
Influences of Shading on Ascorbic Acid Biosynthesis of Blackcurrant (Ribes nigrum L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Treatments
2.3. Measurement of Fruit Quality
2.4. Measurement of Components and Enzyme Activity in AsA Biosynthesis and Metabolism
3. Results
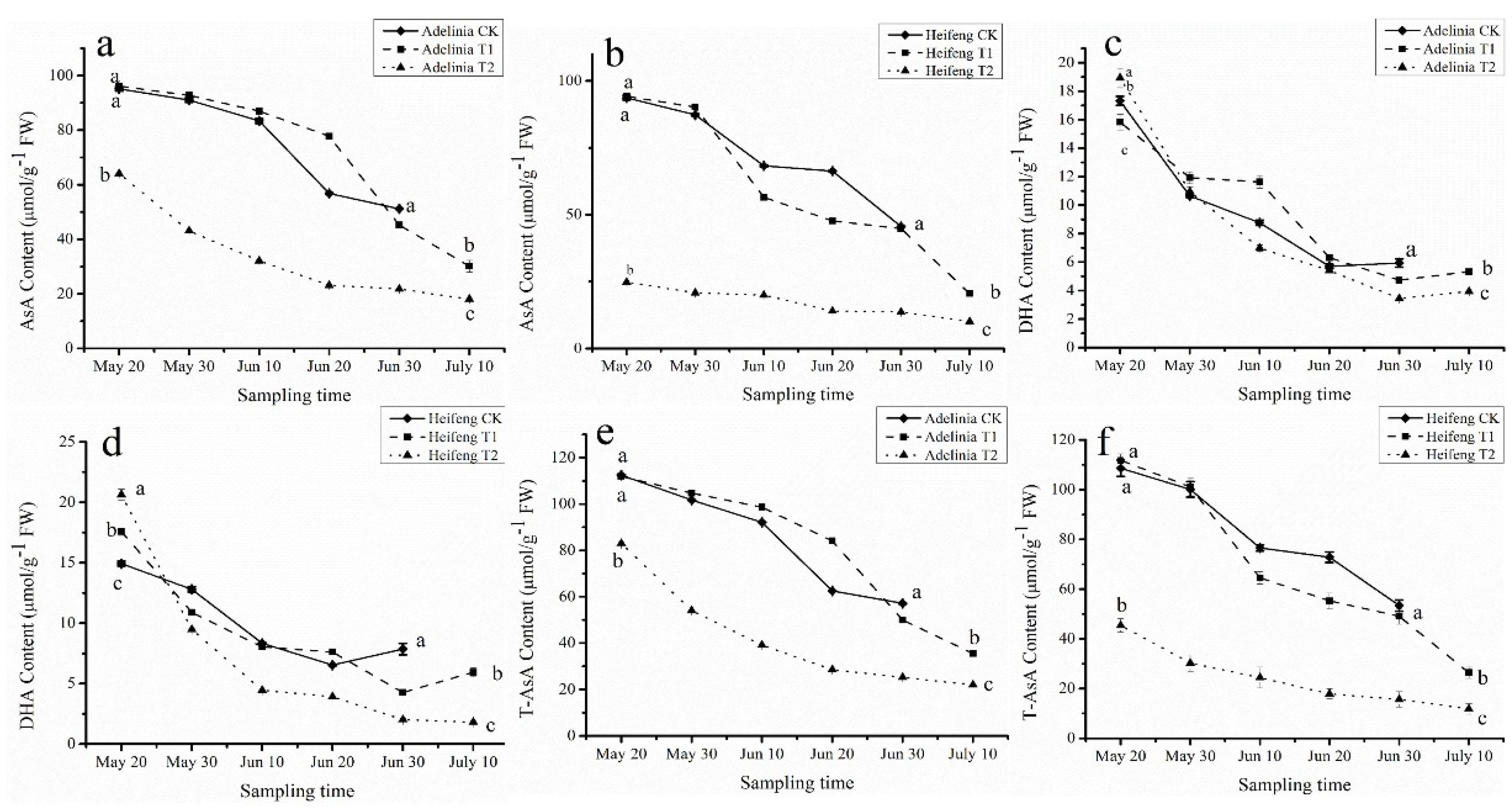
3.1. Effects of Shading on Maturity of Blackcurrant Fruits
3.2. Effects of Shading on Firmness and Soluble Solid Level in Blackcurrant Fruits
3.3. Effects of Shading on AsA, DHA and T-AsA Levels in Blackcurrant Fruits
3.4. Effects of Shading on GSH, GSSG, and T-GSH Levels in Blackcurrant Fruits
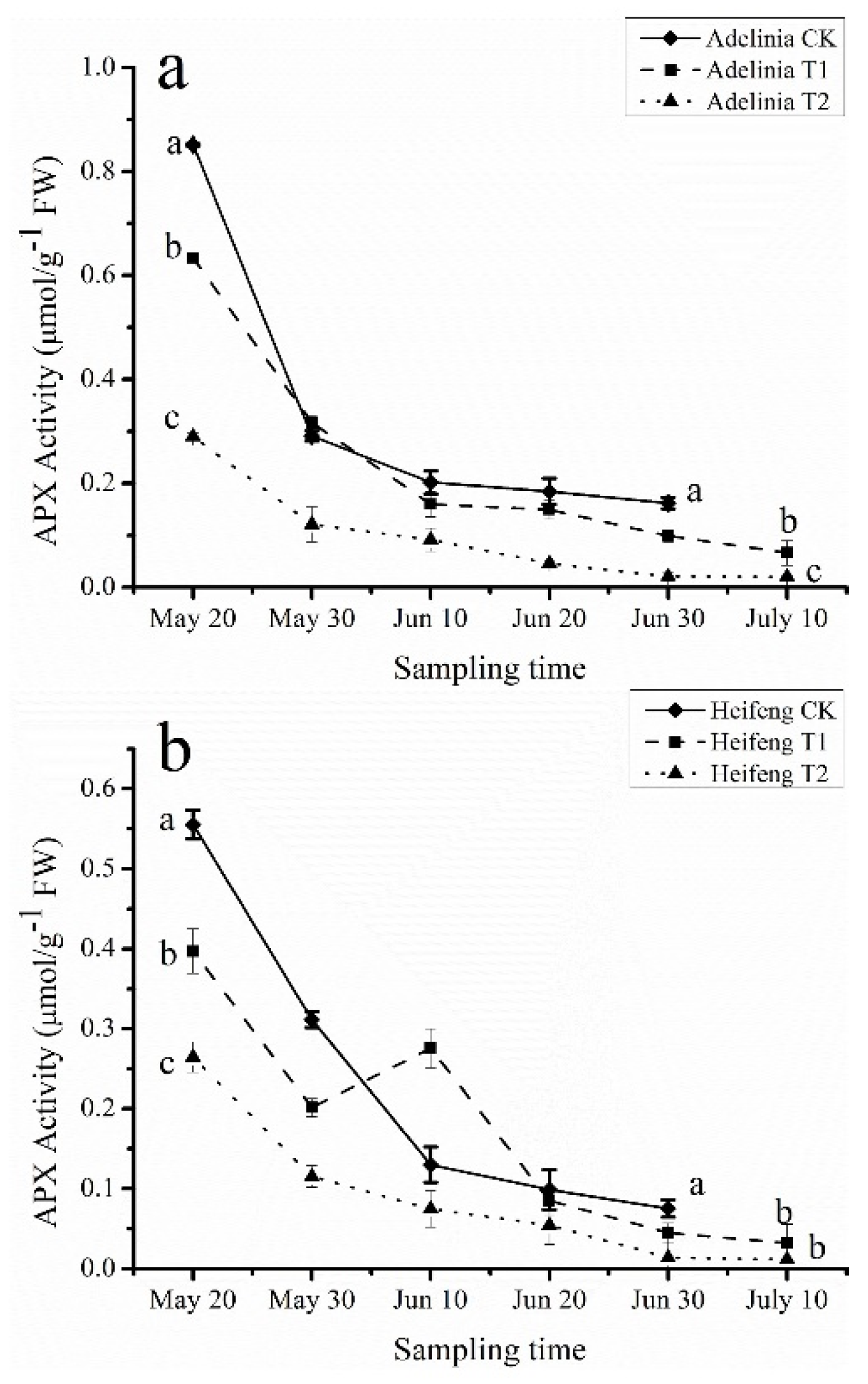
3.5. Effects of Shading on APX, DHAR, MDHAR, and GR Enzyme Activity in Blackcurrant Fruits
3.6. Effects of Shading on GalDH Activity in Blackcurrant Fruits
3.7. The Correlation between AsA Content and Enzyme Activity
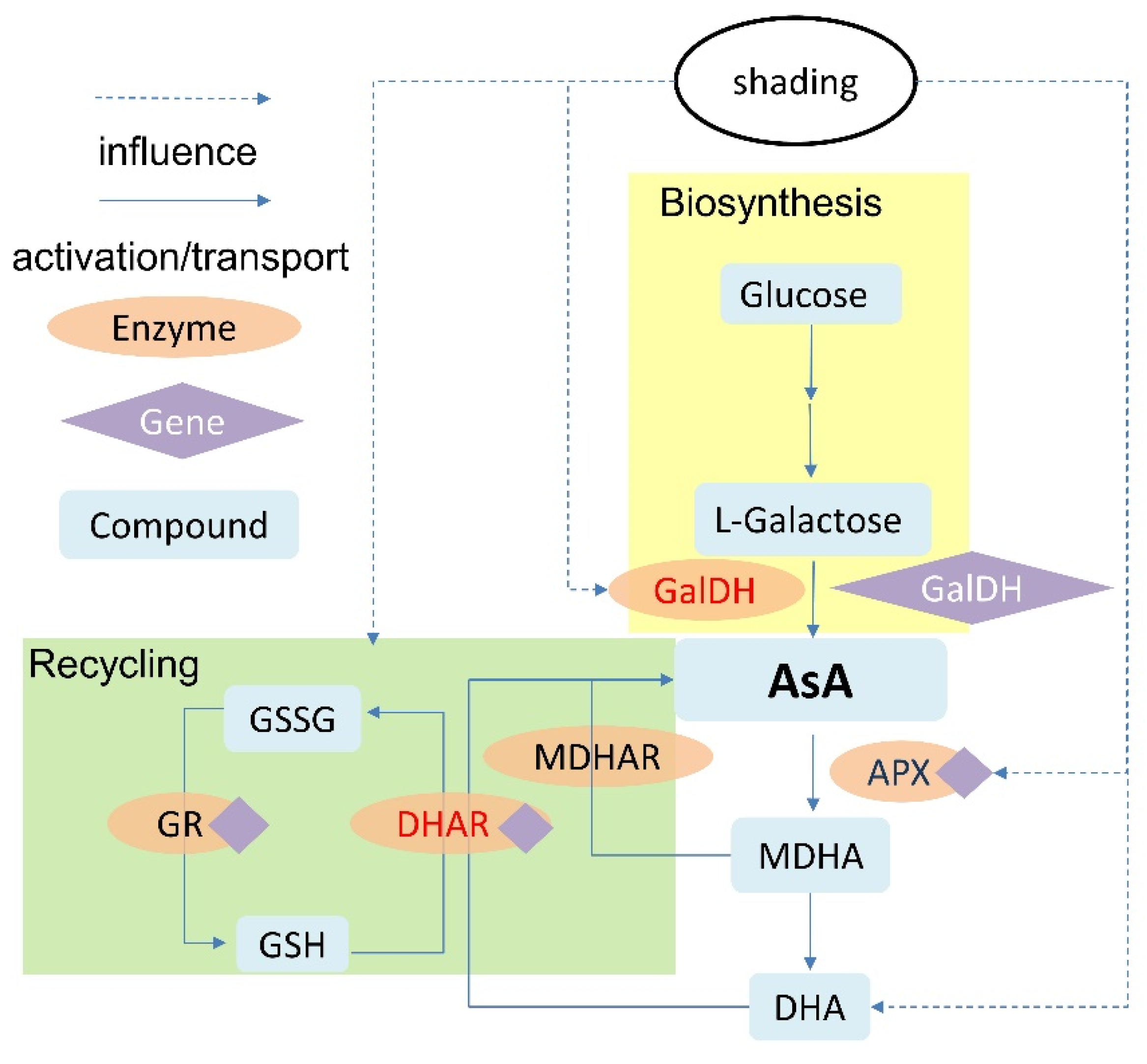
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hummer, K.E.; Dale, A. Horticulture of Ribes. For. Pathol. 2010, 40, 251–263. [Google Scholar] [CrossRef]
- Chen, Y.; Cai, X.; Zhang, Y.; Rao, X.; Fu, S. Dynamics of Understory Shrub Biomass in Six Young Plantations of Southern Subtropical China. Forests 2017, 8, 419. [Google Scholar] [CrossRef] [Green Version]
- Suring, L.H.; Goldstein, M.I.; Howell, S.; Nations, C.S. Effects of spruce beetle infestations on berry productivity on the Kenai Peninsula, Alaska. For. Ecol. Manag. 2006, 227, 247–256. [Google Scholar] [CrossRef]
- Gođevac, D.; Tesevic, V.; Vajs, V.; Milosavljevic, S.; Zdunić, G.; Đorđević, B.; Stankovic, M. Chemical Composition of Currant Seed Extracts and Their Protective Effect on Human Lymphocytes DNA. J. Food Sci. 2012, 77, C779–C783. [Google Scholar] [CrossRef]
- Schoenfeld, J.D.; Sibenaller, Z.A.; Mapuskar, K.A.; Wagner, B.A.; Cramer-Morales, K.L.; Furqan, M.; Sandhu, S.; Carlisle, T.L.; Smith, M.C.; Hejleh, T.A.; et al. O2 and H2O2-Mediated Disruption of Fe Metabolism Causes the Differential Susceptibility of NSCLC and GBM Cancer Cells to Pharmacological Ascorbate-ScienceDirect. Cancer Cell 2017, 31, 487–500. