
Phenotypic Plasticity of Cunninghamialanceolata (Lamb.) Hook. Seedlings in Response to Varied Light Quality Treatments

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatments
2.2. Measurements of Seedling Growth and Morphological Traits
2.3. Statistical Analysis
3. Results
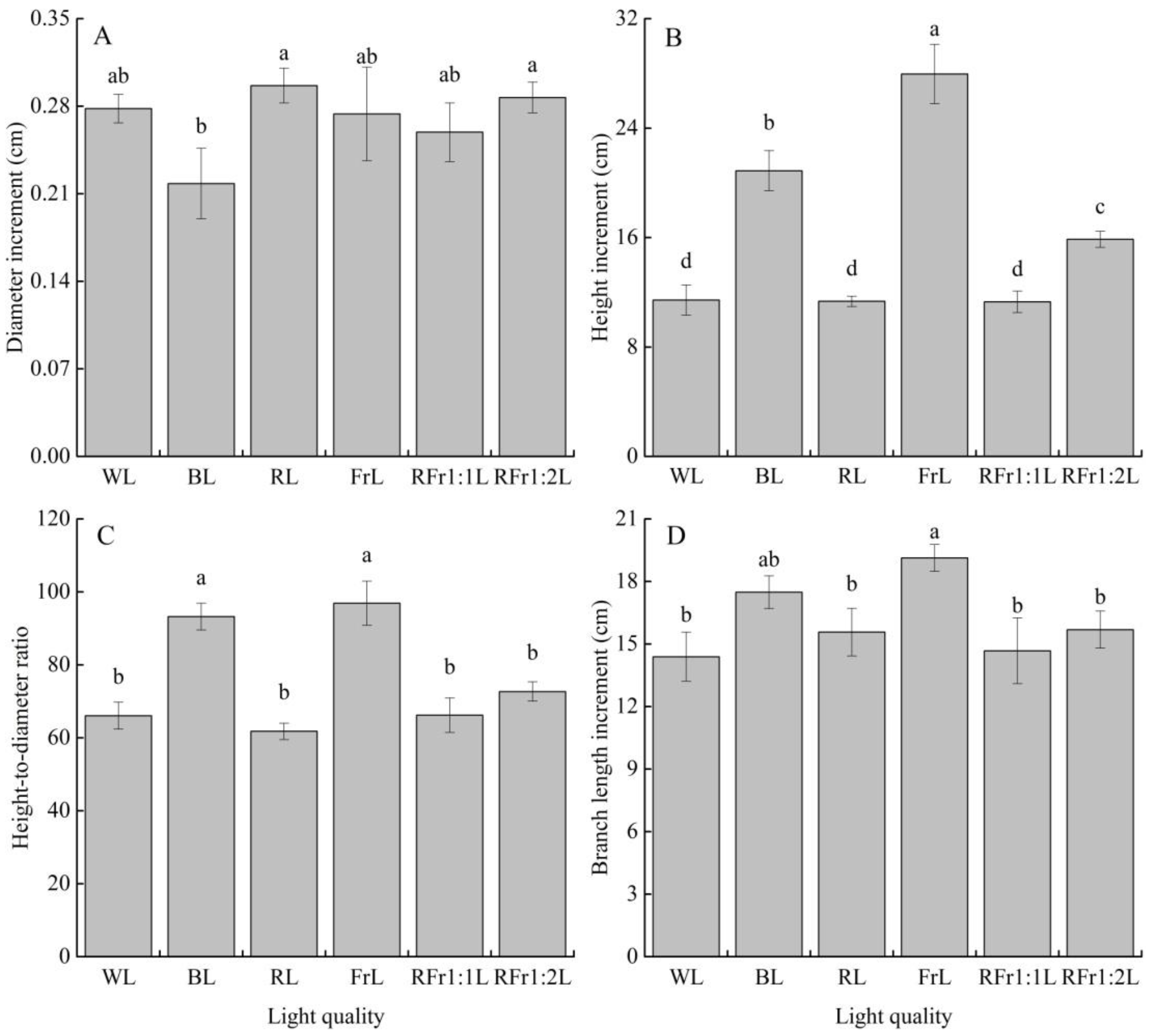
3.1. Seedling Growth Response to Light Quality
3.2. Response of Bud Growth Traits to Light Quality
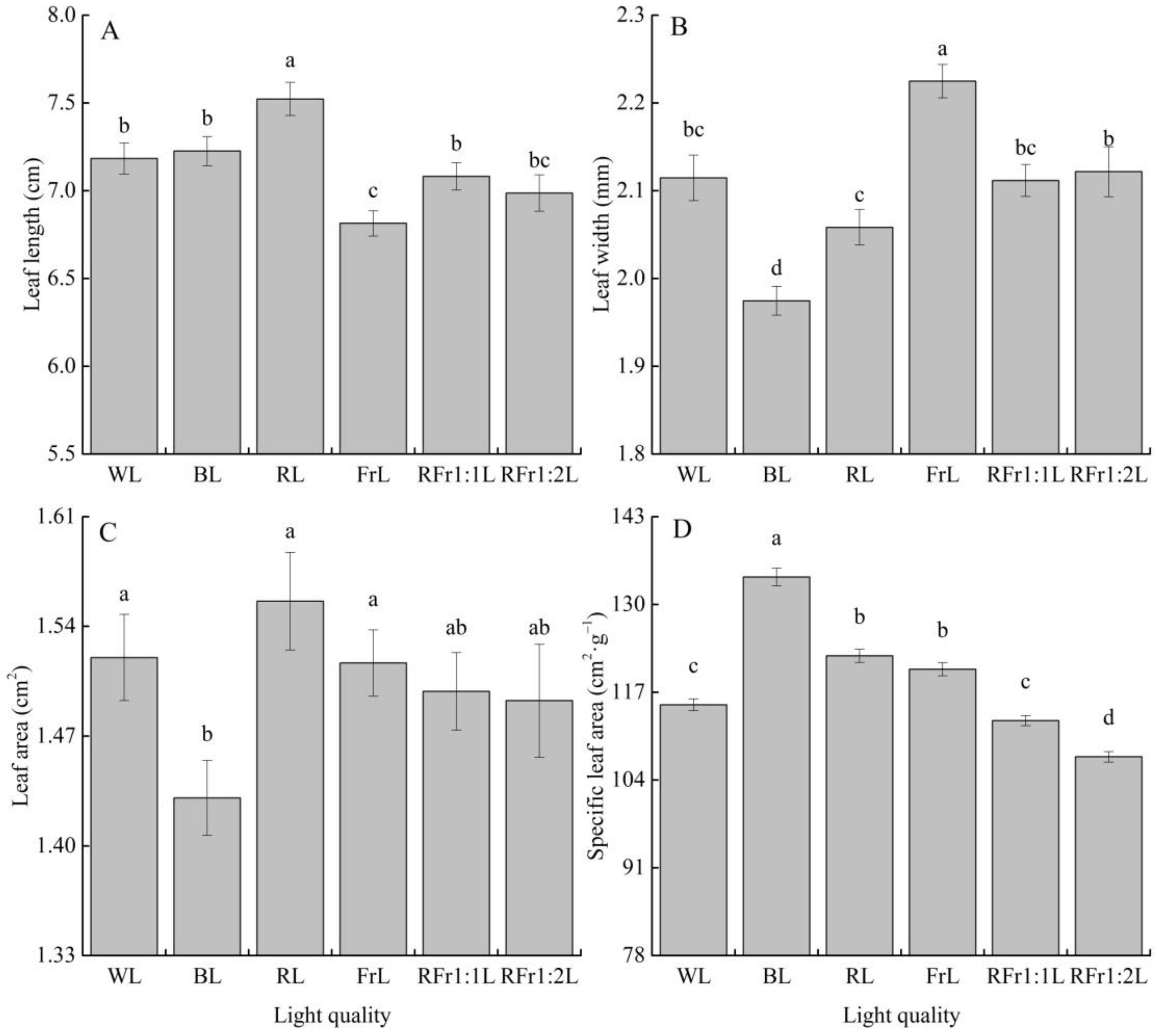
3.3. Response of Leaf Morphological Traits to Light Quality
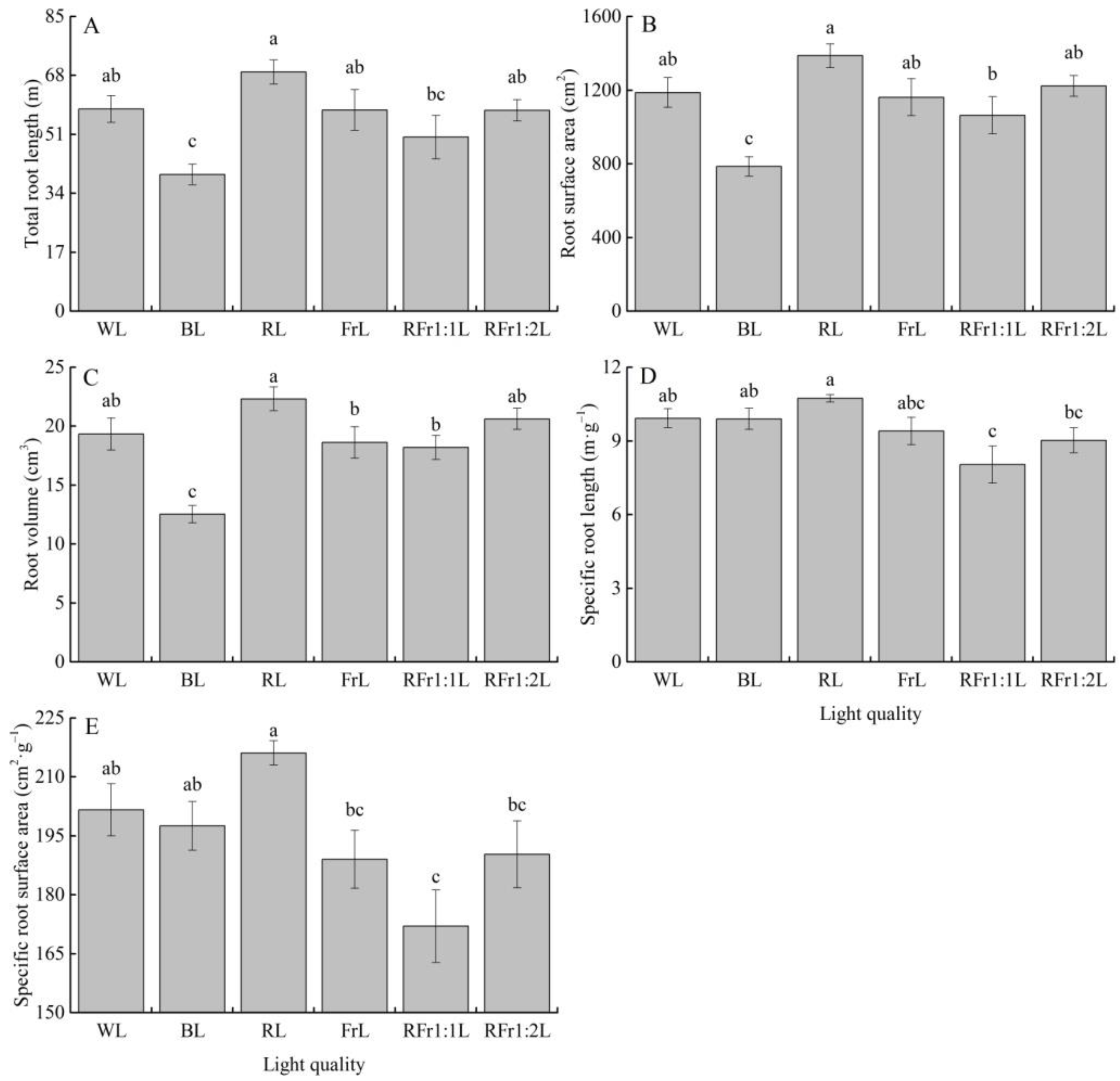
3.4. Response of Root Morphological Traits to Light Quality
3.5. Response of Biomass Accumulation and Allocation to Light Quality
3.6. Phenotypic Plasticity and Patterns of Traits and Light Quality Relations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wirth, R.; Weber, B.; Ryel, R.J. Spatial and temporal variability of canopy structure in a tropical moist forest. Acta Oecol. 2001, 22, 235–244. [Google Scholar] [CrossRef]
- Nascimento, K.C.; Pastorini, L.H.; Romagnolo, M.B.; Souza, L.A.D. Do Eugenia hiemalis seedling leaves under different light conditions develop phenotypic plasticity? Plant Ecol. 2015, 216, 1571–1581. [Google Scholar] [CrossRef]
- Pearcy, R.W. Responses of plants to heterogeneous light environments. In Functional Plant Ecology; Pugnaire, F.I., Valladares, F., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 213–257. [Google Scholar]
- Montgomery, R.; Chazdon, R.J. Light gradient partitioning by tropical tree seedlings in the absence of canopy gaps. Oecologia 2002, 131, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Possart, A.; Fleck, C.; Hiltbrunner, A. Shedding (far-red) light on phytochrome mechanisms and responses in land plants. Plant Sci. 2014, 217, 36–46. [Google Scholar] [CrossRef]
- Godo, T.; Fujiwara, K.; Guan, K.; Miyoshi, K. Effects of wavelength of LED-light on in vitro asymbiotic germination and seedling growth of Bletilla ochracea Schltr. (Orchidaceae). Plant Biotechnol. 2011, 28, 397–400. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Hernández, R.; Kubota, C. Tomato seedling growth and morphological responses to supplemental LED lighting red: Blue rations under varied daily solar light integrals. Acta Hortic. 2012, 956, 187–194. [Google Scholar] [CrossRef]
