
The Photosynthesis of Populus euphratica Oliv. Is Not Limited by Drought Stress in the Hyper-Arid Zone of Northwest China

Abstract
:1. Introduction
2. Materials and Methods
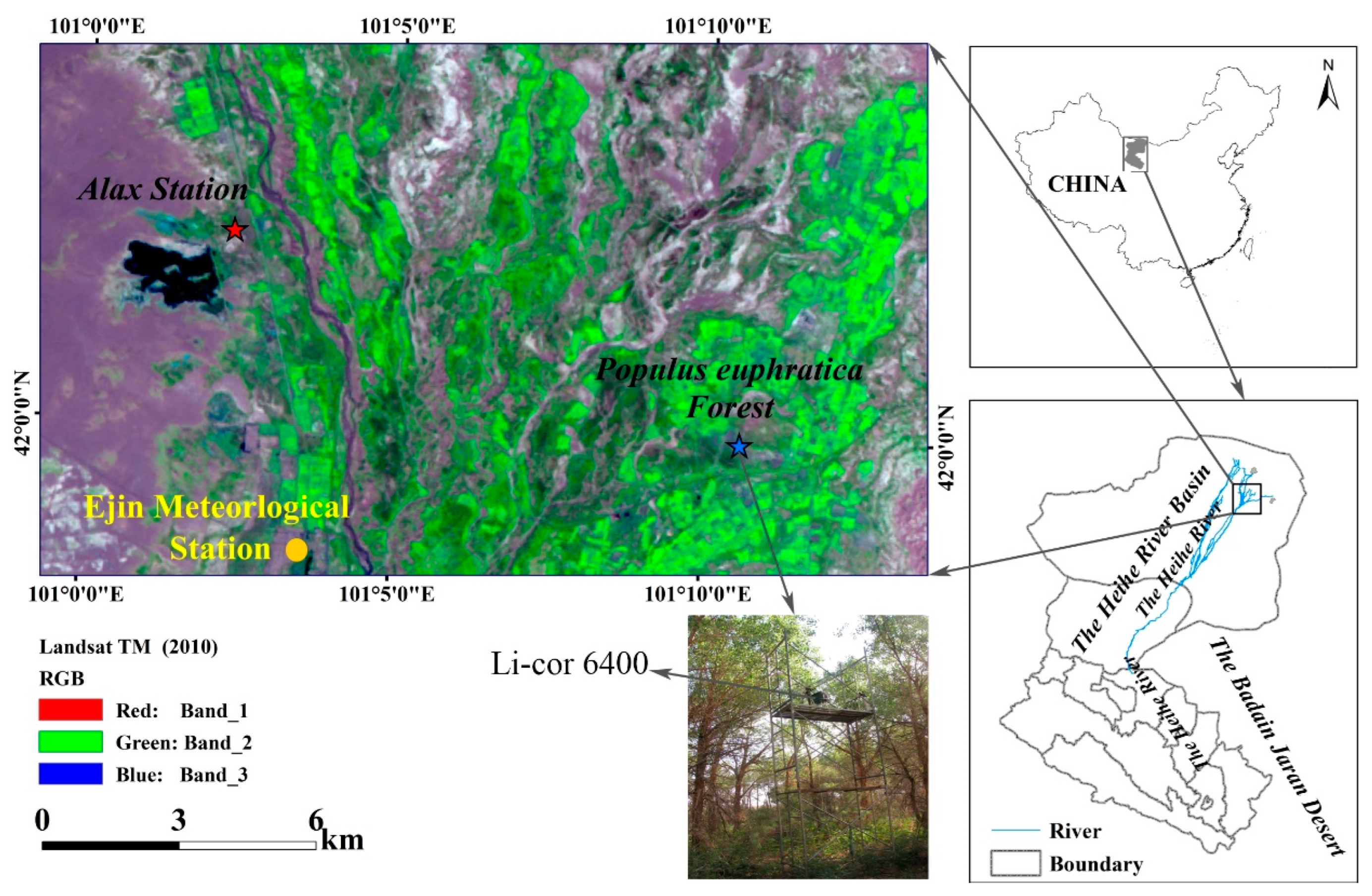
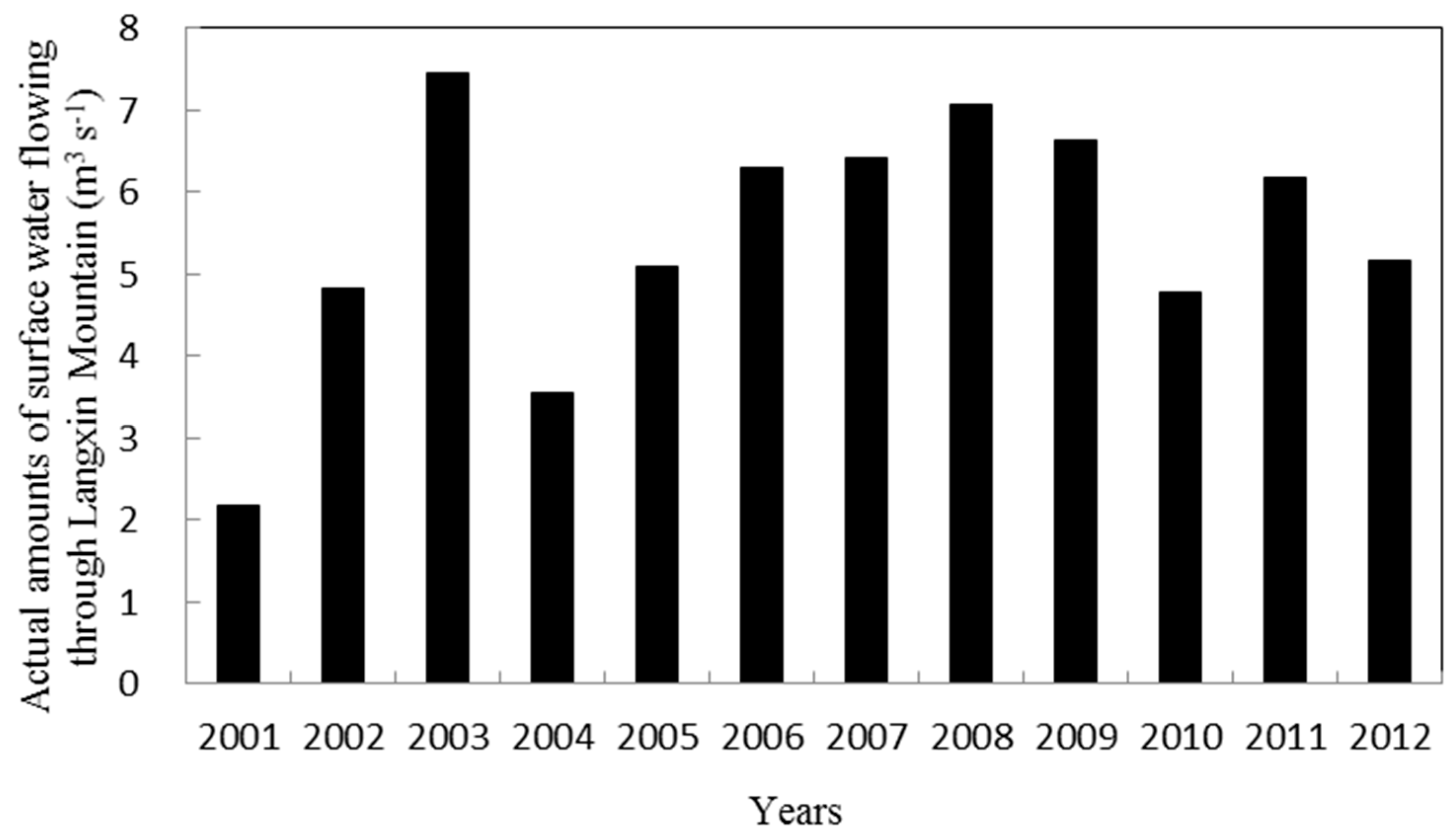
2.1. Experimental Site
2.2. Sap Flow and Transpiration
2.3. Leaf Gas-Exchange and Water Potential
2.4. Meteorological and Hydrologic Parameters
2.5. Data Analysis
2.6. Formulae
2.6.1. Stomatal, Non-Stomatal, and Relative Stomatal Limitations of Photosynthesis and Related Parameters
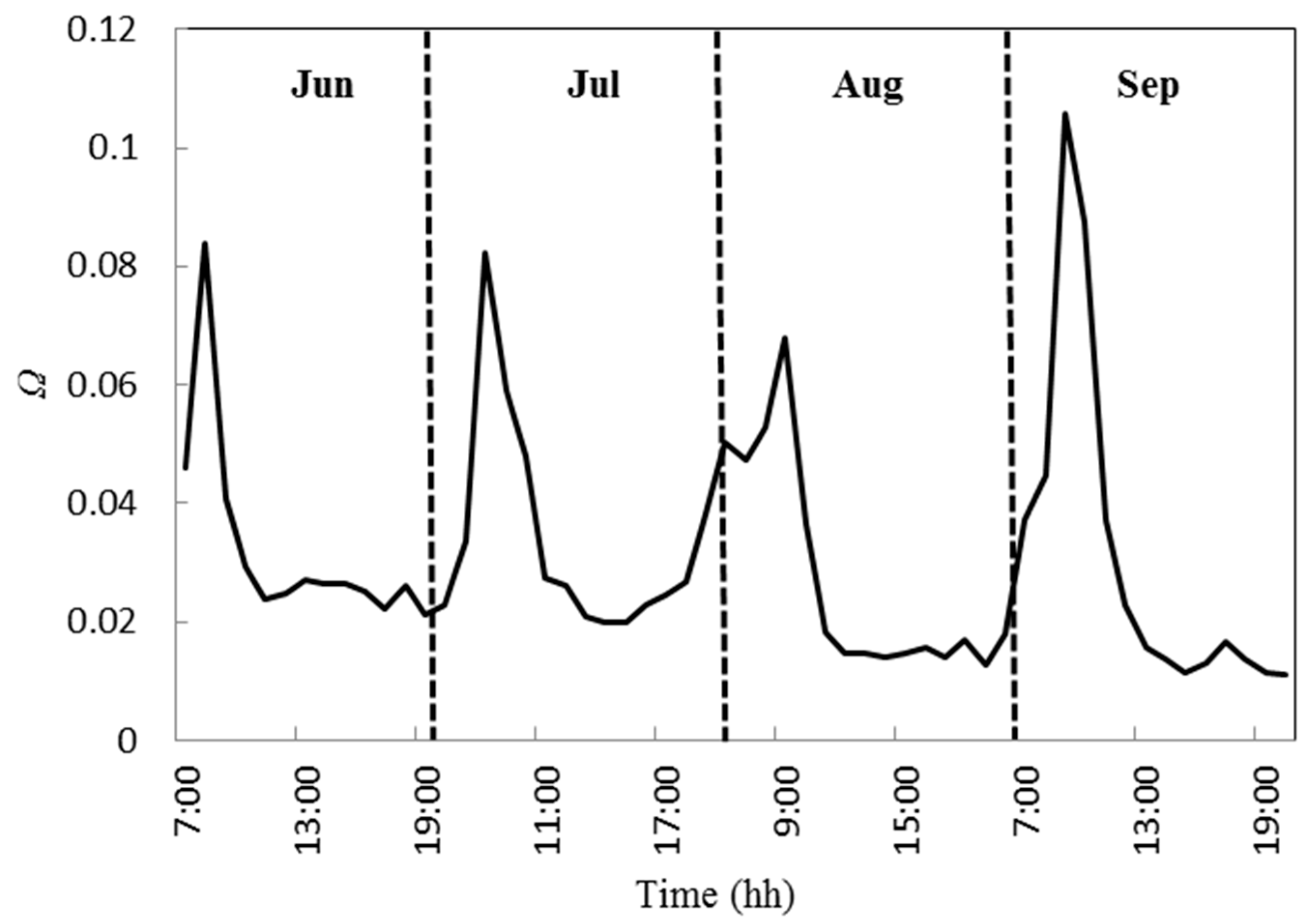
2.6.2. Decoupling Coefficient and Related Parameters
3. Results
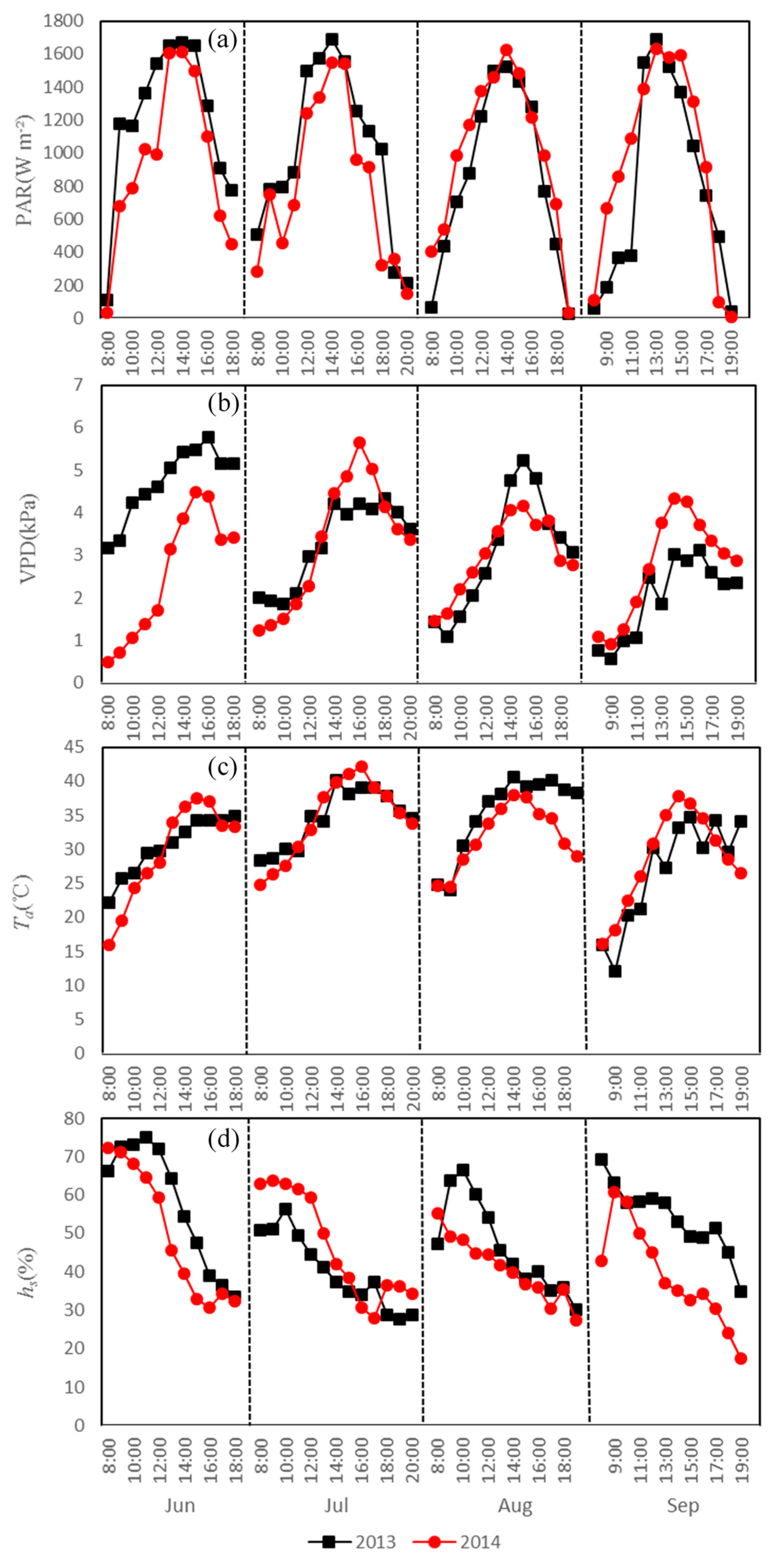
3.1. Diurnal Variations in Main Environmental Factors
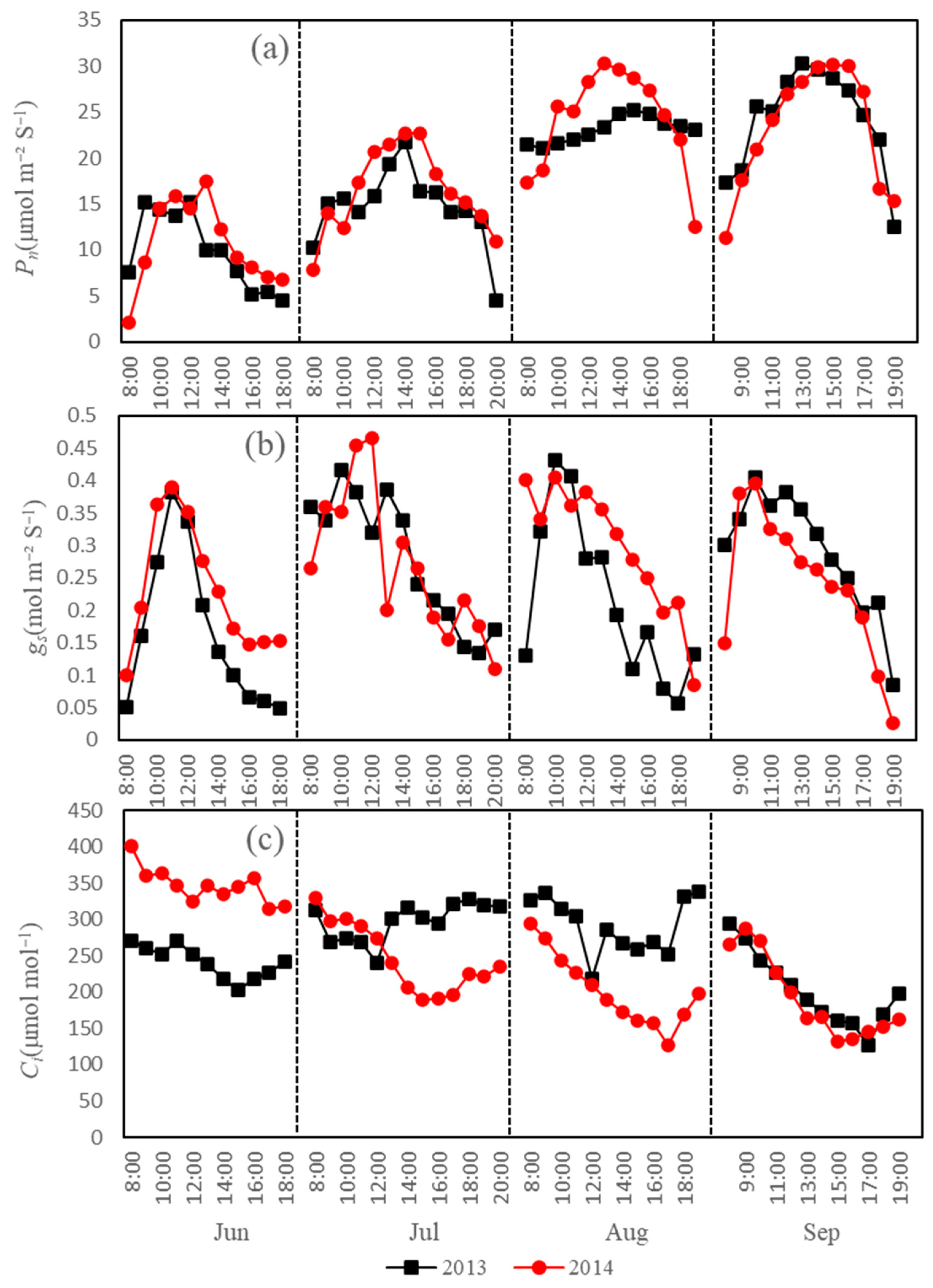
3.2. Diurnal Variations in the Main Physiological Factors
3.3. Relationships between Environmental Parameters and Pn
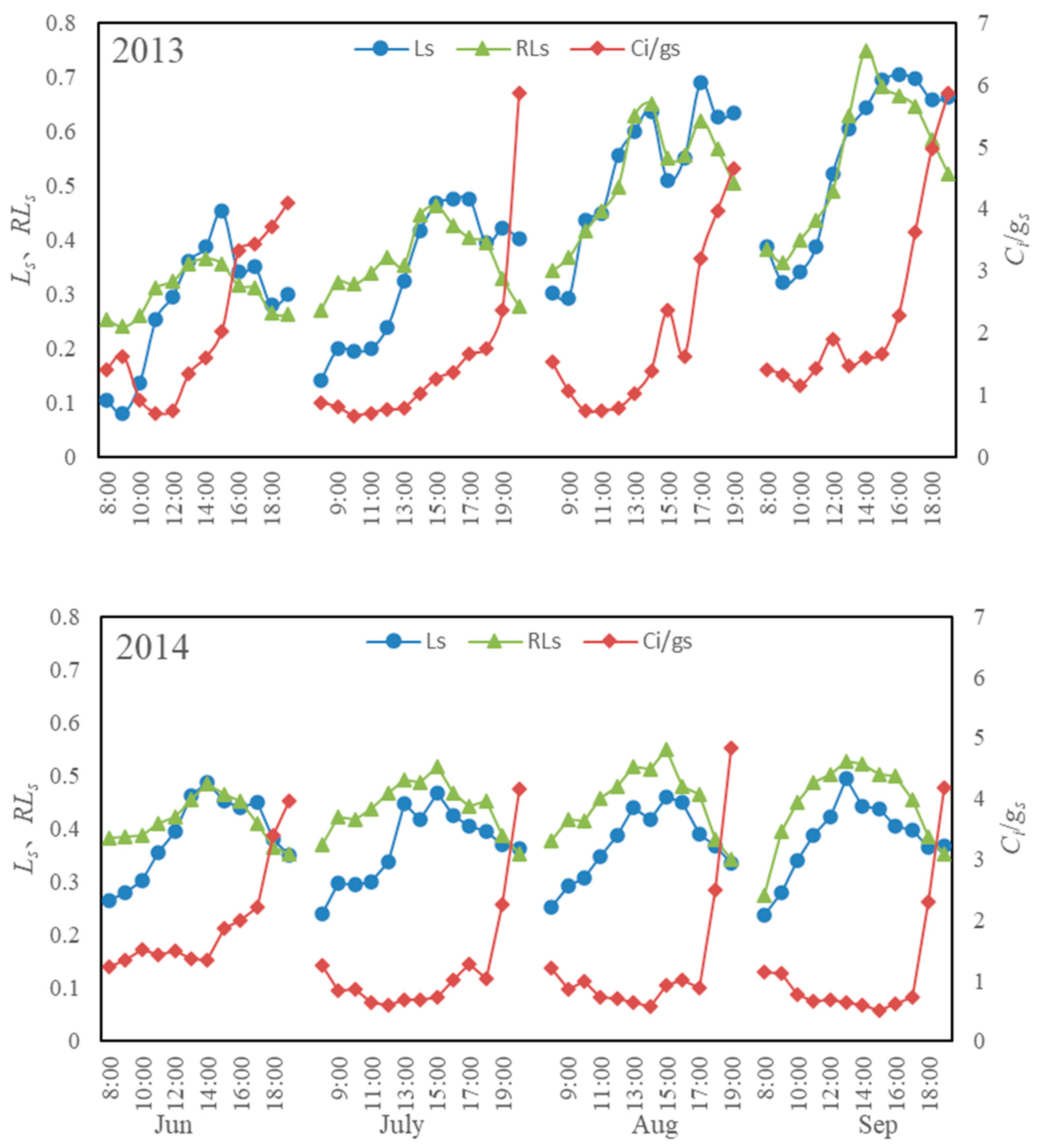
3.4. Diurnal Variations in Stomatal, Relative Stomatal, and Non-Stomatal Limitations of Photosynthesis
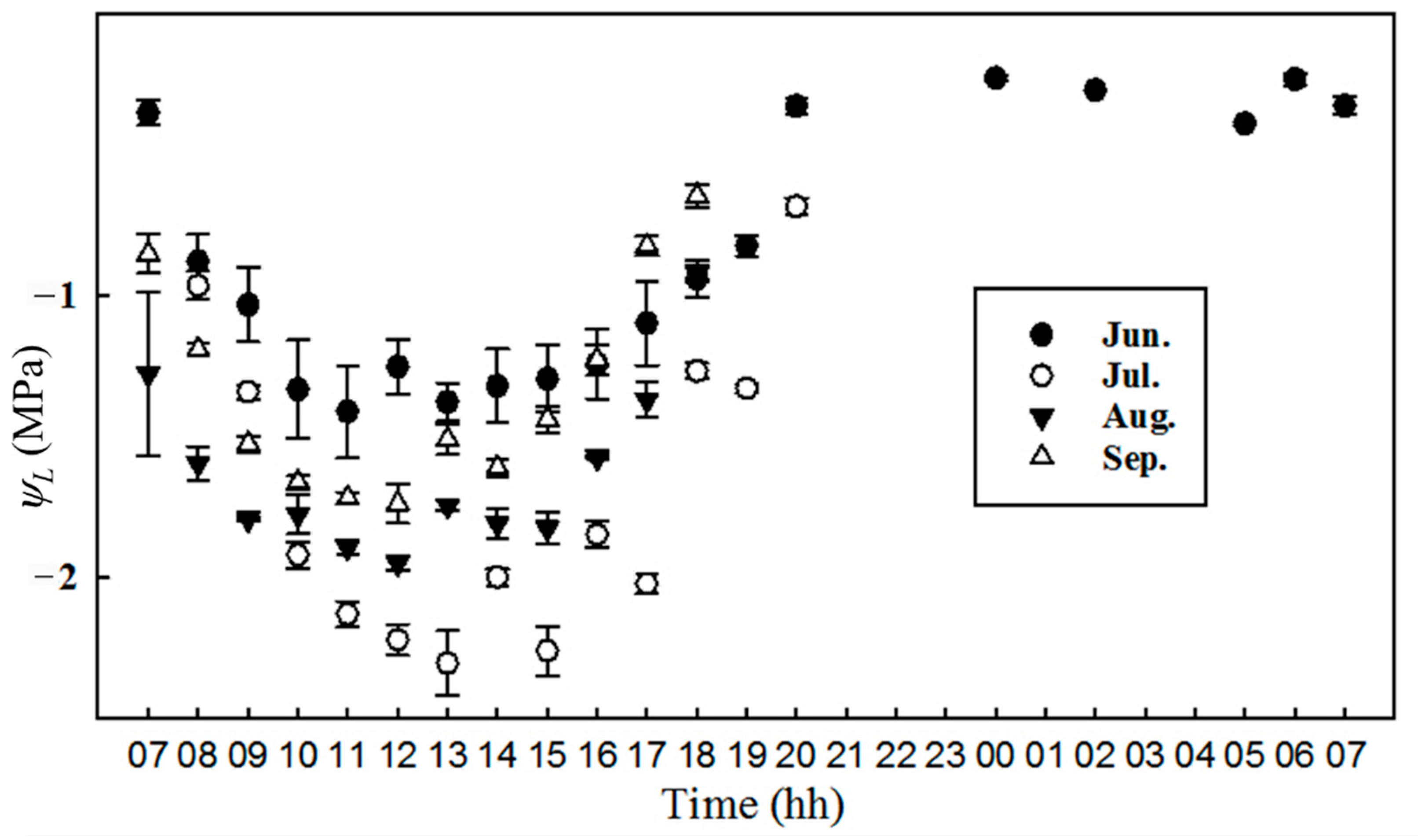
3.5. Variations in Parameters Related to Water Conditions
4. Discussion
4.1. Variations in Physiological Factors and the Phenomenon of Midday Depression of Photosynthesis
4.2. Stomatal, Relative Stomatal, and Non-Stomatal Limitations of Photosynthesis of P. euphratica
4.3. Is the Photosynthesis of P. euphratica Limited by Drought Stress?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Maximum Ls values | Location | Source of Data |
|---|---|---|---|
| Dacrydium cupressinum Lamb. | 0.29 | New Zealand | [74] |
| Meterosideros umbellate Cav. | 0.41 | ||
| Weinmannia racemosa L.f. | 0.42 | ||
| Quintinia acutifolia Kirk. | 0.41 | ||
| Quercus robur L. | 0.17 | Italy | [57] |
| Fraxinus oxyphylla Bieb. | 0.21 | ||
| Spring wheat | 0.64 | China | [51] |
| Populus euphratica Oliv. | 0.70 (2013) 0.51 (2014) | China | This study |
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, D.; Si, J.H.; Zhang, X.Y.; Gao, Y.Y.; Wang, C.L.; Luo, H.; Qin, J.; Gao, G.L. The mechanism of changes in hydraulic properties of Populus euphratica in response to drought stress. Forests 2019, 10, 904. [Google Scholar] [CrossRef] [Green Version]
- Li, C.Y.; Wang, K.Y. Differences in drought responses of three contrasting Eucalyptus microtheca F. Muell. Populations. For. Ecol. Manag. 2003, 179, 377–385. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, N.; Li, C. Physiological and growth responses of Populus davidiana ecotypes to different soil water contents. J. Arid Environ. 2005, 60, 567–579. [Google Scholar] [CrossRef]
