
Larval Instars and Adult Flight Period of Monochamus saltuarius (Coleoptera: Cerambycidae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Larvae Collection
2.2. Measuring Larval Instar
2.3. Trapping Adults
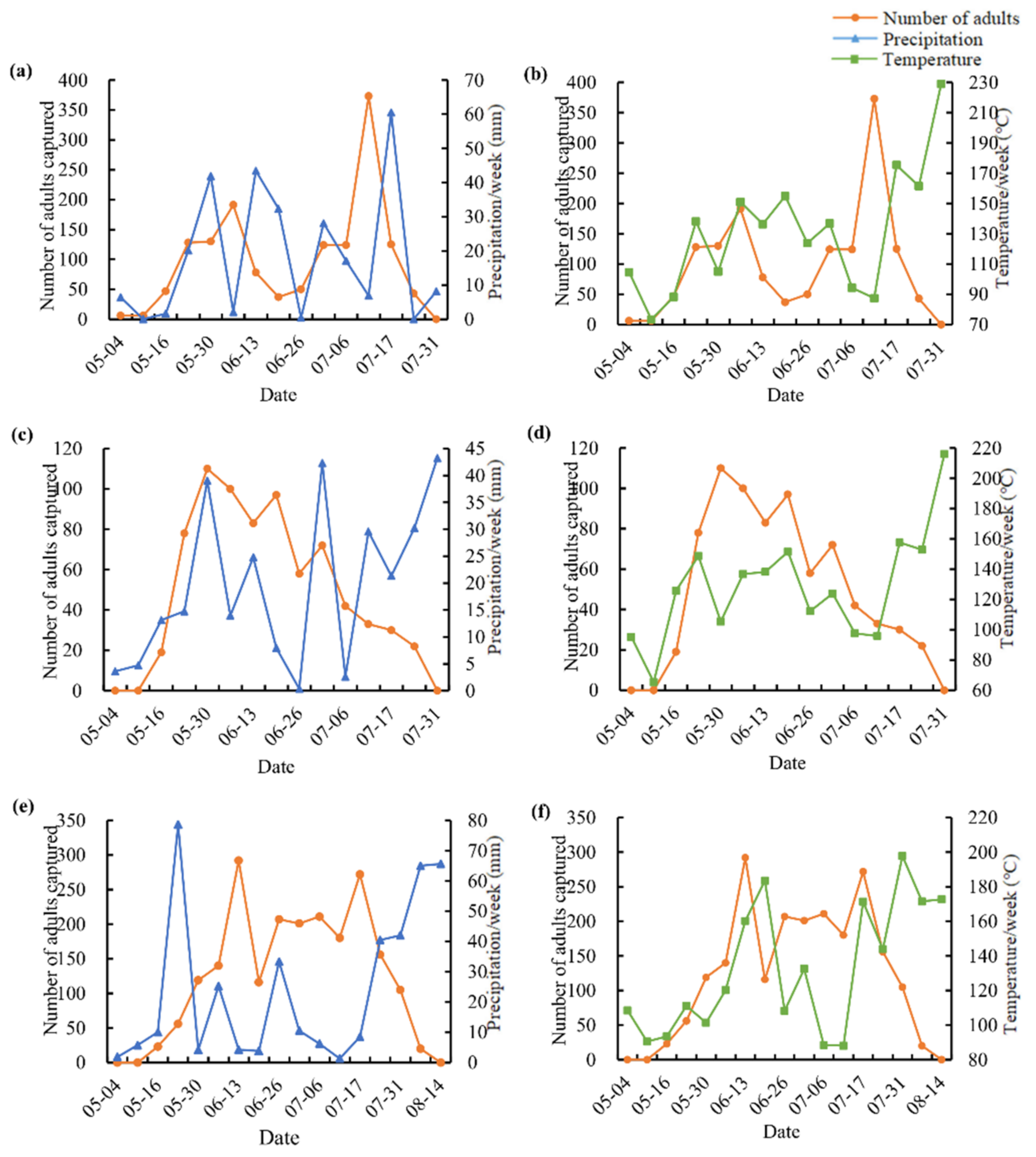
2.4. Survey of Emergence Period
2.5. Statistical Analysis
3. Results
3.1. Larval Instars
3.2. Adult Flight Period
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Z.C. Monocraphia of Original Colored Longicorn Beetles of China’s Northeast; Jilin Science and Technology Publishing House: Jilin, China, 2003; pp. 348–372. [Google Scholar]
- Cesari, M.; Marescalchi, O.; Francardi, V.; Mantovani, B. Taxonomy and phylogeny of European Monochamus species, first molecular and karyological data. J. Zool. Syst. Evol. Res. 2010, 43, 1–7. [Google Scholar] [CrossRef]
- Heisuke, S.; Takeshi, S.; Mitsunori, K. Transmission of Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle (Nematoda: Aphelenchoididae) by Monochamus saltuarius (Gebler) (Coleoptera: Cerambycidae). J. Jpn. For. Soc. 1987, 69, 492–496. [Google Scholar]
- Kwon, T.S.; Lim, J.H.; Sim, S.J.; Kwon, Y.D.; Son, S.K.; Lee, K.Y.; Kim, Y.T.; Park, J.W.; Shin, C.H.; Ryu, S.B. Distribution patterns of Monochamus alternatus and M. saltuarius (Coleoptera: Cerambycidae) in Korea. J. Korean For. Soc. 2006, 95, 543–550. [Google Scholar]
- Togashi, K.; Jikumaru, S. Horizontal transmission of Bursaphelenchus mucronatus (Nematoda: Aphelenchoididae) between Insect Vectors of Monochamus saltuarius (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1996, 60, 90–92. [Google Scholar] [CrossRef]
- Han, J.H.; Kim, H.K.; King, W.J.; Kim, G. Feeding and oviposition preference of the Sakhalin pine sawyer Monochamus saltuarius (Coleoptera: Cerambycidae) for various tree species. Entomol. Res. 2016, 46, 331–336. [Google Scholar] [CrossRef]
- Lee, H.R.; Lee, S.C.; Lee, D.H.; Choi, W.S.; Jung, C.S.; Jeon, J.H.; Kim, J.E.; Park, I.K. Identification of the aggregation-sex pheromone produced by male Monochamus saltuarius, a major insect vector of the pine wood nematode. J. Chem. Ecol. 2017, 43, 670–678. [Google Scholar] [CrossRef]
- Lee, K.S. First record of the family Bothrideridae (Coleoptera) in Korea represented by the wood-boring beetle ectoparasite, Dastarcus helophoroides. J. Asia-Pac. Entomol. 2012, 15, 273–275. [Google Scholar]