- Paquette, A.; Bouchard, A.; Cogliastro, A. Morphological plasticity in seedlings of three deciduous species under shelterwood under-planting management does not correspond to shade tolerance ranks. For. Ecol. Manag. 2007, 241, 278–287. [Google Scholar] [CrossRef]
- Takahashi, K.; Seino, T.; Kohyama, T. Responses to canopy openings in architectural development of saplings in eight deciduous broad-leaved tree species. Can. J. For. Res. 2001, 31, 1336–1347. [Google Scholar] [CrossRef]
- Delagrange, S.; Messier, C.; Lechowicz, M.J.; Dizengremel, P. Physiological, morphological and allocational plasticity in understory deciduous trees: Importance of plant size and light availability. Tree Physiol. 2004, 24, 775–784. [Google Scholar] [CrossRef] [Green Version]
- Rolhauser, A.G.; Pucheta, E. Annual plant functional traits explain shrub facilitation in a desert community. J. Veg. Sci. 2016, 27, 60–68. [Google Scholar] [CrossRef]
- Xie, J.B.; Xu, G.Q.; Jenerette, G.D.; Bai, Y.F.; Wang, Z.Y.; Li, Y. Apparent plasticity in functional traits determining competitive ability and spatial distribution: A case from desert. Sci. Rep. 2015, 5, 12174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freschet, G.T.; Violle, C.; Bourget, M.Y.; Scherer-Lorenzen, M.; Fort, F. Allocation, morphology, physiology, architecture: The multiple facets of plant above- and below-ground responses to resource stress. New Phytol. 2018, 219, 1338–1352. [Google Scholar] [CrossRef]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Massa, G.D.; Kim, H.H.; Wheeler, R.M.; Mitchell, C.A. Plant productivity in response to LED lighting. Hortscience. 2008, 43, 1951–1956. [Google Scholar] [CrossRef]
- He, J.; Qin, L.; Liu, Y.M.; Choong, T.W. Photosynthetic capacities and productivity of indoor hydroponically grown Brassica alboglabra bailey under different light sources. Am. J. Plant Sci. 2015, 6, 554–563. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, C.A.; Dzakovich, M.P.; Gomez, C.; Lopez, R.; Burr, J.F.; Hernandez, R.; Kubota, C.; Currey, C.J.; Meng, Q.W.; Runkle, E.S.; et al. Light-emitting diodes in horticulture. Hortic. Rev. 2015, 43, 1–88. [Google Scholar]