- Flexas, J.; Barón, M.; Bota, J.; Ducruet, J.M.; Gallé, A.; Galmés, J.; Jiménez, M.; Pou, A.; Ribas-Carbó, M.; Sajnani, C.; et al. Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri × V. rupestris). J. Exp. Bot. 2009, 60, 2361–2377. [Google Scholar] [CrossRef] [Green Version]
- Makino, A. Photosynthesis, grain yield, and nitrogen utilization in rice and wheat. Plant Physiol. 2011, 155, 125–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Jones, H.G. Stomatal control of photosynthesis and transpiration. J. Exp. Bot. 1998, 49, 387–398. [Google Scholar] [CrossRef]
- Signarbieux, C.; Feller, U. Non-stomatal limitations of photosynthesis in grassland species under artificial drought in the field. Environ. Exp. Bot. 2011, 71, 192–197. [Google Scholar] [CrossRef]
- Varone, L.; Ribas-Carbo, M.; Cardona, C.; Gallé, A.; Medrano, H.; Gratani, L.; Flexas, J. Stomatal and non-stomatal limitations to photosynthesis in seedlings and saplings of Mediterranean species pre-conditioned and aged in nurseries: Different response to water stress. Environ. Exp. Bot. 2012, 75, 235–247. [Google Scholar] [CrossRef]
- Campos, H.; Trejo, C.; Peña-Valdivia, C.B.; Garcia-Nava, R.; Conde-MartÍnez, F.V.; Cruz-Ortega, M.R. Stomatal and non-stomatal limitations of bell pepper (Capsicum annuum L.) plants under water stress and re-watering: Delayed restoration of photosynthesis during recovery. Environ. Exp. Bot. 2014, 98, 56–64. [Google Scholar] [CrossRef]
- Anev, S.; Marinova, A.; Tzvetkova, N.P.; Panayotov, M.P.; Yurukov, S. Stomatal control on photosynthesis in drought-treated subalpine pine saplings. Genet. Plant Physiol. 2016, 6, 43–53. [Google Scholar]
- Bastin, J.F.; Berrahmouni, N.; Grainger, A.; Maniatis, D.; Mollicone, D.; Moore, R.; Patriarca, C.; Picard, N.; Sparrow, B.; Abraham, E.M.; et al. The extent of forest in dryland biomes. Science 2017, 356, 635–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, J.F. Desertification. In Encyclopedia of Biodiversity; Levin, S.A., Ed.; Academic Press: San Diego, CA, USA, 2000; pp. 61–78. [Google Scholar]
