
Genome-Wide Identification and Expression Analyses of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Eucommia ulmoides

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Determination of Chlorogenic Acid
2.2.1. Materials Treatment
2.2.2. Reaction Conditions
2.3. Identification of the 4CL Gene Family in E. ulmoides
2.4. Bioinformatic Analysis of EU4CL Genes
2.5. RNA Extraction and Quantitative Reverse Transcription-PCR
(qRT-PCR) Assay
2.6. Analysis of 4CL Gene Expression Patterns in E. ulmoides
3. Results
3.1. Identification and Protein Physicochemical Feature Analysis of 4CL in E. ulmoides
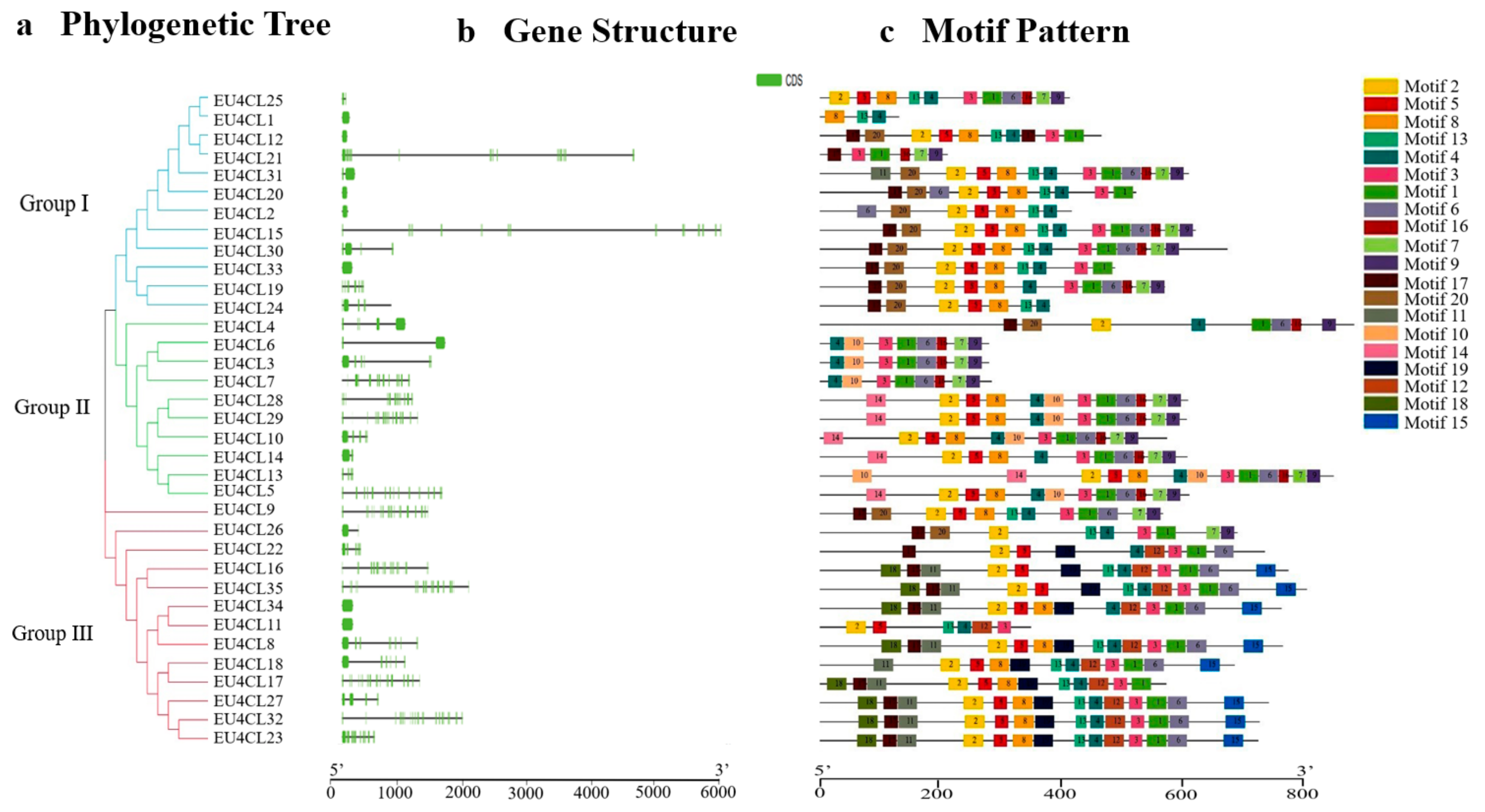
3.2. Conserved Motifs and Gene Structure Analysis
3.3. Phylogenetic Analysis of the 4CL Gene Family in E. ulmoides
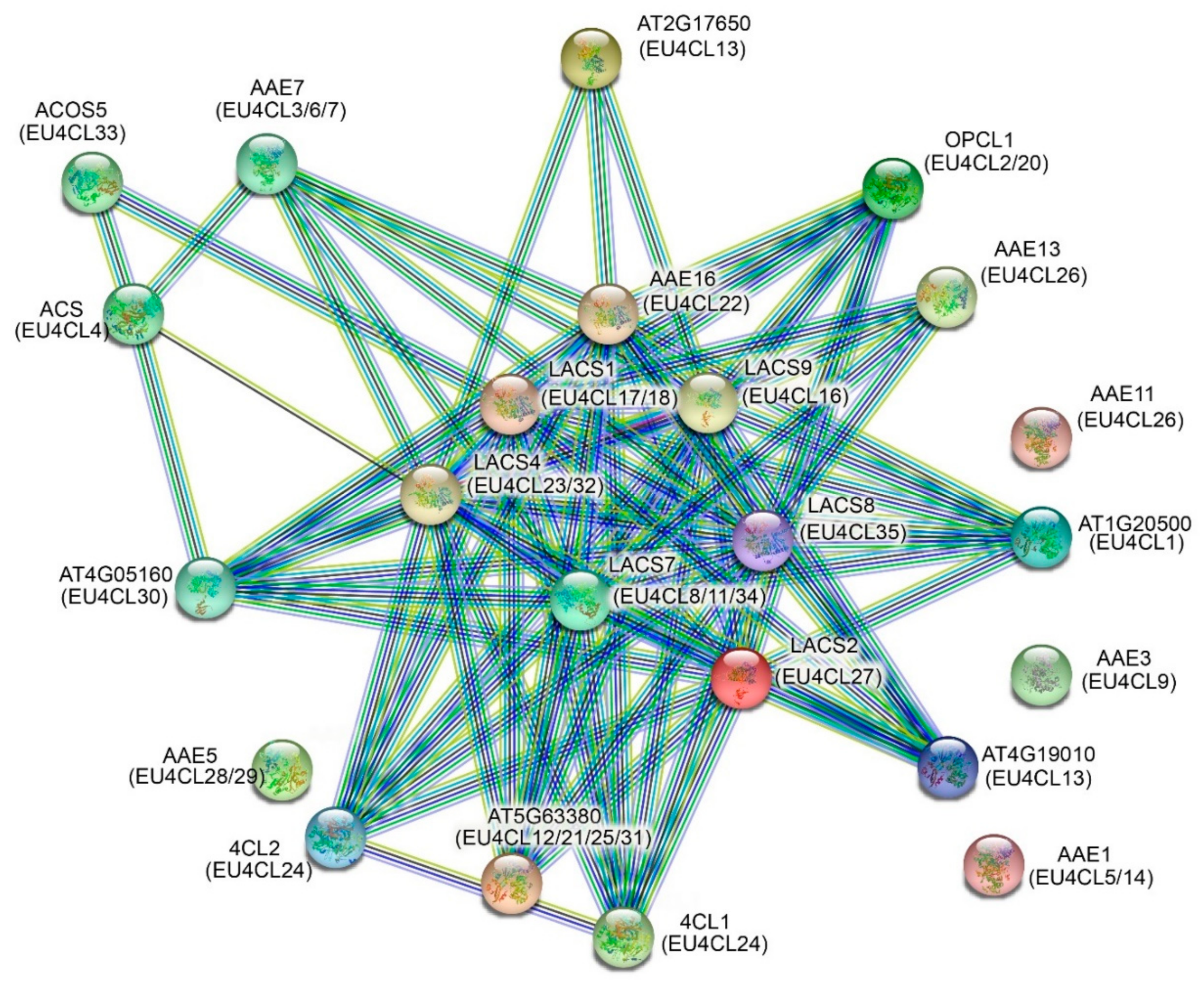
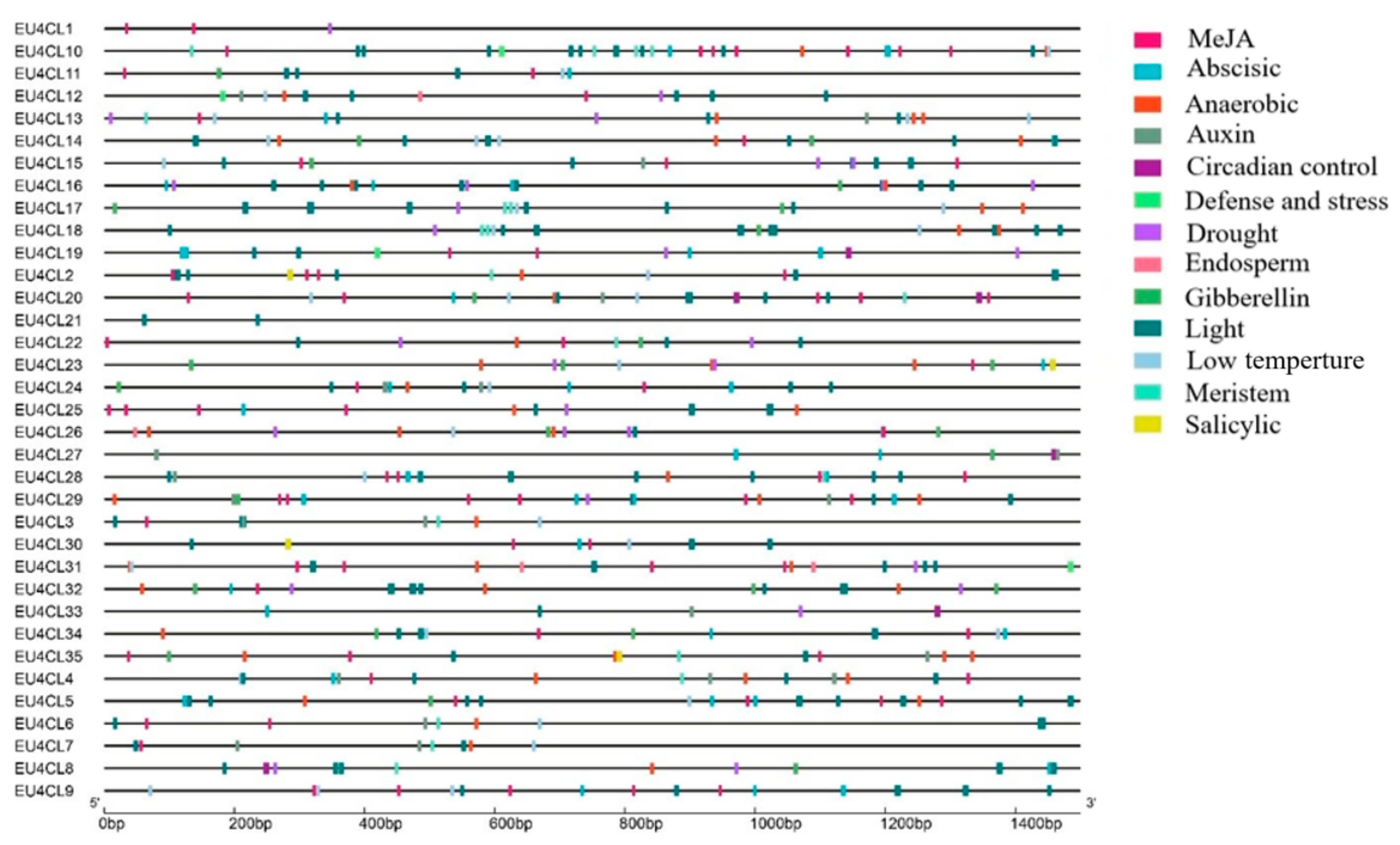
3.4. Interaction Network and Promoter Cis-Acting Element Analysis of Eu4CLs
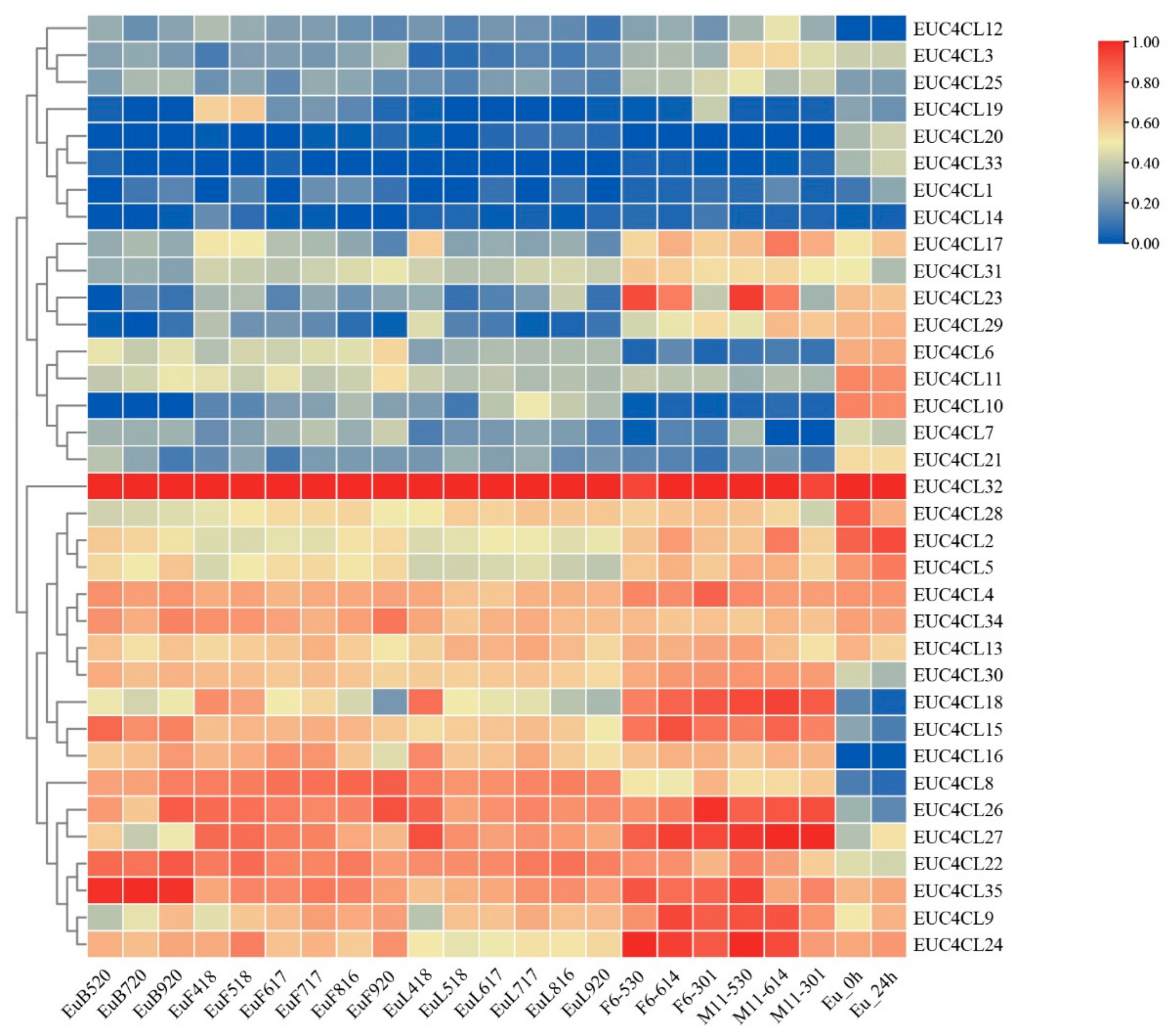
3.5. Analysis of 4CL Expression Patterns in E. ulmoides
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Du, Q.X.; Wei, Y.X.; Liu, P.F.; Du, H.Y. Diversity of the Content of Main Active Components in Eucommia ulmoides Male Flowers. Sci. Silvae Sin. 2017, 53, 35–43. [Google Scholar]
- Qing, J.; Wei, Y.X.; Du, H.Y.; Du, L.Y. Study on Genetic Variation of Main Active Components in Leaves of Eucommia ulmoides. Acta. Bot. Boreali-Occident. Sin. 2018, 38, 316–323. [Google Scholar]
- Dong, J.; Ma, X.; Fu, Z.; Guo, Y. Effects of microwave drying on the contents of functional constituents of Eucommia ulmoides flower tea. Ind. Crops Prod. 2011, 34, 1102–1110. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, K.; Ali, K. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 34, 267–283. [Google Scholar] [CrossRef]