- Marcelis, L.F.M.; Elings, A.; Bakker, M.; Brajeul, E.; Dieleman, J.A.; de Visser, P.H.B.; Heuvelink, E. Modelling dry matter production and partitioning in sweet pepper. Acta Hortic. 2006, 718, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, C.A. Plant lighting in controlled environments for space and earth applications. Acta Hortic. 2012, 956, 23–36. [Google Scholar] [CrossRef]
- Marondedze, C.; Liu, X.Y.; Huang, S.H.; Wong, C.; Zhou, X.; Pan, Z.T.; An, H.T.; Xu, N.; Tian, X.C.; Wong, A. Towards a tailored indoor horticulture: A functional genomics guided phenotypic approach. Hortic. Res. 2018, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Hernández, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LEDs. Environ. Exp. Bot. 2016, 29, 66–74. [Google Scholar] [CrossRef]
- OuYang, F.Q.; Ou, Y.; Zhu, T.Q.; Ma, J.W.; An, S.P.; Zhao, J.; Wang, J.H.; Kong, L.S.; Zhang, H.G.; Tigabu, M. Growth and physiological responses of Norway spruce (Piceaabies (L.) H. Karst) supplemented with monochromatic red, blue and far-red light. Forests. 2021, 12, 164. [Google Scholar] [CrossRef]
- Miler, N.; Kulus, D.; Woźny, A.; Rymarz, D.; Hajzer, M.; Wierzbowski, K.; Nelke, R.; Szeffs, L. Application of wide-spectrum light-emitting diodes in micropropagation of popular ornamental plant species: A study on plant quality and cost reduction. In Vitro Cell. Dev. Biol.-Plant. 2019, 55, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Kulus, D.; Woźny, A. Influence of light conditions on the morphogenetic and biochemical response of selected ornamental plant species under in vitro conditions: A mini-review. Bio Technol. 2020, 101, 75–83. [Google Scholar] [CrossRef]
- Kim, H.J.; Lin, M.Y.; Mitchell, C.A. Light spectral and thermal properties govern biomass allocation in tomato through morphological and physiological changes. Environ. Exp. Bot. 2019, 157, 228–240. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Q.Q.; Daryanto, S.; Guo, S.; Huang, Z.J.; Wang, Z.N.; Wang, L.X.; Ma, X.Q. Responses of Chinese fir and Schima superba seedlings to light gradients: Implications for the restoration of mixed broadleaf-conifer forests from Chinese fir monocultures. For. Ecol. Manag. 2018, 419–420, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.J.; Liu, Q.Q.; An, B.; Wu, X.J.; Sun, L.J.; Wu, P.F.; Ma, X.Q. Effects of planting density on morphological and photosynthetic characteristics of leaves in different positions on Cunninghamia lanceolata saplings. Forests 2021, 12, 853. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Laughlin, D.C. Root nutrient concentration and biomass allocation are more plastic than morphological traits in response to nutrient limitation. Plant Soil. 2017, 416, 539–550. [Google Scholar] [CrossRef]
- Messier, J.; McGill, B.J.; Lechowicz, M.J. How do traits vary across ecological scales? A case for trait-based ecology. Ecol. Lett. 2010, 13, 838–848. [Google Scholar] [CrossRef]
- Albert, C.H.; de Bello, F.; Boulangeat, I.; Pellet, G.; Lavorel, S.; Thuiller, W. On the importance of intraspecific variability for the quantification of functional diversity. Oikos 2012, 121, 116–126. [Google Scholar] [CrossRef]
- Zhang, L.W.; Liu, S.Q.; Zhang, Z.K.; Yang, R.; Yang, X.J. Dynamic of different qualities on growth of Toona sinensis seedlings. Acta Agric. Boreali Occident. Sin. 2010, 19, 115–119. (In Chinese) [Google Scholar]
- Li, S.S.; Pan, R.C. Blue light effect of plant. Plant Physiol. Commun. 1993, 29, 248–252. [Google Scholar]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Far-red radiation promotes growth of seedlings by increasing leaf expansion and whole-plant net assimilation. Environ. Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Dou, H.J.; Niu, G.H.; Gu, M.M.; Masabni, J.G. Effects of light quality on growth and phytonutrient accumulation of herbs under controlled environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Devlin, P.F.; Yanovsky, M.J.; Kay, S.A. A genomic analysis of the shade avoidance response in arabidopsis. Plant Physiol. 2003, 133, 1617–1629. [Google Scholar] [CrossRef] [Green Version]
- Whitelam, G.C.; Halliday, K.J. Annual Plant Reviews, Light and Plant Development; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Kurepin, L.V.; Yip, W.K.; Fan, R.; Yeung, E.C.; Reid, D.M. The roles and interactions of ethylene with gibberellins in the far-red enriched light-mediated growth of Solanum lycopersicum seedlings. Plant Growth Regul. 2010, 61, 215–222. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Emery, R.J.N.; Pharis, R.P.; Reid, D.M. The interaction of light quality and irradiance with gibberellins, cytokinins and auxin in regulating growth of Helianthus annuus hypocotyls. Plant Cell Environ. 