- Dai, A. Drought under global warming: A review. Clim. Chang. 2011, 2, 45–65. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Coops, N.C.; Beck, P.S.A.; Chambers, J.Q.; Gangodagamage, C.; Hicke, J.A.; Huang, C.Y.; Kennedy, R.; Krofcheck, D.J.; Litvak, M.; et al. Global satellite monitoring of climate induced vegetation disturbances. Trends Plant Sci. 2015, 20, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Sheffield, J.; Wood, E.F.; Roderick, M.L. Little change in global drought over the past 60 years. Nature 2012, 491, 435–438. [Google Scholar] [CrossRef]
- Naithani, K.J.; Ewers, B.E.; Pendall, E. Sap flux-scaled transpiration and stomatal conductance response to soil and atmospheric drought in a semi-arid sagebrush ecosystem. J. Hydrol. 2012, 464–465, 176–185. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Li, D.; Si, J.H.; Zhang, X.Y.; Gao, Y.Y.; Wang, C.L.; Luo, H.; Qin, J.; Gao, G.L. Hydraulic characteristics of Populus euphratica in an arid environment. Forests 2019, 10, 407. [Google Scholar] [CrossRef] [Green Version]
- Si, J.H.; Feng, Q.; Wen, X.H.; Su, Y.H.; Xi, H.Y.; Chang, Z.Q. Major ion chemistry of groundwater in the extreme arid region northwest China. Environ. Geol. 2009, 57, 1079–1087. [Google Scholar] [CrossRef]
- Yu, T.F.; Feng, Q.; Si, J.H.; Pinkard, E.A. Coordination of stomatal control and stem water storage on plant water use in desert riparian trees. Trees 2019, 33, 787–801. [Google Scholar] [CrossRef]
- Yu, T.F.; Feng, Q.; Si, J.H.; Xi, H.Y.; O’Grady, A.P.; Pinkard, E.A. Responses of riparian forests to flood irrigation in the hyber-arid zone of NW China. Sci. Total Environ. 2019, 648, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.D.; Li, X.; Zhao, W.Z.; Xu, Z.M.; Feng, Q.; Xiao, S.C.; Xiao, H.L. Integrated study of the water–ecosystem–economy in the Heihe River Basin. Nat. Sci. Rev. 2014, 1, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.Z.; Li, X.; Yang, K.; Zhou, J. Assessing the impacts of an ecological water diversion project on water consumption through high-resolution estimations of actual evapotranspiration in the downstream regions of the Heihe River Basin, China. Agric. For. Meteorol. 2017, 249, 210–227. [Google Scholar] [CrossRef]
- Guo, Q.L.; Feng, Q.; Li, J.L. Environmental changes after ecological water conveyance in the lower reaches of Heihe River, northwest China. Environ. Geol. 2009, 58, 1387–1396. [Google Scholar] [CrossRef]
- Nian, Y.Y.; Li, X.; Zhou, J. Landscape changes of the Ejin delta in the Heihe River Basin in northwest China from 1930 to 2010. Int. J. Remote Sens. 2017, 38, 537–557. [Google Scholar] [CrossRef]