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Cytoprotective Effect of Ascorbic Acid and Rutin against Oxidative Changes in the Proteome of Skin Fibroblasts Cultured in a Three-Dimensional System. Nutrients 2020, 12, 1074. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.W.; Montagu, M.V.; Inze, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N.; Benzie, I.J.J.; Strain, J.J.; Favell, D.; Fletcher, J. Plant L-ascorbic acid: Chemistry, function, metabolism, biovailability and effects of processing. J. Sci. Food Agric. 2000, 80, 825–860. [Google Scholar] [CrossRef]
- Wheeler, G.L.; Jones, M.A.; Smirnoff, N. The biosynthetic pathway of vitamin C in higher plants. Nature 1998, 393, 365–369. [Google Scholar] [CrossRef]
- Alós, E.; Rodrigo, M.J.; Zacarías, L. Differential transcriptional regulation of l-ascorbic acid content in peel and pulp of citrus fruits during development and maturation. Planta 2014, 239, 1113–1128. [Google Scholar] [CrossRef]
- Li, M.; Ma, F.; Liang, N.; Li, J.; Wang, Y. Ascorbate Biosynthesis during Early Fruit Development Is the Main Reason for Its Accumulation in Kiwi. PLoS ONE 2010, 5, e14281. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.D.; Walker, P.G.; Pont, S.D.; Marquis, N.; Vivera, S.; Gordon, S.L.; Brennan, R.M.; Viola, R. L-Ascorbic acid accumulation in fruit of Ribes nigrum occurs by in situ biosynthesis via the L-galactose pathway. Funct. Plant Biol. 2007, 34, 1080–1091. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen Under Control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.G.; Viola, R.; Woodhead, M.; Jorgensen, L.; Gordon, S.L.; Brennan, R.M.; Hancock, R.D. Ascorbic acid content of blackcurrant fruit is influenced by both genetic and environmental factors. Funct. Plant Sci. Biotechnol. 2010, 4, 40–52. [Google Scholar]
- Sari, A.P.; Triadiati, T. Effects of shading and fertiliser on the growth and antioxidant content of olives (Olea europaea L.). Pertanika J. Trop. Agric. Sci. 2017, 40, 269–278. [Google Scholar]