- Li, F.M.; Chen, L.X.; Li, J.J. Identification and Bioinformatics Analysis of Gene for Chlorogenic Acid in Lonicera japonica Thunb. Mol. Plant Breed. 2021, 5, e3781. [Google Scholar]
- Kundu, A.; Vadassery, J. Chlorogenic acid-mediated chemical defence of plants against insect herbivores. Plant Biol. 2019, 21, 185–189. [Google Scholar] [CrossRef]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D. Chlorogenic Acid: Recent Advances on Its Dual Role as a Food Additive and a Nutraceutical against Metabolic Syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Xu, X.B.; Guo, H.; Zhong, C.H. Research Progress on Accumulation and Biosynthesis of Chlorogenic Acid in Lonicerae japonicae. Food Serv. 2021, 42, 252–256. [Google Scholar]
- Luo, Y.E.; Tian, J.X.; Ye, J.M.; Li, J.C. Analysis and Regulation of Biosynthetic Pathways of Chlorogenic Acid in Fungi. Sci. Technol. Eng. 2021, 21, 14029–14039. [Google Scholar]
- Endler, A.; Martens, S.; Wellmann, F.; Matern, U. Unusually divergent 4-coumarate: CoA-ligases from Ruta graveolens L. Plant Mol. Biol. 2008, 67, 335–346. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Zheng, K.; Xie, M.; Feng, K.; Jawdy, S.S. Genome-wide association studies and expression-based quantitative trait loci analyses reveal roles of HCT2 in caffeoylquinic acid biosynthesis and its regulation by defense-responsive transcription factors in Populus. New Phytol. 2018, 220, 502–516. [Google Scholar] [CrossRef] [Green Version]
- Kiewning, H. Structure and evolution of 4-coumarate: Coenzyme A ligase (4CL) gene families. Biol. Chem. 2001, 382, 645–654. [Google Scholar]
- He, L.; Yan, X.; Yuan, Y.; Zhong, J.J. Genome-wide Identification and Bioinformatics Analysis of the 4CL Gene Family in Pepper (Capsicum annuum). Mol. Plant Breed. 2022, 16, 1–15. [Google Scholar]