- Zhang, S.H.; Ye, Z.X.; Chen, Y.N.; Xu, Y.F. Vegetation responses to an ecological water conveyance project in the lower reaches of the Heihe River basin. Ecohydrology 2017, 10, e1866. [Google Scholar] [CrossRef]
- Chen, Y.P.; Chen, Y.N.; Li, W.H.; Xu, C.C. Characterization of photosynthesis of Populus euphratica grown in the arid region. Photosynthetica 2006, 44, 622–626. [Google Scholar] [CrossRef]
- Wang, H.Z.; Han, L.; Xu, Y.L.; Niu, J.L.; Yu, J. Effects of soil water gradient on photosynthetic characteristics and stress resistance of Populus pruinosa in the Tarim Basin, China. Acta Ecol. Sin. 2017, 37, 432–442. (In Chinese) [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.H.; Chen, Y.N.; Li, W.H.; Chen, Y.P. Photosynthesis of Populus euphratica in relation to groundwater tables and high temperature in arid environment, northwest China. Photosynthetica 2010, 48, 257–268. [Google Scholar] [CrossRef]
- Zheng, C.X.; Qiu, J.; Jiang, C.N.; Yue, N. Comparison of stomatal characteristics and photosynthesis of polymorphic Populus euphratica leaves. Front. For. China 2007, 2, 87–93. [Google Scholar] [CrossRef]
- Zhu, G.F.; Li, X.; Su, Y.H.; Lu, L.; Huang, C. Seasonal fluctuations and temperature dependence in photosynthetic parameters and stomatal conductance at the leaf scale of Populus euphratica Oliv. Tree Physiol. 2011, 31, 178–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.P.; Chen, Y.P.; Chen, Y.N.; Wang, R.Z.; Ren, Z.G. Impact of groundwater table on leaf hydraulic properties and drought vulnerability of Populus euphratica in the Northwest of China. Trees 2016, 30, 2029–2039. [Google Scholar] [CrossRef]
- Chen, Y.N.; Chen, Y.P.; Li, W.H.; Zhang, H.F. Response of the accumulation of proline in the bodies of Populus euphratica to the change of groundwater level at the lower reaches of Tarim River. Chin. Sci. Bull. 2003, 48, 1995–1999. [Google Scholar] [CrossRef]
- Kumagai, T.O.; Saitoh, T.M.; Sato, Y.; Morooka, T.; Manfroi, O.J.; Kuraji, K.; Suzuki, M. Transpiration, canopy conductance and the decoupling coefficient of a lowland mixed dipterocarp forest in Sarawak, Borneo: Dry spell effects. J. Hydrol. 2004, 287, 237–251. [Google Scholar] [CrossRef]
- Nicolás, E.; Barradas, V.L.; Ortuño, M.F.; Navarro, A.; Torrecillas, A.; Alarcón, J.J. Environmental and stomatal control of transpiration, canopy conductance and decoupling coefficient in young lemon trees under shading net. Environ. Exp. Bot. 2008, 63, 200–206. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Zhao, P.; McCarthy, H.R.; Zhao, X.H.; Niu, J.F.; Zhu, L.W.; Ni, G.Y.; Ouyang, L.; Huang, Y.Q. Influence of the decoupling degree on the estimation of canopy stomatal conductance for two broadleaf tree species. Agric. For. Meteorol. 2016, 221, 230–241. [Google Scholar] [CrossRef]