- Li, M.; Ma, F.; Shang, P.; Zhang, M.; Hou, C.; Liang, D. Influence of light on ascorbate formation and metabolism in apple fruits. Planta 2009, 230, 39–51. [Google Scholar] [CrossRef]
- Ma, F.; Cheng, L. The sun-exposed peel of apple fruit has higher xanthophyll cycle-dependent thermal dissipation and antioxidants of the ascorbate–glutathione pathway than the shaded peel. Plant Sci. 2003, 165, 819–827. [Google Scholar] [CrossRef]
- Melino, V.J.; Hayes, M.A.; Soole, K.L.; Ford, C.M. The role of light in the regulation of ascorbate metabolism during berry development in the cultivated grapevine Vitis vinifera L. J. Sci. Food Agric. 2011, 91, 1712–1721. [Google Scholar] [CrossRef]
- Li, M.; Ma, F.; Liu, J.; Li, J. Shading the whole vines during young fruit development decreases ascorbate accumulation in kiwi. Physiol. Plant. 2010, 140, 225–237. [Google Scholar] [CrossRef]
- Massot, C.; Bancel, D.; Lopez Lauri, F.; Truffault, V.; Baldet, P.; Stevens, R.; Gautier, H. High Temperature Inhibits Ascorbate Recycling and Light Stimulation of the Ascorbate Pool in Tomato despite Increased Expression of Biosynthesis Genes. PLoS ONE 2013, 8, e84474. [Google Scholar]
- Massot, C.; Stevens, R.; Génard, M.; Longuenesse, J.-J.; Gautier, H. Light affects ascorbate content and ascorbate-related gene expression in tomato leaves more than in fruits. Planta 2011, 235, 153–163. [Google Scholar] [CrossRef]
- Chen, L.S.; Li, P.; Cheng, L. Effects of high temperature coupled with high light on the balance between photooxidation and photoprotection in the sun-exposed peel of apple. Planta 2008, 228, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Ilić, Z.S.; Koukounaras, A.; Milenković, L.; Kevrešan, Ž.; Bajić, A.; Šunić, L.; Kovac, R.; Fallik, E.; Mastilović, J. Grafting and Shading—The Influence on Postharvest Tomato Quality. Agriculture 2020, 10, 181. [Google Scholar] [CrossRef]
- Tiitinen, K.M.; Yang, B.; Haraldsson, G.G.; Jonsdottir, S.; Kallio, H.P. Fast Analysis of Sugars, Fruit Acids, and Vitamin C in Sea Buckthorn (Hippophaë rhamnoides L.) Varieties. J. Agric. Food Chem. 2006, 54, 2508–2513. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yang, B.; Tuomasjukka, S.; Ou, S.; Kallio, H. Effects of Latitude and Weather Conditions on Contents of Sugars, Fruit Acids, and Ascorbic Acid in Black Currant (Ribes nigrum L.) Juice. J. Agric. Food Chem. 2009, 57, 2977–2987. [Google Scholar] [CrossRef]