- Ragg, H.; Kunhn, D.N.; Hahlbrock, K. Coordinated regulation of 4-coumarate-coA ligase and phenylalanine ammonia-lyase messenger-RNAs in cultured plant-cells. J. Biol. Chem. 1981, 256, 10061–10065. [Google Scholar] [CrossRef]
- Ehlting, J.; Buttner, D.; Wang, Q.; Douglas, C.J. Three 4-coumarate: Coenzyme A ligases in Arabidopsis thaliana represent two evolutionarily divergent classes in angiosperms. Plant J. 1999, 19, 9–20. [Google Scholar] [CrossRef]
- Hu, W.J.; Kawaoka, A.; Tsai, C.J.; Lung, J.; Osakabe, K.; Ebinuma, H.; Chiang, V.L. Compartmentalized expression of two structurally and functionally distinct 4-coumarate: CoA ligase genes in aspen (Populus tremuloides). Proc. Natl. Acad. Sci. USA 1998, 95, 5407–5412. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y. Research on the formation and accumulation of main active components and nutritional regulation of Eucommia ulmoides. Chin. Acad. For. Sci. 2011, 12, 43–91. [Google Scholar]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [Green Version]
- Qing, J.; Zhong, J.; Du, H.Y.; Wang, L. Genome-wide analysis of the long noncoding RNAs and mRNAs involved in flower bud development of Eucommia ulmoides Oliver. Sci. Hortic. Amst. 2022, 300, 111038. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Hamberger, B.; Hahlbrock, K. The 4-coumarate: CoA ligase gene family in Arabidopsis thaliana comprises one rare, sinapate-activating and three commonly occurring isoenzymes. Proc. Natl. Acad. Sci. USA 2004, 101, 2209–2214. [Google Scholar] [CrossRef] [Green Version]
- Lavhale, S.G.; Kalunke, R.M.; Giri, A.P. Structural, functional and evolutionary diversity of 4-coumarate-CoA ligase in plants. Planta 2018, 248, 1063–1078. [Google Scholar] [CrossRef]
- Fang, H.; Qi, X.; Li, Y.; Yu, X.; Liu, X. De novo transcriptomic analysis of light-induced flavonoid pathway, transcription factors in the flower buds of Lonicera japonica. Trees 2020, 34, 267–283. [Google Scholar] [CrossRef]