2007, 30, 147–155. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Emery, R.J.N.; Pharis, R.P.; Reid, D.M. Uncoupling light quality from light irradiance effects in Helianthus annuus shoots: Putative roles for plant hormones in leaf and internode growth. J. Exp. Bot. 2007, 58, 2145–2157. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.J.; Han, L.B.; Song, G.L.; Chen, Y.F. Effect of different ratio of red and blue light emitting diodes on growth and physiological characteristics of two cool-season turfgrass. Pratacult. Sci. 2019, 36, 2831–2837. (In Chinese) [Google Scholar]
- Robin, C.; Hay, M.J.M.; Newton, P.C.D. Effect of light quality (red: Far-red ratio) and defoliation treatments applied at a single phytomer on axillary bud outgrowth in Trifoliumrepens L. Oecologia 1994, 100, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. Lond. Ser. A 2014, 369, 20130243. [Google Scholar] [CrossRef] [PubMed]
- Bugbee, B. In Toward an optimal spectral quality for plant growth and development: The importance of radiation capture. In VIII International Symposium on Light in Horticulture; ISHS: East Lansing, MI, USA, 2016; Volume 22–26, pp. 1–12. [Google Scholar]
- Schoefs, B. Chlorophyll and carotenoid analysis in food products. Properties of the pigments and methods of analysis. Trends Food Sci. Technol. 2002, 13, 361–371. [Google Scholar] [CrossRef]
- Li, H.M.; Xu, Z.G.; Tang, C.M. Effect of light emitting diodes on growth and morphogenesis of upland cotton (Gossypiumhirsutum L.) plantlets in vitro. Plant Cell Tiss. Org. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.; Yoshihara, T. Effect of green light wavelength and intensity on photomorphogenesis and photosynthesis in Lactuca sativa. Environ. Exp. Bot. 2012, 75, 128–133. [Google Scholar] [CrossRef]
- Zhai, S.H.; Wang, P.; Sheng, L.X. Phenotypic plasticity of plant functional traits in competition environments. J. Beihua Univ. 2017, 18, 538–546. (In Chinese) [Google Scholar]
- Bergmann, J.; Ryo, M.; Prati, D.; Hempel, S.; Rillig, M.C. Root traits are more than analogues of leaf traits: The case for diaspore mass. New Phytol. 2017, 216, 1130–1139. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Gu, Q.; Dai, Y.J.; Xu, Z.G. Different light qualities on growth of chrysanthemum (Dendranthema morifolium) plantlets in virto. Chin. Agric. Sci. Bull. 2008, 24, 344–349. (In Chinese) [Google Scholar]