- Abdurahman, M.; Kurban, A.; Halik, U.; Ablekim, A.; Duan, H. Study on phenological characters of Populus euphratica Oliv. and its relation with the tree diameter. Vegetos 2013, 26, 88–92. [Google Scholar] [CrossRef]
- Jarvis, P.G.; McNaughton, K.G. Stomatal control of transpiration: Scaling up from leaf to region. Adv. Ecol. Res. 1986, 15, 1–49. [Google Scholar] [CrossRef]
- Berry, J.A.; Downton, W.J.S. Environmental regulation of photosynthesis. In Photosynthesis Vol. II; Govindjee, N.Y., Ed.; Academic Press: New York, NY, USA, 1982; pp. 263–343. [Google Scholar]
- Gao, G.L.; Feng, Q.; Zhang, X.Y.; Si, J.H.; Yu, T.F. An overview of stomatal and non-stomatal limitations to photosynthesis of plants. Arid. Zone Res. 2018, 35, 929–937. (in Chinese). [Google Scholar] [CrossRef]
- Ye, Z.P.; Xie, Z.L.; Duan, S.H.; An, T.; Zeng, G.H.; Kang, H.J. Stomatal and non-stomatal limitations of photosynthesis for Tetrastigma hemsleyanum under the condition of facility cultivation. Plant Physiol. J. 2020, 56, 41–48. (in Chinese). [Google Scholar] [CrossRef]
- Song, X.Y.; Zhou, G.S.; He, Q.J.; Zhou, H.L. Stomatal limitations to photosynthesis and their critical water conditions in different growth stages of maize under water stress. Agr. Water. Manag. 2020, 241, 106330. [Google Scholar] [CrossRef]
- Kicheva, M.I.; Tsonev, T.T.; Popova, L.P. Stomatal and nonstomatal limitations to photosynthesis in two wheat cultivars subjected to water stress. Photosynthetica 1994, 30, 107–116. [Google Scholar] [CrossRef]
- Ramanjulu, S.; Sreenivasulu, N.; Sudhakar, C. Effect of water stress on photosynthesis in two mulberry genotypes with different drought tolerance. Photosynthetica 1998, 35, 279–283. [Google Scholar] [CrossRef]
- Burgess, S.S.O.; Adams, M.A.; Turner, N.C.; Beverly, C.R.; Ong, C.K.; Khan, A.A.H.; Bleby, T.M.; Notes, A. An improved heat pulse method to measure low and reverse rates of sap flow in woody plants. Tree Physiol. 2001, 21, 589–598. [Google Scholar] [CrossRef]
- Granier, A.; Loustau, D.; Bréda, N. A generic model of forest canopy conductance dependent on climate, soil water availability and leaf area index. Ann. For. Sci. 2000, 57, 755–765. [Google Scholar] [CrossRef]