- Guo, J.Y.; Zheng, X.Y.; Zou, C.X. Research Progress of cis-elements of Abiotic Stress Inducible Promoters and Associated Transcription Factors. Biotechnol. Bull. 2011, 4, 16–20. [Google Scholar]
- Xu, J.; Lin, Y.H.; Wang, X.N.; Han, Y.S.; Tang, L.Q.; Wang, X.H.; Zhu, H.L. Bioinformatic Identification and Expression Analysis of 4CL Genes in Ipomoea batatas. Acta Bot. Boreali-Occident. Sin. 2020, 40, 581–587. [Google Scholar]
- Harding, S.A.; Leshkevich, J.; Chiang, V.L.; Tsai, C.J. Differential substrate inhibition couples kinetically distinct 4-coumarate: Coenzyme A ligases with spatially distinct metabolic roles in quaking aspen. Plant Physiol. 2002, 128, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kim, J.I.; Pysh, L.; Chapple, C. Four isoforms of Arabidopsis thaliana 4-coumarate: CoA ligase (4CL) have overlapping yet distinct roles in phenylpropanoid metabolism. Plant Physiol. 2015, 35, 838–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.H.; Peng, X.X.; Liu, Z.; Xu, X.L. Bioinformatics Analysis on Tobacco Nt4CL Gene Family. Guizhou Agric. Sci. 2018, 46, 11–15. [Google Scholar]
- Park, J.J.; Yoo, C.G.; Flanagan, Y.Q.; Debnath, Y.X. Defined tetra-allelic gene disruption of the 4-coumarate: Coenzyme A ligase 1 (Pv4CL1) gene by CRISPR/Cas9 in switchgrass results in lignin reduction and improved sugar release. Biotechnol. Biofuels 2017, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Xiong, X.; Zhang, X.; Feng, H.; Li, Y. Characterization of the Gh4CL gene family reveals a role of Gh4CL7 in drought tolerance. BMC Plant Biol. 2020, 45, 337–346. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, T.; Wang, X.; Wang, J.H.; Gu, K.D. Genome-wide identification and expression analyses of homeodomain-leucine zipper family genes reveal their involvement in stress response in apple (Malus × domestica). Hortic. Plant J. 2022, 8, 261–278. [Google Scholar] [CrossRef]