- Yabuta, Y.; Motoki, T.; Yoshimura, K.; Takeda, T.; Ishikawa, T.; Shigeoka, S. Thylakoid membrane-bound ascorbate peroxidase is a limiting factor of antioxidative systems under photo-oxidative stress. Plant J. 2002, 32, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Zhang Hong, G.J. Analysis of Nutritional Components of Different Pomegranate Varieties. J. Anhui Agric. Sci. 2006, 3, 460–477. [Google Scholar]
- Alvarenga, A.A.D.; Castro, E.M.D.; Lima Junior, É.D.C.; Magalhães, M.M. Effects of different light levels on the initial growth and photosynthesis of Croton urucurana Baill. in southeastern Brazil. Rev. Rvore 2003, 27, 53–57. [Google Scholar] [CrossRef]
- Han, C.J.; Wang, Q.; Zhang, H.B.; Dong, H.Z. Seed development and nutrient accumulation as affected by light shading in oilseed peony (Paeonia ostii ‘Feng Dan’). Sci. Hortic. 2019, 251, 25–31. [Google Scholar] [CrossRef]
- Qiao, X.; Sai, L.; Chen, X.; Xue, L.; Lei, J. Impact of fruit-tree shade intensity on the growth, yield, and quality of intercropped wheat corresponding. PLoS ONE 2009, 14, e0203238. [Google Scholar]
- Li, F.; Qin, D.; Li, S.; Zhan, G.; Huo, J.; Wang, H.; Yi, Z.; Wei, H.; Rengzhi, B. Ascorbic acid content and difference of related enzyme activities in fruit of blackcurrant. J. South. Agric. 2014, 52, 98–110. [Google Scholar]
- Qin Dong, H.J.; Wei, S. Breeding of Adrienaya, a new cold-resistant large fruit variety of black currant. China Fruit Tree 2012, 1, 10–12. [Google Scholar]
- Hou, C.; Li, M.; Ma, F.; Liang, D. Changes of Product Accumulation and Related Enzyme Activities in AsA Metabolism During Kiwifruit Growth and Development. Acta Hortic. Sin. 2009, 36, 1269–1276. [Google Scholar]
- Bingxiu, Z.; Zhugang, L.; Yuan, G.; Dan, L.; Qingyu, G. Relationship between DHAR and ASA accumulation in strawberry and genetic transformation of DHAR RNAi. J. South. Agric. 2012, 43, 7. [Google Scholar]
