
Forest Structure and Composition Are Critical to Hurricane Mortality



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Tree Census
2.3. Topographic Exposure
2.4. Accumulated Cyclone Energy at Site
2.5. Significance Test on the Difference of Mortality between Two Events
3. Results
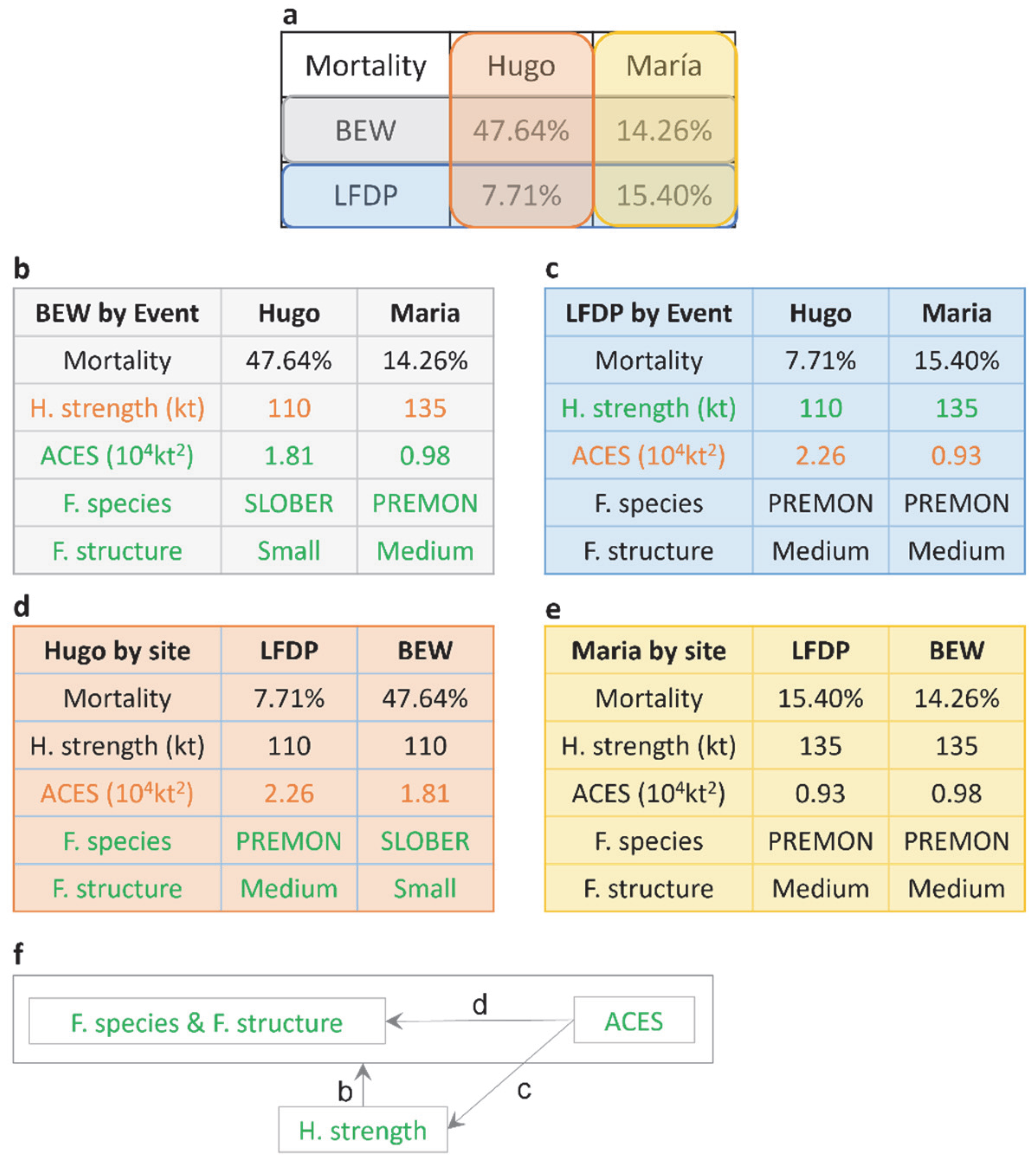
3.1. Observed Damage and Mortality from Hurricanes María and Hugo
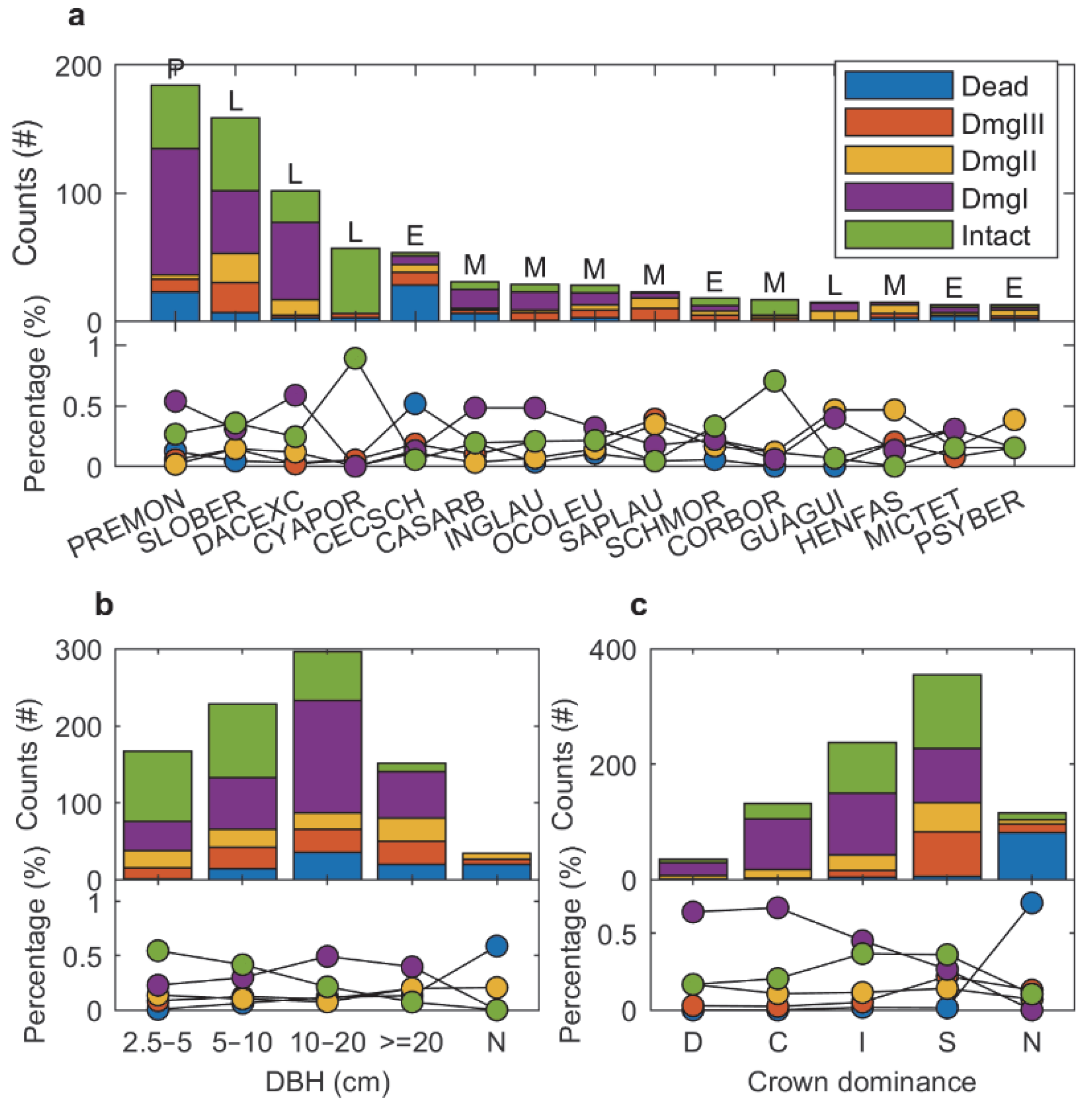
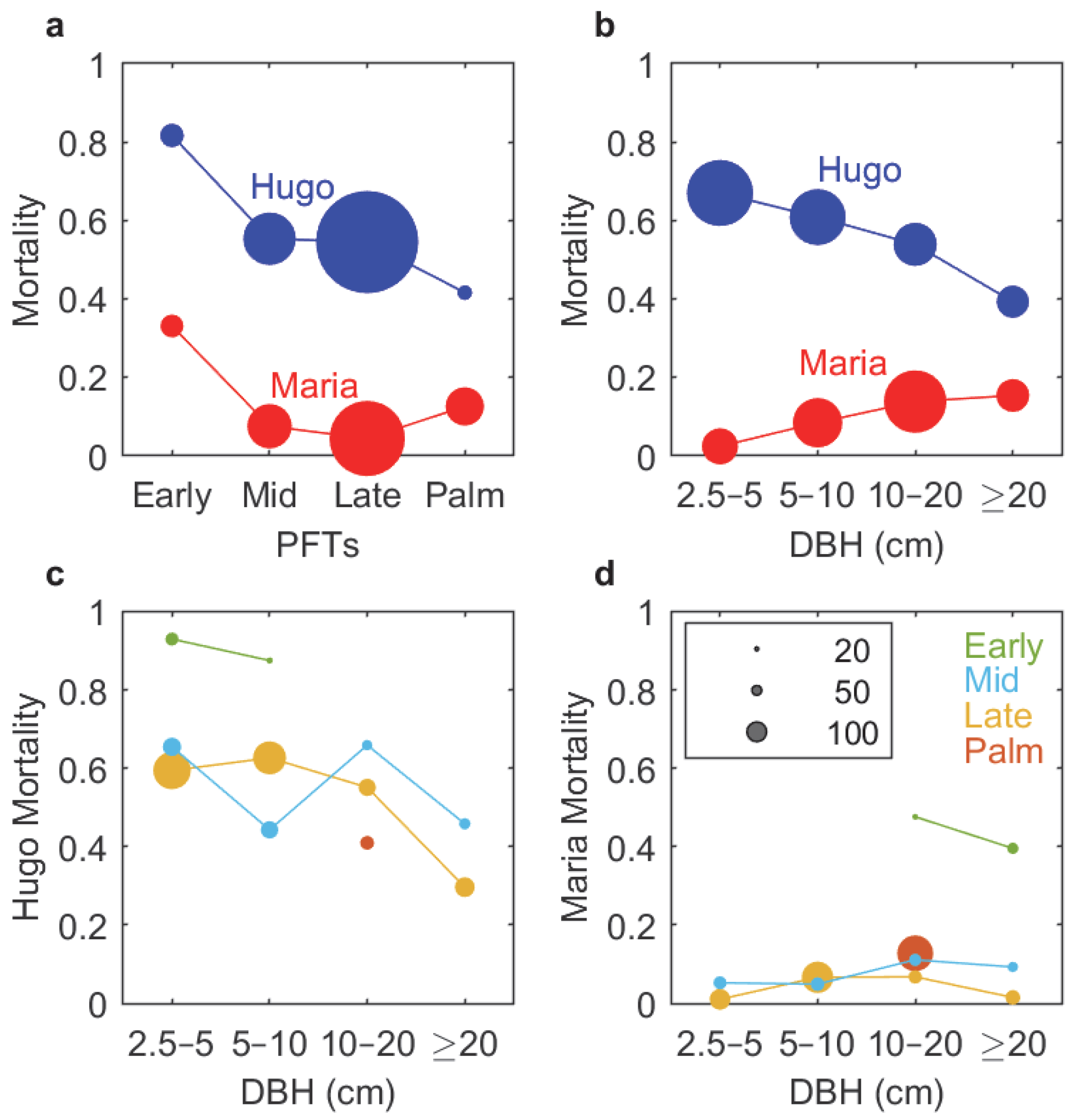
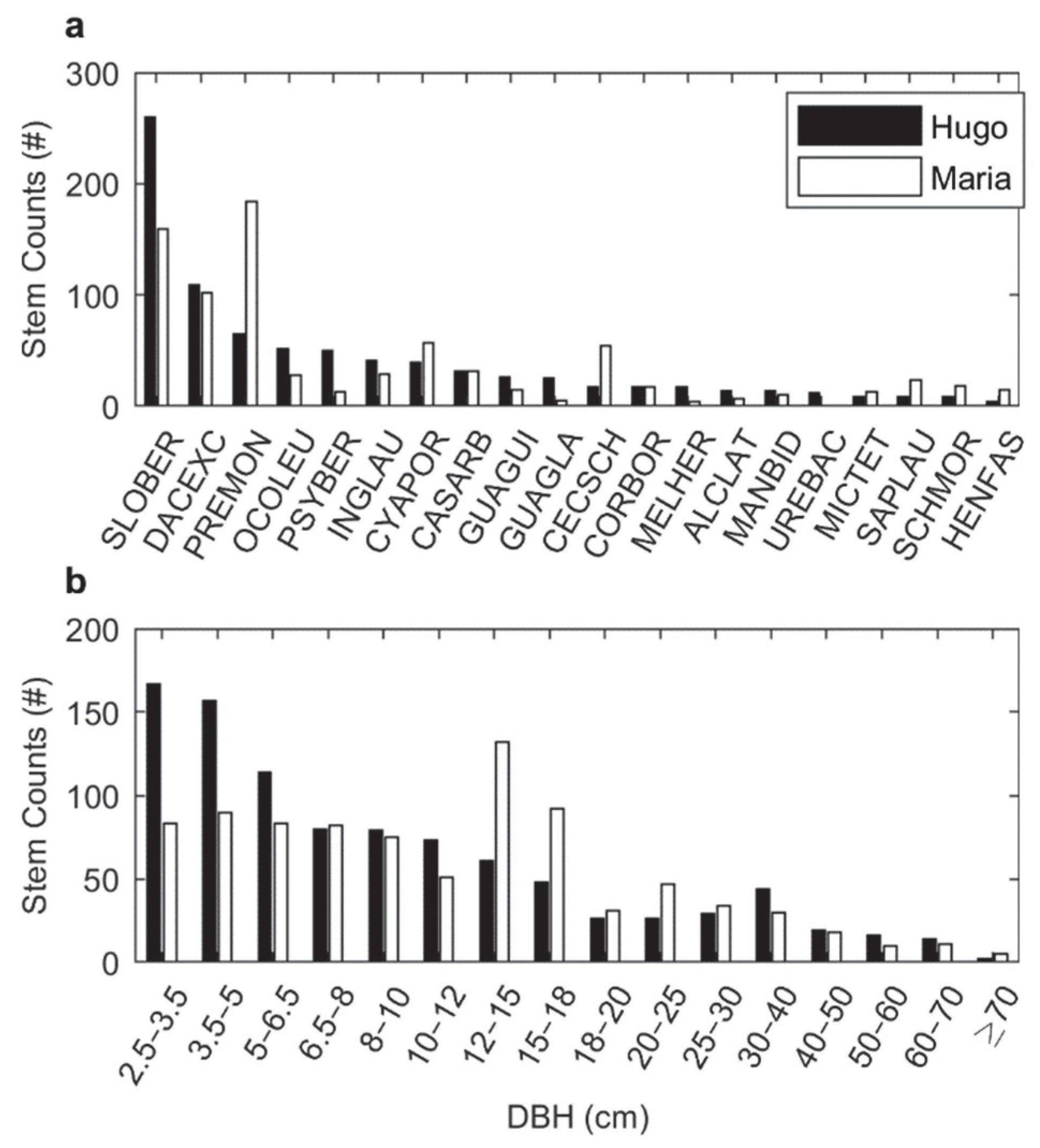
3.2. Forest Structure and Composition
3.3. Forest Exposure to Hurricane Wind
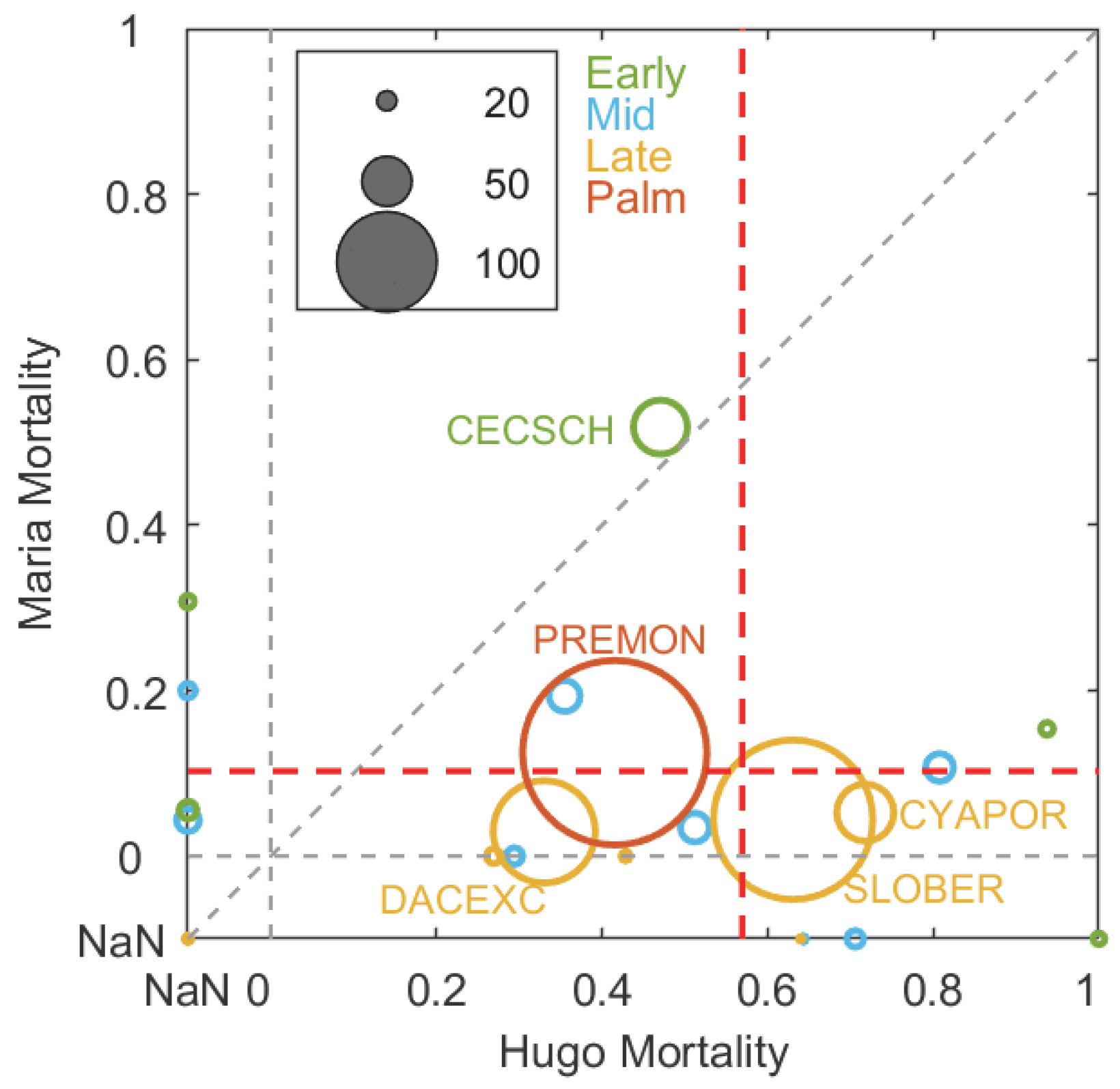
3.4. Same Forest Type and Hurricane Events with Contrasting Site Outcomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lugo, A.E. Visible and invisible effects of hurricanes on forest ecosystems: An international review. Austral Ecol. 2008, 33, 368–398. [Google Scholar] [CrossRef]
- Flynn, D.F.B.; Uriarte, M.; Crk, T.; Pascarella, J.B.; Zimmerman, J.K.; Aide, T.M.; Caraballo Ortiz, M.A. Hurricane disturbance alters secondary forest recovery in Puerto Rico. Biotropica 2010, 42, 149–157. [Google Scholar] [CrossRef]
- Shiels, A.B.; Gonzalez, G.; Lodge, D.L.; Willing, M.R.; Zimmerman, J.K. Cascading effects of canopy opening and debris deposition from a large-scale hurricane experiment in a tropical rain forest. BioScience 2015, 65, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Uriarte, M.; Thompson, J.; Zimmerman, J.K. Hurricane María tripled stem breaks and doubled tree mortality relative to other major storms. Nat. Commun. 