- Xu, D.Q. Some problems in stomatal limitation analysis of photosynthesis. Plant Physiol. Commun. 1997, 33, 241–244. (In Chinese) [Google Scholar]
- Gao, Z.J.; Xu, B.C.; Wang, J.; Luo, L.J.; Li, S. Diurnal and seasonal variations in photosynthetic characteristics of switchgrass in semiarid region on the Loess Plateau of China. Photosynthetica 2015, 53, 489–498. [Google Scholar] [CrossRef]
- Yang, Z.S.; Zhang, Q.; Hao, X.C. Stomatal or non-stomatal limitation of photosynthesis of spring wheat flag leaf at late growth stages under natural conditions in semiarid rainfed regions. Chin. J. Eco-Agric. 2015, 23, 174–182. (In Chinese) [Google Scholar]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Raschke, K.; Resemann, A. The midday depression of CO2 assimilation in leaves of Arbutus unedo L.: Diurnal changes in photosynthetic capacity related to changes in temperature and humidity. Planta 1986, 168, 546. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, G.; Massacci, A.; Gabrielli, R. Drought effects on photosynthesis and fluorescence in hard wheat cultivars grown in the field. Physiol. Plant 1988, 74, 385–390. [Google Scholar] [CrossRef]
- Tenhunen, J.D.; Lange, O.L.; Gebel, J.; Beyschlag, W.; Weber, J.A. Changes in photosynthetic capacity, carboxylation efficiency, and CO2 compensation point associated with midday stomatal closure and midday depression of net CO2 exchange of leaves of Quercus suber. Planta 1984, 162, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.X.; Dai, J.Y.; Lin, Y. The photosynthetic stomatal and nonstomatal limitation of plant leaves under water stress. Plant Physiol. Commun. 1995, 31, 293. (In Chinese) [Google Scholar]
- Grassi, G.; Magnani, F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Cescatti, A.; Rodeghiero, M.; Tosens, T. Complex adjustments of photosynthetic potentials and internal diffusion conductance to current and previous light availabilities and leaf age in Mediterranean evergreen species Quercusilex. Plant Cell Environ. 2005, 29, 1159–1178. [Google Scholar] [CrossRef] [Green Version]
- Warren, C.R. Stand aside stomata, another actor deserves centre stage: The forgotten role of the internal conductance to CO2 transfer. J. Exp. Bot. 2008, 59, 1475–1487. [Google Scholar] [CrossRef] [Green Version]
- Nobel, P.S. Internal leaf area and cellular CO2 resistance: Photosynthetic implications of variations with growth conditions and plant species. Physiol. Plant. 1977, 40, 137–144. [Google Scholar] [CrossRef]