- Xia, H.; Lin, L.; Gao, F.; Ni, Z.; Gao, L.; Liang, D. Changes of AsA Content and Related Enzyme Activities in Sweet Cherry ’Satonishiki’ Fruit during Development. Acta Bot. Boreali-Occident. Sin. 2016, 36, 7. [Google Scholar]
- Hossain, M.A.; Munné-Bosch, S.; Burritt, D.J.; Diaz-Vivancos, P.; Fujita, M.; Lorence, A. Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Springer International Publishing: Cham, Switzerland, 2017; pp. 177–200. [Google Scholar]
- Viviani, A.; Fambrini, M.; Giordani, T.; Pugliesi, C. L-Ascorbic acid in plants: From biosynthesis to its role in plant development and stress response. Agrochim.-Pisa 2021, 65, 151–170. [Google Scholar]
- Broad, R.C.; Bonneau, J.P.; Hellens, R.P.; Johnson, A.A. Manipulation of Ascorbate Biosynthetic, Recycling, and Regulatory Pathways for Improved Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2020, 21, 1790. [Google Scholar] [CrossRef] [Green Version]
- Hao, C.; Fan, R.; Zhang, X.; Wang, L.; Chen, W.; Chen, Z. Physiological response of Monimopetalum chinense to light stress under habitat fragmentation. Plant Soil Environ. 2010, 56, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Yabuta, Y.; Mieda, T.; Rapolu, M.; Nakamura, A.; Motoki, T.; Maruta, T.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. Light regulation of ascorbate biosynthesis is dependent on the photosynthetic electron transport chain but independent of sugars in Arabidopsis. J. Exp. Bot. 2007, 58, 2661–2671. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, V.R.; Tarlyn, N.M. l-Ascorbic Acid Is Accumulated in Source Leaf Phloem and Transported to Sink Tissues in Plants. Plant Physiol. 2002, 130, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Tedone, L.; Hancock, R.D.; Alberino, S.; Haupt, S.; Viola, R. Long-distance transport of L-ascorbic acid in potato. BMC Plant Biol. 2004, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Imai, T.; Ban, Y.; Terakami, S.; Yamamoto, T.; Moriguchi, T. l-Ascorbate biosynthesis in peach: Cloning of six l-galactose pathway-related genes and their expression during peach fruit development. Physiol. Plant. 