2019, 10, 1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heartsill Scalley, T.; Scatena, F.N.; Lugo, A.E.; Moya, S.; Estrada, C.R. Changes in structure, composition, and nutrients during 15 years of hurricane-induced succession in a subtropical wet forest in Puerto Rico. Biotropica 2010, 42, 455–463. [Google Scholar] [CrossRef]
- Heartsill Scalley, T. Insights on forest structure and composition from long-term research in the Luquillo mountains. Forests 2017, 8, 204. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.-C.; Hogan, J.A.; Chang, C.-T. Tropical cyclone ecology: A scale-link perspective. Trends Ecol. Evol. 2020, 35, 594–604. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.-Y.; Shaner, P.-J.L.; Lin, T.-C. Characteristics of old-growth and secondary forests in relation to typhoon disturbance. Ecosystems 2018, 21, 1521–1532. [Google Scholar] [CrossRef]
- Lin, T.-C.; Hamburg, S.P.; Lin, K.-C.; Wang, L.-J.; Chang, C.-T.; Hsia, Y.-J.; Vadeboncoeur, M.A.; McMullen, C.M.M.; Liu, C.-P. Typhoon disturbance and forest dynamics: Lessons from a northwest Pacific subtropical forest. Ecosystems 2011, 14, 127–143. [Google Scholar] [CrossRef]
