitation, and water vapor on diurnal temperature range. J. Clim. 1999, 12, 2451–2473. [Google Scholar] [CrossRef]
- Gimeno, T.E.; Camarero, J.J.; Granda, E.; Pías, B.; Valladares, F. Enhanced growth of Juniperus thurifera under a warmer climate is explained by a positive carbon gain under cold and drought. Tree Physiol. 2012, 32, 326–336. [Google Scholar] [CrossRef]
- Tao, Q.; Zhang, Q.B.; Chen, X. Tree-ring reconstructed diurnal temperature range on the eastern Tibetan plateau and its linkage to El Niño-Southern Oscillation. Int. J. Climatol. 2020, 41, 1696–1711. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, M.; Wang, X.; Gu, F.; Liu, S. Divergent tree growth response to recent climate warming of Abies faxoniana at alpine treelines in east edge of Tibetan Plateau. Ecol. Res. 2018, 33, 303–311. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y.; Qian, H.; Liu, R. Inverse effects of recent warming on trees growing at the low and high altitudes of the Dabie Mountains, subtropical China. Dendrochronologia 2020, 59, 125649. [Google Scholar] [CrossRef]
- Rakthai, S.; Fu, P.L.; Fan, Z.X.; Gaire, N.P.; Pumijumnong, N.; Eiadthong, W.; Tangmitcharoen, S. Increased Drought Sensitivity Results in a Declining Tree Growth of Pinus latteri in Northeastern Thailand. Forests 2020, 11, 361. [Google Scholar] [CrossRef]
- Hza, E.; Qcab, C.; Yu, L. Altitudinal difference of growth-climate response models in the north subtropical forests of China. Dendrochronologia 2022, 72, 125935. [Google Scholar]
- Jacoby, G.C.; D’Arrigo, R.D. Tree ring width and density evidence of climatic and potential forest change in Alaska. Glob. Biogeochem. Cycles 1995, 9, 227–234. [Google Scholar] [CrossRef]
- Cook, E.R.; Peters, K. Calculating unbiased tree-ring indices for the study of climatic and environmental change. Holocene 1997, 7, 361–370. [Google Scholar] [CrossRef]
- Barber, V.A.; Juday, G.P.; Finney, B.P. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef]
- Yonenobu, H.; Eckstein, D. Reconstruction of early spring temperature for central Japan from the tree-ring widths of Hinoki cypress and its verification by other proxy records. Geophys. Res. Lett. 2006, 33, L10701. [Google Scholar] [CrossRef]
- Esper, J.; Frank, D.; Büntgen, U.; Verstege, A.; Hantemirov, R.M.; Kirdyanov, A.V. Trends and uncertainties in Siberian indicators of 20th century warming. Glob. Chang. Biol. 2010, 16, 386–398. [Google Scholar] [CrossRef]
- Youngblut, D.K.; Luckman, B.H. Evaluating the temperature sensitivity of radial growth patterns from whitebark pine in the western Canadian Cordillera. Dendrochronologia 2013, 31, 16–28. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Z.; Fang, K.; Chen, Y.; Dong, Z.; Zhou, F.; Li, Y. Is the Pinus massoniana Lamb. Tree-Ring Latewood Formation Influenced by the Diurnal Temperature Range in Humid Subtropical China? Forests 2022, 13, 1439. https://doi.org/10.3390/f13091439
Zheng Z, Fang K, Chen Y, Dong Z, Zhou F, Li Y. Is the Pinus massoniana Lamb. Tree-Ring Latewood Formation Influenced by the Diurnal Temperature Range in Humid Subtropical China? Forests. 2022; 13(9):1439. https://doi.org/10.3390/f13091439
Chicago/Turabian StyleZheng, Zhuangpeng, Keyan Fang, Yao Chen, Zhipeng Dong, Feifei Zhou, and Yingjun Li. 2022. "Is the Pinus massoniana Lamb. Tree-Ring Latewood Formation Influenced by the Diurnal Temperature Range in Humid Subtropical China?" Forests 13, no. 9: 1439. https://doi.org/10.3390/f13091439