- Yu, W.W.; Liu, Y.; Song, L.L.; Jacobs, D.F.; Du, X.H.; Ying, Y.Q.; Shao, Q.S.; Wu, J.S. Effect of differential light quality on morphology, photosynthesis, and antioxidant enzyme activity in Camptotheca acuminata seedlings. J. Plant Growth Regul. 2017, 36, 148–160. [Google Scholar] [CrossRef]
- Ohashi-Kaneko, K.; Takase, M.; Kon, N.; Fujiwara, K.; Kurata, K. Effect of light quality on growth and vegetable quality in leaf lettuce, spinach and komatsuna. Environ. Control Biol. 2007, 45, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef] [PubMed]
- Saldaña-Acosta, A.; Meave, J.A.; Sánchez-Velásquez, L.R. Seedling biomass allocation and vital rates of cloud forest tree species: Responses to light in shade house conditions. Forest Ecol. Manag. 2009, 258, 1650–1659. [Google Scholar] [CrossRef]
- Nishimura, E.; Suzaki, E.; Irie, M.; Nagashima, H.; Hirose, T. Architecture and growth of an annual plant Chenopodium album in different light climates. Ecol. Res. 2010, 25, 383–393. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Light Quality | |||||
|---|---|---|---|---|---|---|
| WL | BL | RL | FrL | RFr1:1L | RFr1:2L | |
| Bud height (cm) | 3.58 ± 0.32 b | 6.20 ± 0.88 a | 3.44 ± 0.24 b | 7.12 ± 1.00 a | 3.47 ± 0.23 b | 3.51 ± 0.22 b |
| Bud biomass (g) | 1.43 ± 0.25 b | 1.85 ± 0.40 ab | 1.67 ± 0.18 b | 3.14 ± 0.81 a | 2.83 ± 0.63 ab | 1.93 ± 0.36 ab |
| Bud number | 25.50 ± 2.53 c | 12.83 ± 2.02 d | 33.17 ± 2.76 bc | 13.50 ± 3.19 d | 41.67 ± 2.99 a | 34.67 ± 3.19 ab |
| Trait | Light Quality | |||||
|---|---|---|---|---|---|---|
| WL | BL | RL | FrL | RFr1:1L | RFr1:2L | |
| Root biomass (g) | 5.88 ± 0.34 a | 3.97 ± 0.19 b | 6.42 ± 0.27 a | 6.10 ± 0.36 a | 6.10 ± 0.34 a | 6.43 ± 0.13 a |
| Stem biomass (g) | 5.27 ± 0.28 a | 4.90 ± 0.43 a | 5.42 ± 0.24 a | 6.00 ± 0.71 a | 4.59 ± 0.22 a | 5.50 ± 0.30 a |
| Leaf biomass (g) | 9.61 ± 0.47 a | 8.15 ± 0.46 a | 9.00 ± 0.27 a | 8.88 ± 0.79 a | 8.76 ± 0.40 a | 9.57 ± 0.33 a |
| Total biomass (g) | 20.76 ± 1.04 a | 17.02 ± 1.00 a | 20.83 ± 0.59 a | 20.98 ± 1.77 a | 19.45 ± 0.90 a | 21.50 ± 0.56 a |