- Ibanez, T.; Keppel, G.; Menkes, C.; Gillespie, T.W.; Lengaigne, M.; Mangeas, M.; Rivas-Torres, G.; Birnbaum, P. Globally consistent impact of tropical cyclone on the structure of tropical and subtropical forests. J. Ecol. 2019, 107, 279–292. [Google Scholar] [CrossRef]
- Vandermeer, J.; Boucher, D.; Perfecto, I.; de la Cerda, I.G. A theory of disturbance and species diversity: Evidence from Nicaragua after hurricane Joan. Biotropica 1996, 28, 600–613. [Google Scholar] [CrossRef]
- Xi, W. Synergistic effects of tropical cyclones on forest ecosystems: A global synthesis. J. For. Res. 2015, 26, 1–21. [Google Scholar] [CrossRef]
- Xi, W.; Peet, R.K.; Urban, D.L. Changes in forest structure, species diversity and spatial pattern following hurricane disturbance in a Piedmont North Carolina forest, USA. J. Plant Ecol. 2008, 1, 43–57. [Google Scholar] [CrossRef]
- Arnone, E.; Noto, L.V.; Lepore, C.; Bras, R.L. Physically-based and distributed approach to analyze rainfall-triggered landslides at watershed scale. Geomorphology 2011, 133, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Arnone, E.; Dialynas, Y.G.; Noto, L.V.; Bras, R.L. Accounting for soil parameter uncertainty in a physically based and distributed approach for rainfall-triggered landslides. Hydrol. Process. 2015, 30, 927–944. [Google Scholar] [CrossRef]
- Lepore, C.; Kamal, S.A.; Shanahan, P.; Bras, R.L. Rainfall-induced landslide susceptibility zonation of Puerto Rico. Environ. Earth Sci. 2012, 66, 1667–1681. [Google Scholar] [CrossRef]