2009, 136, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, X.; Wang, P.; Ma, F. Ascorbic Acid Accumulation and Expression of Genes Involved in Its Biosynthesis and Recycling in Developing Apple Fruit. J. Am. Soc. Hortic. Sci. 2011, 136, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Aragüez, I.; Cruz-Rus, E.; Botella, M.; Medina-Escobar, N.; Valpuesta, V. Proteomic analysis of strawberry achenes reveals active synthesis and recycling of l-ascorbic acid. J. Proteom. 2013, 83, 160–179. [Google Scholar] [CrossRef] [PubMed]
- Mieda, T.; Yabuta, Y.; Rapolu, M.; Motoki, T.; Takeda, T.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. Feedback Inhibition of Spinach l-Galactose Dehydrogenase by l-Ascorbate. Plant Cell Physiol. 2004, 45, 1271–1279. [Google Scholar] [CrossRef]
- Chen, Z.; Young, T.E.; Ling, J.; Chang, S.C.; Gallie, D.R. Increasing vitamin C content of plants through enhanced ascorbate recycling. Proc. Natl. Acad. Sci. USA 2003, 100, 3525–3530. [Google Scholar] [CrossRef] [Green Version]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Shibahara, T.; Inanaga, S.; Tanaka, K. Overexpression of monodehydroascorbate reductase in transgenic tobacco confers enhanced tolerance to ozone, salt and polyethylene glycol stresses. Planta 2007, 225, 1255–1264. [Google Scholar] [CrossRef]
- Qin, A.; Shi, Q.; Yu, X. Ascorbic acid contents in transgenic potato plants overexpressing two dehydroascorbate reductase genes. Mol. Biol. Rep. 2010, 38, 1557–1566. [Google Scholar] [CrossRef]
- Han, W. Cloning and Quantitative Expression Analysis of Ascorbic Acid Synthase-Related Genes in Actinidia Chinensis. Master Thesis, Jiangxi Agricultural University, Nanchang, China, 2015. [Google Scholar]
- Bartoli, C.G.; Yu, J.; Gomez, F.; Fernández, L.; McIntosh, L.; Foyer, C.H. Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J. Exp. Bot. 2006, 57, 1621–1631. [Google Scholar] [CrossRef]
- Bingliang, Z.; Min, X.; Qionqiu, Q.; Qinghua, S.; Jiashu, C. Effect of shading on curd growth and antioxidant system in early cauliflower. J. Zhejiang Univ. Agric. Life Sci. Ed. 2005, 31, 535–540. [Google Scholar]