- Zhang, T.; Li, X.M.; Zhao, Q.; Shi, Y.; Hao, Y.J. Genome-wide identification and functional characterization of the MdCLE peptide family in apple (Malus × domestica). Hortic. Plant J. 2022, 8, 279–288. [Google Scholar] [CrossRef]
- Gui, J.; Shen, J.; Li, L. Functional Characterization of Evolutionarily Divergent 4-Coumarate: Coenzyme A Ligases in Rice. Plant Physiol. 2011, 157, 574–586. [Google Scholar] [CrossRef] [Green Version]
- Soltani, B.M.; Ehlting, J.; Hamberger, B.; Douglas, C.J. Multiple cis-regulatory elements regulate distinct and complex patterns of developmental and wound-induced expression of Arabidopsis thaliana 4CL gene family members. Planta 2006, 224, 1239–1240. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Song, J.N.; Wang, J.; Lin, Y.C.; Liu, J.; Li, Q.Z. Systems Biology of Lignin Biosynthesis in Populus trichocarpa: Heteromeric 4-Coumaric Acid: Coenzyme A Ligase Protein Complex Formation, Regulation, and Numerical Modeling. Plant Cell 2014, 26, 876–893. [Google Scholar] [CrossRef] [Green Version]
- Egea, C.; Ahmed, A.S.; Candela, M.; Candela, M.E. Elicitation of peroxidase activity and lignin biosynthesis in pepper suspension cells by Phytophthora capsici. Plant Physiol. 2001, 158, 151–158. [Google Scholar] [CrossRef]
- Burbulis, I.E.; Winkel-Shirley, B. Interactions among enzymes of the Arabidopsis flavonoid biosynthetic pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 12929–12934. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Hu, W.; Jiang, A.; Xu, Y.; Yang, X. Effect of Methyl Jasmonate on Phenolic Accumulation in Wounded Broccoli. Molecules 2019, 24, 3537. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Han, Y.; Li, D.; Lin, Y.; Cai, Y. Systematic Analysis of the 4-Coumarate: Coenzyme A Ligase (4CL) Related Genes and Expression Profiling during Fruit Development in the Chinese Pear. Genes 2016, 7, 89. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Kannan, B.; Dermawan, H.; Moxley, G.W.; Altpeter, F. Precision breeding for RNAi suppression of a major 4-coumarate: Coenzyme A ligase gene improves cell wall saccharification from field grown sugarcane. Plant Mol. Biol. 2016, 92, 505–517. [Google Scholar] [CrossRef]
- Zhi, L.; Nair, S. Structural Basis for Specificity and Flexibility in a Plant 4-Coumarate: CoA Ligase. Structure 2015, 23, 2032–2042. [Google Scholar]
- Zhang, C.H.; Ma, T.; Luo, W.C.; Xu, J.M.; Liu, J.Q. Identification of 4CL Genes in Desert Poplars and Their Changes in Expression in Response to Salt Stress. Genes 2015, 6, 901–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.J.; Harding, S.A.; Tschaplinski, T.J.; Lindroth, R.L.; Yuan, Y. Genome-wide analysis of the structural genes regulating defense phenylpropanoid metabolism in Populus. New Phytol. 2010, 172, 47–62. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequences |
|---|---|
| Eu4CL4-F | TCGTTGGCGACTCTTCCATC |
| Eu4CL4-R | GCGGGATCCTCAATTGACCT |
| Eu4CL5-F | GTTCGCCTCGTCTTGTCGTA |
| Eu4CL5-R | CGTCGTGCCGGAGGTATAAT |
| Eu4CL13-F | CGCTATTACGGTTCGCTGGA |
| Eu4CL13-R | TATTCTGTGCACGCCGACTT |
| Eu4CL15-F | TCTTGCCTTTACTGTGGCTG |
| Eu4CL15-R | GCCGCAGTGTCTAGCTGAT |
| Eu4CL22-F | ATTCTTGCCGGTGGCTTGTT |
| Eu4CL22-R | GAACGGAAGGCCTCCACAAT |
| Eu4CL26-F | CTTATCTCGTCCGCTTCGCA |
| Eu4CL26-R | GTGCTAGAGGGACTGCAACA |
| Eu4CL32-F | GGCATGCAATGCTCATGGTC |
| Eu4CL32-R | AAGAAACTCCGCCGAAGCTC |
| Eu4CL34-F | AGGGGATACCGAAGGGTTCA |
| Eu4CL34-R | ACTTGACAGCATCTGGACCG |
| Eu4CL35-F | ATGGGCAGGTGTTTGATCGT |
| Eu4CL35-R | CACTCCGCTCGAGTTTCAGA |