- Lepore, C.; Arnone, E.; Noto, L.V.; Sivandran, G.; Bras, R.L. Physically based modeling of rainfall-triggered landslides: A case study in the Luquillo forest, Puerto Rico. Hydrol. Earth Syst. Sci. 2013, 17, 3371–3387. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.R.; Dracup, E.; MacLean, D.A.; Boulanger, Y.; Endicott, S. Forest structure more important than topography in determining windthrow during Hurricane Juan in Canada’s Acadian Forest. For. Ecol. Manag. 2019, 434, 255–263. [Google Scholar] [CrossRef]
- Boose, E.R.; Chamberlin, K.E.; Foster, D.R. Landscape and regional impacts of hurricanes in New England. Ecol. Monogr. 2001, 71, 27–48. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Tanner, E.V.J.; Healey, J.R. Damage and responsiveness of Jamaican montane tree species after disturbance by a hurricane. Ecology 1995, 76, 2562–2580. [Google Scholar] [CrossRef]
- Liebsch, D.; Marcilio-Silva, V.; Marcon, A.K.; Galvão, F.; Mikich, S.B.; Marques, M.C.M. How do trees survive a cyclone? The relative role of individual and site characteristics over mortality. Aus. Ecol. 2021, 46, 1356–1365. [Google Scholar] [CrossRef]
- Lewis, R.J.; Bannar-Martin, K.H. The impact of cyclone Fanele on a tropical dry forest in Madagascar. Biotropica 2012, 44, 135–140. [Google Scholar] [CrossRef]