- Flexas, J.; Ribas-Carbó, M.; Diaz-Espejo, A.; Galmés, J.; Medrano, H. Mesophyll conductance to CO2: Current knowledge and future prospects. Plant Cell Environ. 2008, 31, 602–621. [Google Scholar] [CrossRef]
- Scartazza, A.; Lauteri, M.; Guido, M.C.; Brugnoli, E. Carbon isotope discrimination in leaf and stem sugars, water-use efficiency and mesophyll conductance during different developmental stages in rice subjected to drought. Aust. J. Plant Physiol. 1998, 25, 489–498. [Google Scholar] [CrossRef]
- Delfine, S.; Loreto, F.; Alvino, A. Drought-stress effects on physiology, growth and biomass production of rainfed and irrigated bell pepper plants in the Mediterranean region. J. Am. Soc. Hortic. Sci. 2001, 126, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Monti, A.; Brugnoli, E.; Scartazza, A.; Amaducci, M.T. The effect of transient and continuous drought on yield, photosynthesis and carbon isotope discrimination in sugar beet (Beta vulgaris L.). J. Exp. Bot. 2006, 57, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bongi, G.; Loreto, F. Gas-exchange properties of salt-stressed Olive (Olea europea L.) Leaves. Plant Physiol. 1989, 90, 1408–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gries, D.; Zeng, F.J.; Arndt, S.K.; Bruelheide, H.; Thomas, F.M.; Runge, M. Growth and water relations of Tamarix ramosissima and Populus euphratica on Taklamakan desert dunes in relation to depth to a permanent water table. Plant Cell Environ. 2003, 26, 725–736. [Google Scholar] [CrossRef]
- Zeng, F.; Bleby, T.M.; Landman, P.A.; Adams, M.A.; Arndt, S.K. Water and nutrient dynamics in surface roots and soils are not modified by short-term flooding of phreatophytic plants in a hyperarid desert. Plant Soil 2006, 279, 129–139. [Google Scholar] [CrossRef]
- Li, W.; Zhou, H.; Fu, A.; Chen, Y. Ecological response and hydrological mechanism of desert riparian forest in inland river, northwest of China. Ecohydrology 2013, 6, 949–955. [Google Scholar] [CrossRef]
- Lang, P.; Ahlborn, J.; Schäfer, P.; Wommelsdorf, T.; Jeschke, M.; Zhang, X.M.; Thomas, F.M. Growth and water use of Populus euphratica trees and stands with different water supply along the Tarim River, NW China. For. Ecol. Manag. 2016, 380, 139–148. [Google Scholar] [CrossRef]