| Actin-F | TTGTTAGCAACTGGGATGATATGG |
| Actin-R | CAGGGTGTTCTTCAGGAGCAA |
| Gene Name | Number of Amino Acids | Molecular Weight (kDa) | Theoretical pI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity | Subcellular Location |
|---|---|---|---|---|---|---|---|
| EU4CL1 | 119 | 12.61 | 6.56 | 25.02 | 91.85 | 0.246 | plasma membrane |
| EU4CL2 | 378 | 41.06 | 9.13 | 39.44 | 95.16 | 0.044 | cytoplasm |
| EU4CL3 | 254 | 27.78 | 6.33 | 42.75 | 78.27 | −0.315 | plasma membrane |
| EU4CL4 | 802 | 89.16 | 6.41 | 37.2 | 81.2 | −0.23 | cytoplasm |
| EU4CL5 | 555 | 61.04 | 6.93 | 35.41 | 88.22 | −0.033 | chloroplast |
| EU4CL6 | 254 | 28.01 | 7.02 | 43.38 | 78.27 | −0.344 | cytoplasm |
| EU4CL7 | 258 | 28.52 | 8.25 | 38 | 81.94 | −0.29 | cytoplasm |
| EU4CL8 | 695 | 76.85 | 6.89 | 29.23 | 88.78 | −0.074 | plasma membrane |
| EU4CL9 | 515 | 55.54 | 5.79 | 39.23 | 92.83 | −0.062 | plasma membrane |
| EU4CL10 | 521 | 57.76 | 6.69 | 36.28 | 89.81 | −0.139 | cytoplasm |
| EU4CL11 | 317 | 35.04 | 7.14 | 35.34 | 84.79 | −0.205 | cytoplasm |
| EU4CL12 | 423 | 45.84 | 6.32 | 47.09 | 99.15 | 0.115 | cytoplasm |
| EU4CL13 | 771 | 85.09 | 9.52 | 46.67 | 84.16 | −0.129 | plasma membrane |
| EU4CL14 | 552 | 60.61 | 6.15 | 35.05 | 96.92 | 0.084 | chloroplast |
| EU4CL15 | 564 | 61.94 | 6.17 | 36.24 | 101.44 | 0.109 | chloroplast |
| EU4CL16 | 703 | 77.19 | 6.39 | 34.65 | 92.36 | −0.08 | chloroplast |
| EU4CL17 | 520 | 58.48 | 5.35 | 34.28 | 96.15 | −0.054 | mitochondrion |
| EU4CL18 | 623 | 70.14 | 8.21 | 38.97 | 92.3 | −0.21 | plasma membrane |
| EU4CL19 | 518 | 56.67 | 6.38 | 34.99 | 103.46 | 0.078 | plasma membrane |
| EU4CL20 | 475 | 51.62 | 5.54 | 40.09 | 96.25 | 0.029 | cytoplasm |
| EU4CL21 | 192 | 21.54 | 8.77 | 42.66 | 87.34 | −0.264 | chloroplast |
| EU4CL22 | 668 | 74.74 | 8.29 | 48.55 | 96.32 | −0.183 | cytoplasm |