- Ni, Y.; Wang, T.; Cao, H.; Li, Y.; Bin, Y.; Zhang, R.; Wang, Y.; Lian, J.; Ye, W. An old-growth subtropical evergreen broadleaved forest suffered more damage from Typhoon Mangkhut than an adjacent secondary forest. For. Ecol. Manag. 2021, 496, 119433. [Google Scholar] [CrossRef]
- Tanner, E.V.J.; Kapos, V.; Healey, J.R. Hurricane effects on forest ecosystems in the Caribbean. Biotropica 1991, 23, 513–521. [Google Scholar] [CrossRef]
- Chirici, G.; Bottalico, F.; Giannetti, F.; Del Perugia, B.; Travaglini, D.; Nocentini, S.; Kutchartt, E.; Marchi, E.; Foderi, C.; Fioravanti, M.; et al. Assessing forest windthrow damage using single-date, post-event airborne laser scanning data. Forestry 2018, 91, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, J.N.; Little, A.G.; Majumdar, S.J.; Schoener, T.W.; Fisher, D.N. Call-to-action: A global consortium for tropical cyclone ecology. Trends Ecol. Evol. 2019, 34, 588–590. [Google Scholar] [CrossRef]
- Hogan, J.A.; Feagin, R.A.; Starr, G.; Ross, M.; Lin, T.C.; O’connell, C.; Huff, T.P.; Stauffer, B.A.; Robinson, K.L.; Lara, M.C.; et al. A research framework to integrate cross-ecosystem responses to tropical cyclones. BioScience 2020, 70, 477–489. [Google Scholar] [CrossRef]
- Scatena, F.N. An Introduction to the Physiography and History of the Bisley Experimental Watersheds in the Luquillo Mountains of Puerto Rico; Gen. Tech. Rep. SO-72.; U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1989; 22p. [Google Scholar]