| Root mass ratio | 0.28 ± 0.01 a | 0.23 ± 0.01 b | 0.31 ± 0.01 a | 0.30 ± 0.02 a | 0.31 ± 0.01 a | 0.30 ± 0.01 a |
| Stem mass ratio | 0.25 ± 0.00 c | 0.29 ± 0.01 a | 0.26 ± 0.01 bc | 0.28 ± 0.01 ab | 0.24 ± 0.00 c | 0.26 ± 0.01 c |
| Leaf mass ratio | 0.46 ± 0.01 ab | 0.48 ± 0.00 a | 0.43 ± 0.01 cd | 0.42 ± 0.00 d | 0.45 ± 0.01 bc | 0.44 ± 0.01 bc |
| Root-to-shoot ratio | 0.40 ± 0.01 b | 0.31 ± 0.02 c | 0.45 ± 0.02 ab | 0.42 ± 0.03 ab | 0.46 ± 0.01 a | 0.43 ± 0.02 ab |
| Group | Trait | CV% | Average CV% |
| Bud growth | Bud height (cm) | 120.11 | 75.59 |
| Bud biomass (g) | 59.60 | ||
| Bud number | 47.06 | ||
| Seedling growth | Height increment (cm) | 41.81 | 25.99 |
| Height-to-diameter ratio | 21.98 | ||
| Diameter increment (cm) | 21.77 | ||
| Branch length increment (cm) | 18.41 | ||
| Root morphology | Total root length (m) | 24.63 | 18.86 |
| Root surface area (cm2) | 22.68 | ||
| Root volume (cm3) | 21.22 | ||
| Specific root length (m·g−1) | 14.91 | ||
| Specific root surface area (cm2·g−1) | 10.87 | ||
| Biomass accumulation and allocation | Stem biomass (g) | 19.30 | 13.71 |
| Root biomass (g) | 18.53 | ||
| Root-to-shoot ratio | 16.89 | ||
| Total biomass (g) | 14.13 | ||
| Leaf biomass (g) | 13.42 | ||
| Root mass ratio | 12.27 | ||
| Stem mass ratio | 9.78 | ||
| Leaf mass ratio | 5.34 | ||
| Leaf morphology | Leaf area (cm2) | 14.51 | 10.65 |
| Leaf length (cm) | 9.88 | ||
| Specific leaf area (cm2·g−1) | 9.47 | ||
| Leaf width (mm) | 8.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Huang, Z.; Ma, X.; Tigabu, M.; Xing, X.; Jin, S.; Liu, B. Phenotypic Plasticity of Cunninghamialanceolata (Lamb.) Hook. Seedlings in Response to Varied Light Quality Treatments. Forests 2022, 13, 201. https://doi.org/10.3390/f13020201
Liu Q, Huang Z, Ma X, Tigabu M, Xing X, Jin S, Liu B. Phenotypic Plasticity of Cunninghamialanceolata (Lamb.) Hook. Seedlings in Response to Varied Light Quality Treatments. Forests. 2022; 13(2):201. https://doi.org/10.3390/f13020201
Chicago/Turabian StyleLiu, Qingqing, Zhijun Huang, Xiangqing Ma, Mulualem Tigabu, Xianshuang Xing, Shaofei Jin, and Bo Liu. 2022. "Phenotypic Plasticity of Cunninghamialanceolata (Lamb.) Hook. Seedlings in Response to Varied Light Quality Treatments" Forests 13, no. 2: 201. https://doi.org/10.3390/f13020201