- Hao, X.M.; Li, W.H.; Huang, X.; Zhu, C.G.; Ma, J.X. Assessment of the groundwater threshold of desert riparian forest vegetation along the middle and lower reaches of the Tarim River, China. Hydrol. Process. 2010, 24, 178–186. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Y.; Yu, J.; Fu, G.; Ao, F. Vegetation dynamics induced by groundwater fluctuations in the lower Heihe River Basin, northwestern China. J. Plant Ecol. 2011, 4, 77–90. [Google Scholar] [CrossRef]
- Wang, G.X.; Cheng, G.D. Water demand of eco-system and estimate method in arid inland river basins. J. Desert Res. 2002, 22, 129–134. (In Chinese) [Google Scholar]
- Jiang, X.H.; Liu, C.M. The response of vegetation to water transport in the lower reaches of the Heihe River. Acta Geogr. Sin. 2009, 64, 791–797. (in Chinese). [Google Scholar] [CrossRef]
- Tissue, D.T.; Griffin, K.L.; Turnbull, M.H.; Whitehead, D. Stomatal and non-stomatal limitations to photosynthesis in four tree species in a temperate rainforest dominated by Dacrydium cupressinum in New Zealand. Tree Physiol. 2005, 25, 447–456. [Google Scholar] [CrossRef] [PubMed]








| Tree Number | Height (m) | DBH (cm) | As (cm2) |
|---|---|---|---|
| 1 | 10.6 | 23.9 | 225.6 |
| 2 | 10.4 | 21.7 | 206.4 |
| 3 | 10.3 | 30.4 | 361.2 |
| Sig. (2-tailed) | 0.105 | 0.623 | 0.718 |
| Year | PAR | VPD | Ta | hs | |
|---|---|---|---|---|---|
| Pn | 2013 | 0.820 ** | 0.463 ** | 0.489 ** | −0.180 |
| 2014 | 0.709 ** | 0.470 ** | 0.464 ** | −0.370 ** |
| Objectives | Criteria | Source of Data |
|---|---|---|
| Stomatal limitation | The decreasing range of Ci is much lower than that of Pn | [53] |
| Non-stomatal limitation | Ci remains almost constant | [54,55] |
| Stomatal and non-stomatal limitation | Comparing the values of Ls and Ci/gs | [56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, G.; Feng, Q.; Liu, X.; Yu, T.; Wang, R. The Photosynthesis of Populus euphratica Oliv. Is Not Limited by Drought Stress in the Hyper-Arid Zone of Northwest China. Forests 2022, 13, 2096. https://doi.org/10.3390/f13122096
Gao G, Feng Q, Liu X, Yu T, Wang R. The Photosynthesis of Populus euphratica Oliv. Is Not Limited by Drought Stress in the Hyper-Arid Zone of Northwest China. Forests. 2022; 13(12):2096. https://doi.org/10.3390/f13122096
Chicago/Turabian StyleGao, Guanlong, Qi Feng, Xiande Liu, Tengfei Yu, and Rongxin Wang. 2022. "The Photosynthesis of Populus euphratica Oliv. Is Not Limited by Drought Stress in the Hyper-Arid Zone of Northwest China" Forests 13, no. 12: 2096. https://doi.org/10.3390/f13122096