- Cho, W.S.; Koo, H.N.; Yun, S.H.; Seon, L.J.; Hun, J.D.; Jin, K.W.; Lee, S.J.; Kyung, K.H.; Han, I.H.; Kwon, Y.D. Electron beam-induced sterility and inhibition of ovarian development in the sakhalin pine longicorn, Monochamus saltuarius (Coleoptera: Cerambycidae). J. Econ. Entomol. 2018, 111, 725–731. [Google Scholar] [CrossRef]
- Jung, C.S.; Koh, S.H.; Nam, Y.; Ahn, J.J.; Lee, C.Y.; Cho, W.I. A model for predicting spring emergence of Monochamus saltuarius (Coleoptera: Cerambycidae) from Korean white pine, Pinus koraiensis. J. Econ. Entomol. 2015, 108, 1830–1836. [Google Scholar] [CrossRef]
- Yu, H.Y.; Wu, H. New host plants and new vector insects for pine wood nematodes discovered in Liaoning. For. Pest Dis. 2018, 37, 61. (In Chinese) [Google Scholar]
- Fan, L.C.; Shi, Y.; Jiang, S.W.; Zheng, Y.N. Study on the species of long-horned beetles carrying Bursaphelenchus xylophilus in Liaoning. For. Res. 2021, 34, 174–181. (In Chinese) [Google Scholar]
- Zheng, Y.N.; Jiang, S.W.; Huang, Y.F.; Yang, Z.Q.; Gao, G.P.; Liu, P.X.; Shi, Y.; Fan, L.C.; Zhang, Y.L. Dastarcus helophoroides, natural enemy of Monochamus saltuarius was found in pine tree infected by Bursaphelenchus xylophilus in Liaoning, China. Chin. J. Biol. Control 2020, 36, 31. (In Chinese) [Google Scholar]
- Zheng, Y.N.; Shi, Y.; Li, Y.; Fan, L.C.; Wang, J.; Wang, W.T. Feeding preference of Monochamus saltuarius (Coleoptera: Cerambycidae) adults for four host pine trees. Acta Entomol. Sin. 2021, 64, 1478–1482. (In Chinese) [Google Scholar]
- Zheng, Y.N.; Zhang, Y.L.; Shi, Y.; Fan, L.C.; Li, Y. Preliminary study on the control efficiency of Monochamus saltuarius by Sclerodermus spp. Chin. J. Biol. Control 2022, 2, 306–311. (In Chinese) [Google Scholar]
- Zheng, Y.N.; Fan, L.C.; Shi, Y.; Chen, G.F.; Wang, J.; Jiang, S.W. A Comparative Study on Trapping Effect of Different Attractants on Monochamus saltuarius. J. Northeast For. Univ. 2021, 49, 102–104+115. (In Chinese) [Google Scholar]
- Li, M.; Dai, Y.; Wang, Y.; Wang, L.C.; Sun, S.H.; Chen, F.M. New insights into the life history of Monochamus saltuarius (Cerambycidae, Coleoptera) can enhance surveillance strategies for pine wilt disease. J. For. Res. 2021, 6, 2699–2707. [Google Scholar] [CrossRef]