| EU4CL23 | 658 | 73.50 | 6.04 | 31.5 | 91.09 | −0.071 | chloroplast |
| EU4CL24 | 346 | 37.73 | 5.85 | 40.53 | 94.36 | 0.046 | cytoplasm |
| EU4CL25 | 375 | 40.42 | 8.52 | 39.11 | 91.55 | 0.023 | plasma membrane |
| EU4CL26 | 627 | 68.64 | 6.66 | 36.45 | 89.76 | −0.137 | plasma membrane |
| EU4CL27 | 674 | 75.82 | 5.79 | 38.76 | 83.04 | −0.199 | chloroplast |
| EU4CL28 | 553 | 60.72 | 8.2 | 40.53 | 88.12 | −0.06 | chloroplast |
| EU4CL29 | 551 | 60.13 | 6.4 | 36.81 | 86.46 | −0.055 | nucleus |
| EC4CL30 | 612 | 66.96 | 9.01 | 32.81 | 100.18 | 0.174 | chloroplast |
| EU4CL31 | 554 | 60.35 | 6.44 | 44.79 | 95.13 | 0.022 | chloroplast |
| EU4CL32 | 665 | 74.00 | 5.9 | 35.46 | 89.17 | −0.126 | cytoplasm |
| EU4CL33 | 443 | 48.09 | 5.31 | 27.4 | 104.04 | 0.147 | chloroplast |
| EU4CL34 | 693 | 76.44 | 6.45 | 33.66 | 91.96 | −0.038 | chloroplast |
| EU4CL35 | 731 | 79.93 | 8.26 | 30.33 | 91 | −0.068 | cytoplasm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, J.; Qing, J.; Wang, Q.; Liu, C.; Du, H.; Liu, P.; Du, Q.; Du, L.; Wang, L. Genome-Wide Identification and Expression Analyses of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Eucommia ulmoides. Forests 2022, 13, 1253. https://doi.org/10.3390/f13081253
Zhong J, Qing J, Wang Q, Liu C, Du H, Liu P, Du Q, Du L, Wang L. Genome-Wide Identification and Expression Analyses of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Eucommia ulmoides. Forests. 2022; 13(8):1253. https://doi.org/10.3390/f13081253
Chicago/Turabian StyleZhong, Jian, Jun Qing, Qi Wang, Chenlu Liu, Hongyan Du, Panfeng Liu, Qingxin Du, Lanying Du, and Lu Wang. 2022. "Genome-Wide Identification and Expression Analyses of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Eucommia ulmoides" Forests 13, no. 8: 1253. https://doi.org/10.3390/f13081253