- Castro-Rivera, A.; López-Marrero, T. Allá viene el temporal: Major hurricanes in Puerto Rico between 1851 and 2019. Caribb. Geogr. 2019, 24, 24–39. [Google Scholar]
- Zimmerman, J.K.; Wood, T.E.; González, G.; Ramirez, A.; Silver, W.L.; Uriarte, M.; Willig, M.R.; Waide, R.B.; Lugo, A.E. Disturbance and resilience in the Luquillo Experimental Forest. Biol. Conserv. 2021, 253, 108891. [Google Scholar] [CrossRef]
- Zhang, J.; Heartsill Scalley, T.; Bras, R.L. Tree Census at Bisley Experimental Watersheds since Hurricane Hugo in 1989 to 2014; Forest Service Research Data Archive: Fort Collins, CO, USA, 2022; in preparation. [Google Scholar]
- Zhang, J.; Bras, R.L.; Heartsill Scalley, T. Tree Census at Bisley Experimental Watersheds Three Months after Hurricane María; Forest Service Research Data Archive: Fort Collins, CO, USA, 2020. [Google Scholar]
- Walker, L.R. Tree damage and recovery from hurricane Hugo in Luquillo Experimental Forest, Puerto Rico. Part A. special issue: Ecosystem, plant, and animal responses to hurricanes in the Caribbean. Biotropica 1991, 23, 379–385. [Google Scholar] [CrossRef]
- Schowalter, T.D.; Ganio, L.M. Invertebrate communities in a tropical rain forest canopy in Puerto Rico following hurricane Hugo. Ecol. Entomol. 1999, 24, 191–201. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Brokaw, N. Seedling recruitment in a hurricane-driven tropical forest: Light limitation, density-dependence and the spatial distribution of parent trees. J. Ecol. 2005, 93, 291–304. [Google Scholar] [CrossRef]