- Wang, K.H.; Zhong, Q.L.; Nie, X.M. Analysis of occurrence reasons of pine wilt disease caused by Bursaphelenchus xylophilus in Fushun. Liaoning For. Sci. Technol. 2021, 3, 56–58. (In Chinese) [Google Scholar]
- Jiang, S.N. Chinese Longhorn Beetle Larva; Chongqing Publishing House: Chongqing, China, 1989. (In Chinese) [Google Scholar]
- Wang, X.Y.; Yang, Z.Q.; Tang, Y.L.; Jiang, J.; Yang, Y.L.; Gao, C. Determination of larval instar number and duration in the oak longhorn beetle Massicus raddei (Coleoptera: Cerambycidae). Acta Entomol. Sin. 2012, 55, 575–584. (In Chinese) [Google Scholar]
- Loerch, C.R.; Cameron, E.A. Determination of larval instars of the Bronze Birch Borer, Agrilus anxius (Coleoptera: Buprestidae). Ann. Entomol. Soc. Am. 1983, 76, 948–952. [Google Scholar] [CrossRef]
- Dyar, H.G. The number of molts of lepidopterous larvae. Psyche 1980, 175, 420–422. [Google Scholar] [CrossRef]
- Dai, Z.Y. Method for determining the age of insect larvae species. Chin. Bull. Entomol. 1980, 4, 185–187. (In Chinese) [Google Scholar]
- Cai, W.Z.; Pang, X.F.; Hua, B.Z.; Liang, G.W.; Song, D.L. General Entomology; China Agricultural University Press: Beijing, China, 2011. (In Chinese) [Google Scholar]
- Jaime, G.; Brenda, Y.C.; Alfredo, C.; Francisco, J.V.; Fernando, E.V. The coffee berry borer (Coleoptera: Curculionidae), How many instars are there? Ann. Entomol. Soc. Am. 2015, 108, 311–315. [Google Scholar]
- Rachael, A.S.; Whitney, S.Q. Life history of Allokermes galliformis (Hemiptera: Kermesidae) in Colorado. Ann. Entomol. Soc. Am. 2018, 111, 265–270. [Google Scholar]
- Huan, J.S.; Tang, C.S.; Song, H.F.; Chen, Q.S.; Kang, W.T.; He, X.Y. The biological characteristic of Monochamus alternatus in Xiamen City I: Life history. Entomol. J. East China 2005, 14, 118–121. (In Chinese) [Google Scholar]
- Liu, J.D.; Li, B.W.; Wang, J.Y.; Zhao, B.A.; Shu, J.P. Primary study on methods to determine instars of Monochamus alternatus larva. J. Zhejiang For. Sci. Technol. 2008, 4, 84–86. (In Chinese) [Google Scholar]
- Togashi, K. Life table for Monochamus alternatus (Coleoptera: Cerambycidae) within dead trees of Pinus thunbergii. Jpn. J. Entomol. 1990, 58, 217–230. [Google Scholar]