- Cangi, R.; Yagcı, A.; Akgul, S.; Kesgin, M.; Yanar, Y. Effects of shading and covering material application for delaying harvest on gray mold disease severity. Afr. J. Biotechnol. 2011, 10, 12182–12187. [Google Scholar]
- Nishikawa, F.; Kato, M.; Hyodo, H.; Ikoma, Y.; Sugiura, M.; Yano, M. Effect of sucrose on ascorbate level and expression of genes involved in the ascorbate biosynthesis and recycling pathway in harvested broccoli florets. J. Exp. Bot. 2005, 409, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, F.; Li, M.; Ma, F.; Cheng, L. Effects of location within the tree canopy on carbohydrates, organic acids, amino acids and phenolic compounds in the fruit peel and flesh from three apple (Malus × Domestica) cultivars. Hortic. Res. 2014, 1, 14019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergqvist, J.; Dokoozlian, N.; Ebisuda, N. Sunlight Exposure and Temperature Effects on Berry Growth and Composition of Cabernet Sauvignon and Grenache in the Central San Joaquin Valley of California. Am. J. Enol. Vitic. 2001, 52, 1–7. [Google Scholar]
- Šavikin, K.; Mikulič-Petkovšek, M.; Djordjević, B.; Zdunić, G.; Janković, T.; Djurović, D.; Veberič, R. Influence of shading net on polyphenol profile and radical scavenging activity in different varieties of black currant berries. Sci. Hortic. 2013, 160, 20–28. [Google Scholar] [CrossRef]
- Pastenes, C.; Santa-María, E.; Infante, R.; Franck, N. Domestication of the Chilean guava (Ugni molinae Turcz.), a forest understorey shrub, must consider light intensity. Sci. Hortic. 2003, 98, 71–84. [Google Scholar] [CrossRef]
- Di Silvestro, R.; Di Loreto, A.; Bosi, S.; Bregola, V.; Marotti, I.; Benedettelli, S.; Segura-Carretero, A.; Dinelli, G. Environment and genotype effects on antioxidant properties of organically-grown wheat varieties. A 3-year study. J. Sci. Food Agric. 2016, 97, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Franck, N.; Winkler, S.; Pastenes, C.; Infante, R. Acclimation to sun and shade of three accessions of the Chilean native berry-crop murta. Agrofor. Syst. 2007, 69, 215–229. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Treatment | Firmness (kg/cm2 × (105 Pa)) | Soluble Solid (%) |