- Muscarella, R.; Uriarte, M.; Forero-Montaña, J.; Comita, L.S.; Swenson, N.G.; Thompson, J.; Nytch, C.J.; Jonckheere, I.; Zimmerman, J.K. Life-history trade-offs during the seed-to-seedling transition in a subtropical wet forest community. J. Ecol. 2013, 101, 171–182. [Google Scholar] [CrossRef]
- Feng, X.; Uriarte, M.; González, G.; Reed, S.; Thompson, J.; Zimmerman, J.K.; Murphy, L. Improving predictions of tropical forest response to climate change through integration of field studies and ecosystem modeling. Glob. Chang. Biol. 2018, 24, e213–e232. [Google Scholar] [CrossRef] [PubMed]
- Boose, E.R.; Serrano, M.I.; Foster, D.R. Landscape and regional impacts of hurricanes in Puerto Rico. Ecol. Monogr. 2004, 74, 335–352. [Google Scholar] [CrossRef]
- Boose, E.R.; Foster, D.R.; Fluet, M. Hurricane Impacts of tropical and temperature forest landscapes. Ecol. Monogr. 1994, 64, 369–400. [Google Scholar] [CrossRef]
- USGS. 1/3-Arc Second National Elevation Dataset; US Geological Survey: Reston, VA, USA, 2018. [Google Scholar]
- Parker, L.; Booth, W. Hurricane Hugo Rips through South Carolina. The Washington Post, Friday. 22 September 1989. Available online: https://www.washingtonpost.com/wp-srv/weather/hurricane/poststories/hugo-sc.htm(accessed on 25 January 2022).