- Koutroumpa, F.A.; Vincent, B.; Rouxmorabito, G.; Martin, C.; Lieutier, F. Fecundity and larval development of Monochamus galloprovincialis (Coleoptera Cerambycidae) in experimental breeding. Ann. For. Sci. 2008, 65, 707. [Google Scholar] [CrossRef]
- Kozima, K.; Katagiri, K. On the larval instar and changes of its composition of Monochamus alternalus HOPE. J. Jpn. For. Soc. 2008, 46, 307–310. [Google Scholar]
- Myeong, S.G.; Soon, H.K.; Su, B.K.; Dong, S.K. The developmental characteristics for the head capsule width of Monochamus alternatus (Coleoptera: Cerambycidae) larvae and determination of the number of instars. J. Insect Sci. 2019, 19, 26. [Google Scholar]
- Hu, X.; Li, M.; Kenneth, F.R.; Luo, Q.Y.; Fu, H.J.; Wu, S.Q.; Liang, G.H.; Wang, R.; Zhang, F.P. Bacterial communities associated with the pine wilt disease vector Monochamus alternatus (Coleoptera: Cerambycidae) during different larval instars. J. Insect Sci. 2017, 17, 115. [Google Scholar] [CrossRef]
- Wen, X.S.; Liao, S.L.; Tang, Y.L.; Yang, Z.Q. Study on the efficacy of releasing Dastarcus helophoroides eggs against Monochamus alternatus in pine forests. Sci. Silvae Sin. 2017, 53, 133–138. (In Chinese) [Google Scholar]
- Lai, Y.X.; Wang, Y.H.; Wang, X.Y.; Yang, Z.Q.; Tang, Y.L.; Qin, R.H.; Zhang, Y.N.; Zhang, Y.F. A field test of controlling the pine wilt disease with the technique of isolating Monochamus alternatus on forest spot and releasing parasitoid Sclerodermus guani. Chin. J. Biol. Control 2012, 28, 460–465. (In Chinese) [Google Scholar]
- Han, J.H.; Yoon, C.M.; Shin, S.C.; Kim, G.H. Seasonal occurrence and morphological measurements of pine sawyer, Monochamus saltuarius adults (Coleoptera: Cerambycidae). J. Asia-Pac. Entomol. 2007, 10, 63–67. [Google Scholar] [CrossRef]
- Huang, W.L. Monitoring population dynamics of Monochamus alternatus and the effect of climate factors using pheromone trapping method. For. Pest Dis. 2020, 39, 19–23. (In Chinese) [Google Scholar]
- Yang, A.M. Study on the change law of population dynamics of Monochamus alternatus Hope adults in forests. J. Fujian For. Sci. Technol. 2004, 31, 62–64. (In Chinese) [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Larval Instar Characteristic | Measurement Method |
|---|---|---|
| L1 | Head capsule width | Measured the width of the head capsule of the larva |