|---|---|---|---|
| Adelinia | CK | 2.44 ± 0.36 c | 18.9 ± 0.46 a |
| T1 | 4.81 ± 0.45 b | 17.8 ± 0.15 b | |
| T2 | 5.77 ± 0.29 a | 17.5 ± 0.33 b | |
| Heifeng | CK | 2.93 ± 0.59 c | 17.4 ± 0.12 a |
| T1 | 4.99 ± 0.32 b | 17.3 ± 0.26 a | |
| T2 | 6.48 ± 0.16 a | 17.4 ± 0.19 a |
| Cultivars | MDHAR μmoL/g·FW | DHAR μmoL/g·FW | GR μmoL/g·FW | APX μmoL/g·FW | GalDH μmoL/g·FW | |
|---|---|---|---|---|---|---|
| AsA μmoL/g·FW | Adelinia CK | 0.772 | 0.928 * | 0.965 ** | 0.657 | 0.590 |
| Adelinia T1 | 0.833 * | 0.983 ** | 0.962 ** | 0.703 | 0.559 | |
| Adelinia T2 | 0.969 ** | 0.987 ** | 0.995 ** | 0.985 ** | 0.990 ** | |
| Heifeng CK | 0.894 * | 0.532 | 0.938 * | 0.877 | 0.872 | |
| Heifeng T1 | 0.950 ** | 0.856 * | 0.816 * | 0.813 * | 0.922 ** | |
| Heifeng T2 | 0.942 ** | 0.941 ** | 0.925 ** | 0.891 * | 0.963 ** | |
| GSH μmoL/g·FW | Adelinia CK | −0.950 * | −0.606 | −0.847 | −0.820 | −0.817 |
| Adelinia T1 | −0.894 * | −0.847 * | −0.759 | −0.596 | −0.541 | |
| Adelinia T2 | −0.641 | −0.797 | −0.785 | −0.726 | −0.770 | |
| Heifeng CK | −0.673 | −0.236 | −0.845 | −0.758 | −0.913 * | |
| Heifeng T1 | −0.883 * | −0.831 * | −0.782 | −0.879 * | −0.994 ** | |
| Heifeng T2 | −0.899 * | −0.906 * | −0.869 * | −0.846 * | −0.906 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gang, H.; Zhang, D.; Sun, X.; Huo, J.; Qin, D. Influences of Shading on Ascorbic Acid Biosynthesis of Blackcurrant (Ribes nigrum L.). Forests 2022, 13, 1127. https://doi.org/10.3390/f13071127
Gang H, Zhang D, Sun X, Huo J, Qin D. Influences of Shading on Ascorbic Acid Biosynthesis of Blackcurrant (Ribes nigrum L.). Forests. 2022; 13(7):1127. https://doi.org/10.3390/f13071127
Chicago/Turabian StyleGang, Huixin, Danni Zhang, Xiaojuan Sun, Junwei Huo, and Dong Qin. 2022. "Influences of Shading on Ascorbic Acid Biosynthesis of Blackcurrant (Ribes nigrum L.)" Forests 13, no. 7: 1127. https://doi.org/10.3390/f13071127