- Pasch, R.J.; Penny, A.B.; Berg, R.; National Hurricane Center. Tropical Cyclone Report: Hurricane María (AL152017). 2019. Available online: https://www.nhc.noaa.gov/data/tcr/AL152017_Maria.pdf (accessed on 25 January 2022).
- Landsea, C.W.; Franklin, J.L. Atlantic hurricane database uncertainty and presentation of a new database format. Mon. Weather Rev. 2013, 141, 3576–3592. [Google Scholar] [CrossRef]
- Bell, G.D.; Halpert, M.S.; Ropelewski, C.F.; Kousky, V.E.; Douglas, A.V.; Schnell, R.C.; Gelman, M.E. Climate assessment for 1999. Bull. Am. Meteorol. Soc. 2000, 81, S1–S50. [Google Scholar] [CrossRef]
- Montgomery, D.C.; Runger, G.C. Large-sample tests on the difference in population proportions. In Applied Statistics and Probability for Engineers, 7th ed.; Wiley: New York, NY, USA, 2018; pp. 274–275. ISBN 978-1-119-40036-3. [Google Scholar]
- Griffith, M.P.; Noblick, L.R.; Dowe, J.L.; Husby, C.E.; Calonje, M.A. Cyclone tolerance in New World Arecaceae: Biogeographic variation and abiotic natural selection. Ann. Bot. 2008, 102, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Luke, D.; McLaren, K.; Wilson, B. Modeling hurricane exposure in a Caribbean lower montane tropical wet forest: The effects of frequent, intermediate disturbances and topography on forest structural dynamics and composition. Ecosystems 2016, 19, 1178–1195. [Google Scholar] [CrossRef]
- Crow, T.R. A rainforest chronicle: A 30-year record of change in structure and composition at El Verde, Puerto Rico. Biotropica 1980, 12, 42–55. [Google Scholar] [CrossRef]
- Emanuel, K. Increasing destructiveness of tropical cyclones over the past 30 years. Nature 2005, 436, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Knutson, T.R.; McBride, J.L.; Chan, J.; Emanuel, K.; Holland, G.; Landsea, C.; Held, I.; Kossin, J.P.; Srivastava, A.K.; Sugi, M. Tropical cyclones and climate change. Nat. Geosci. 2010, 3, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Bender, M.A.; Knutson, T.R.; Tuleya, R.E.; Sirutis, J.J.; Vecchi, G.A.; Garner, S.T.; Held, I.M. Modeled impact of anthropogenic warming on the frequency of intense Atlantic Hurricanes. Science 2010, 327, 454–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandermeer, J.; de la Cerda, I.G.; Boucher, D.; Perfecto, I.; Ruiz, J. Hurricane disturbance and tropical tree species diversity. Science 2000, 290, 788–791. [Google Scholar] [CrossRef]
- Kossin, J.; Emanuel, K.; Vecchi, G. The poleward migration of the location of tropical cyclone maximum intensity. Nature 2014, 509, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Corona, P. Consolidating new paradigms in large-scale monitoring and assessment of forest ecosystems. Environ. Res. 2016, 144, 8–14. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Heartsill-Scalley, T.; Bras, R.L. Forest Structure and Composition Are Critical to Hurricane Mortality. Forests 2022, 13, 202. https://doi.org/10.3390/f13020202
Zhang J, Heartsill-Scalley T, Bras RL. Forest Structure and Composition Are Critical to Hurricane Mortality. Forests. 2022; 13(2):202. https://doi.org/10.3390/f13020202
Chicago/Turabian StyleZhang, Jiaying, Tamara Heartsill-Scalley, and Rafael L. Bras. 2022. "Forest Structure and Composition Are Critical to Hurricane Mortality" Forests 13, no. 2: 202. https://doi.org/10.3390/f13020202