| L2 | Mandible length | Measured the right mandible length, viewed from the back of the abdomen |
| L3 | Distance between the main ocelli | Measured the distance between the abdominal ocelli from the abdomen |
| L4 | Prothoracic plate width | Measured the width of the posterior margin of the sclerotized part of the anterior thoracolumbar plate, excluding the pterygoid flap |
| L5 | Mesothoracic spiracle length | Measured the right valve of the midrib, as viewed from the back |
| L6 | Body width | Measured the maximum width of the somite |
| L7 | Body length | Measured the length from the mouth to the end of the abdomen |
| Variable | Instar | Number of Measured Larvae | Mean ± SE | Minimum (mm) | Maximum (mm) | Coefficient of Variation (CV) (%) | Growth Rate | Crosby’s Ratio (%) |
|---|---|---|---|---|---|---|---|---|
| Head capsule width (L1) | 1 | 110 | 0.82 ± 0.06 e | 0.64 | 1.00 | 18.93 | ||
| 2 | 89 | 1.26 ± 0.14 d | 1.03 | 1.50 | 15.21 | 1.38 | ||
| 3 | 296 | 1.96 ± 0.39 c | 1.52 | 2.30 | 10.96 | 1.62 | −6.34 | |
| 4 | 1663 | 3.17 ± 0.31 b | 2.31 | 3.81 | 17.21 | 1.54 | 3.21 | |
| 5 | 457 | 4.25 ± 0.31 a | 3.82 | 5.99 | 13.33 | 1.36 | 3.71 | |
| Mandible length (L2) | 1 | 110 | 0.18 ± 0.02 e | 0.15 | 0.24 | 6.55 | ||
| 2 | 89 | 0.32 ± 0.05 d | 0.24 | 0.39 | 13.92 | 1.42 | ||
| 3 | 296 | 0.51 ± 0.10 c | 0.40 | 0.57 | 14.24 | 1.36 | −4.92 | |
| 4 | 1663 | 0.79 ± 0.12 b | 0.57 | 1.05 | 14.88 | 1.55 | 14.05 | |
| 5 | 457 | 1.21 ± 0.13 a | 1.05 | 1.69 | 10.75 | 1.53 | −1.05 | |
| Distance between the main ocelli (L3) | 1 | 110 | 0.61 ± 0.05 e | 0.48 | 0.73 | 23.23 | ||
| 2 | 89 | 1.00 ± 0.11 d | 0.74 | 1.22 | 27.21 | 2.43 | ||
| 3 | 296 | 1.57 ± 0.31 c | 1.23 | 1.86 | 27.42 | 1.73 | −29.00 | |
| 4 | 1663 | 2.61 ± 0.27 b | 1.86 | 3.11 | 21.41 | 1.97 | 13.82 | |
| 5 | 457 | 3.41 ± 0.23 a | 3.11 | 5.81 | 16.07 | 1.65 | −16.04 | |
| Prothoracic plate width (L4) | 1 | 110 | 0.90 ± 0.13 e | 0.61 | 1.34 | 16.07 | ||
| 2 | 89 | 1.67 ± 0.18 d | 1.36 | 1.97 | 6.28 | 1.53 | ||
| 3 | 296 | 2.49 ± 0.47 c | 1.97 | 2.97 | 13.88 | 1.48 | 7.32 | |
| 4 | 1663 | 4.66 ± 0.60 b | 2.97 | 5.85 | 12.90 | 1.37 | −7.20 | |
| 5 | 457 | 6.28 ± 0.33 a | 5.85 | 7.65 | 9.78 | 1.50 | 9.45 | |
| Mesothoracic spiracle length (L5) | 1 | 110 | 0.03 ± 0.01 d | 0.02 | 0.07 | 11.95 | ||
| 2 | 89 | 0.10 ± 0.02 c | 0.07 | 0.12 | 10.99 | 1.49 | ||
| 3 | 296 | 0.18 ± 0.05 b | 0.12 | 0.24 | 25.23 | 1.38 | −0.68 | |
| 4 | 1663 | 0.44 ± 0.08 a | 0.24 | 0.58 | 11.95 | 1.52 | −7.09 | |
| 5 | 457 | 0.68 ± 0.08 e | 0.58 | 1.02 | 7.30 | 1.37 | −9.62 | |
| Body width (L6) | 1 | 110 | 0.92 ± 0.15 a | 0.77 | 1.49 | 16.01 | ||
| 2 | 89 | 1.85 ± 0.17 c | 1.52 | 2.12 | 13.66 | 1.49 | ||
| 3 | 296 | 2.85 ± 0.44 d | 2.12 | 3.06 | 25.32 | 1.46 | 8.37 | |
| 4 | 1663 | 4.91 ± 0.70 b | 3.06 | 6.07 | 22.99 | 1.34 | −1.76 | |
| 5 | 457 | 6.71 ± 0.46 e | 6.07 | 8.23 | 7.09 | 1.85 | −8.47 | |
| Body length (L7) | 1 | 110 | 3.00 ± 0.71 e | 2.42 | 5.64 | 6.54 | ||
| 2 | 89 | 7.68 ± 1.03 d | 5.73 | 9.26 | 10.95 | 1.30 | ||
| 3 | 296 | 12.67 ± 3.00 c | 9.28 | 16.02 | 12.82 | 1.46 | 12.82 | |
| 4 | 1663 | 23.60 ± 2.62 b | 16.04 | 28.02 | 15.63 | 1.45 | −15.42 | |
| 5 | 457 | 30.34 ± 1.74 a | 28.04 | 36.50 | 14.85 | 1.60 | 1.86 |
| Variable | Regression Equation | Correlation Coefficient | Statistical Parameters |
|---|---|---|---|
| Head capsule width (L1) | YL1 = 0.22x0.37 | 0.9994 | df = 1,4, F = 3561.03, p < 0.0001 |
| Mandible length (L2) | YL2 = 0.04x0.42 | 0.9950 | df = 1,4, F = 398.34, p < 0.0001 |
| Distance between main ocelli (L3) | YL3 =0.16x0.39 | 0.9980 | df = 1,4, F = 1010.42, p < 0.0001 |
| Prothoracic plate width (L4) | YL4 = 0.19x0.45 | 0.9990 | df = 1,4, F = 2000.11, p < 0.0001 |
| Mesothoracic spiracle length (L5) | YL5 = 0.0032x0.69 | 0.9950 | df = 1,4, F = 595.68, p < 0.0001 |
| Body width (L6) | YL6 = 0.19x0.46 | 0.9960 | df = 1,4, F = 953.31, p < 0.0001 |
| Body length (L7) | YL7 = 0.66x0.50 | 0.9950 | df = 1,4, F = 843.26, p < 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, L.; Wang, J.; Wang, W.; Zheng, Y. Larval Instars and Adult Flight Period of Monochamus saltuarius (Coleoptera: Cerambycidae). Forests 2022, 13, 910. https://doi.org/10.3390/f13060910
Fan L, Wang J, Wang W, Zheng Y. Larval Instars and Adult Flight Period of Monochamus saltuarius (Coleoptera: Cerambycidae). Forests. 2022; 13(6):910. https://doi.org/10.3390/f13060910
Chicago/Turabian StyleFan, Lichun, Jue Wang, Weitao Wang, and Yanan Zheng. 2022. "Larval Instars and Adult Flight Period of Monochamus saltuarius (Coleoptera: Cerambycidae)" Forests 13, no. 6: 910. https://doi.org/10.3390/f13060910