
Further Test of Pneumatic Method in Constructing Vulnerability Curves Using Six Tree Species with Contrasting Xylem Anatomy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Conduit Length and Diameter
2.3. Volume Estimation of Open Vessels Reservoir
2.4. Vulnerability Curves via Hydraulic Methods
2.5. Vulnerability Curves via Pneumatic Method
2.6. Leaf Water Potential at Turgor Loss Point
2.7. Native Xylem Pressure and Embolism
2.8. Data Synthesis
2.9. Statistical Analyses
3. Results
3.1. Xylem Anatomy and the Volume of Open Vessels Reservoir
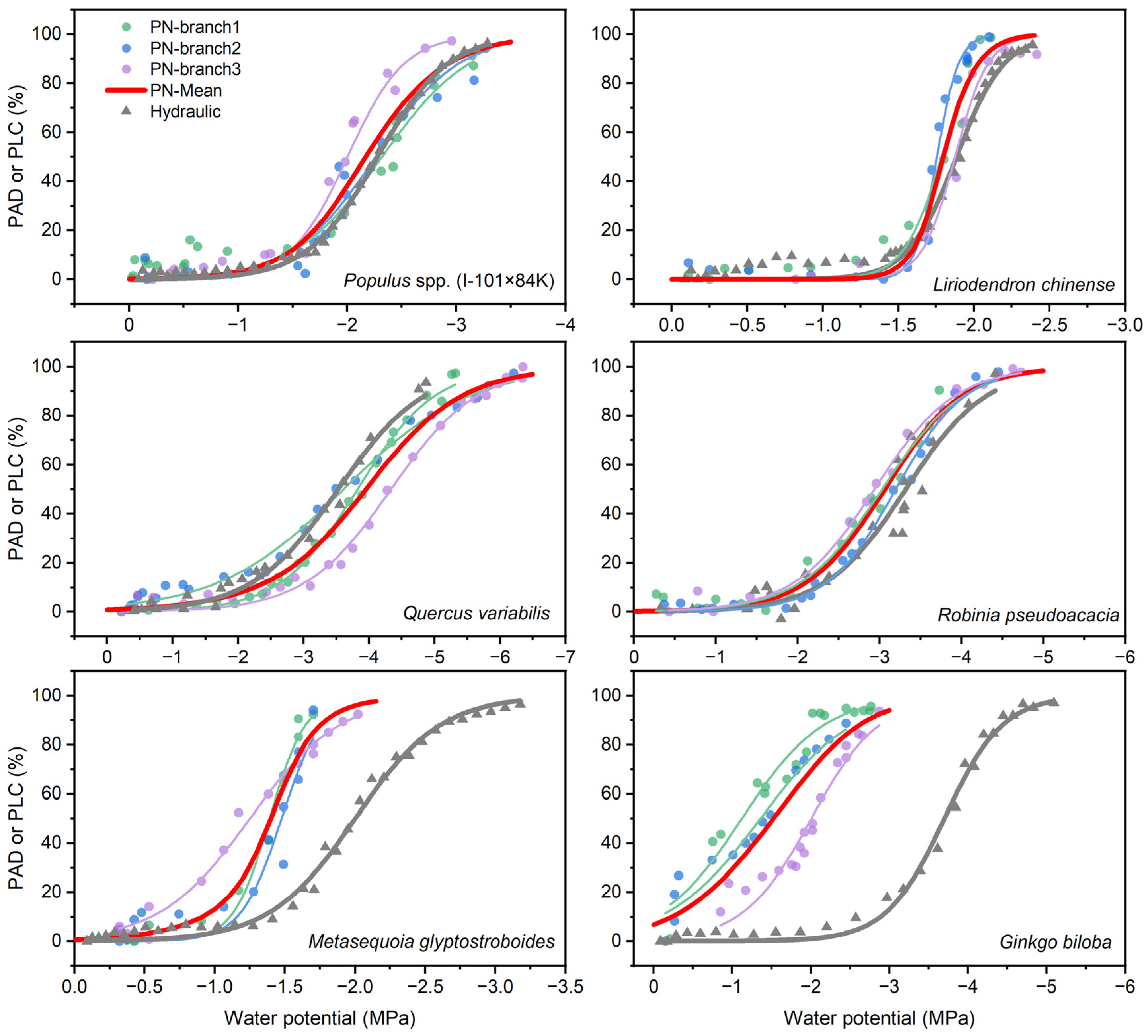
3.2. VCs Constructed by Pneumatic and Hydraulic Methods
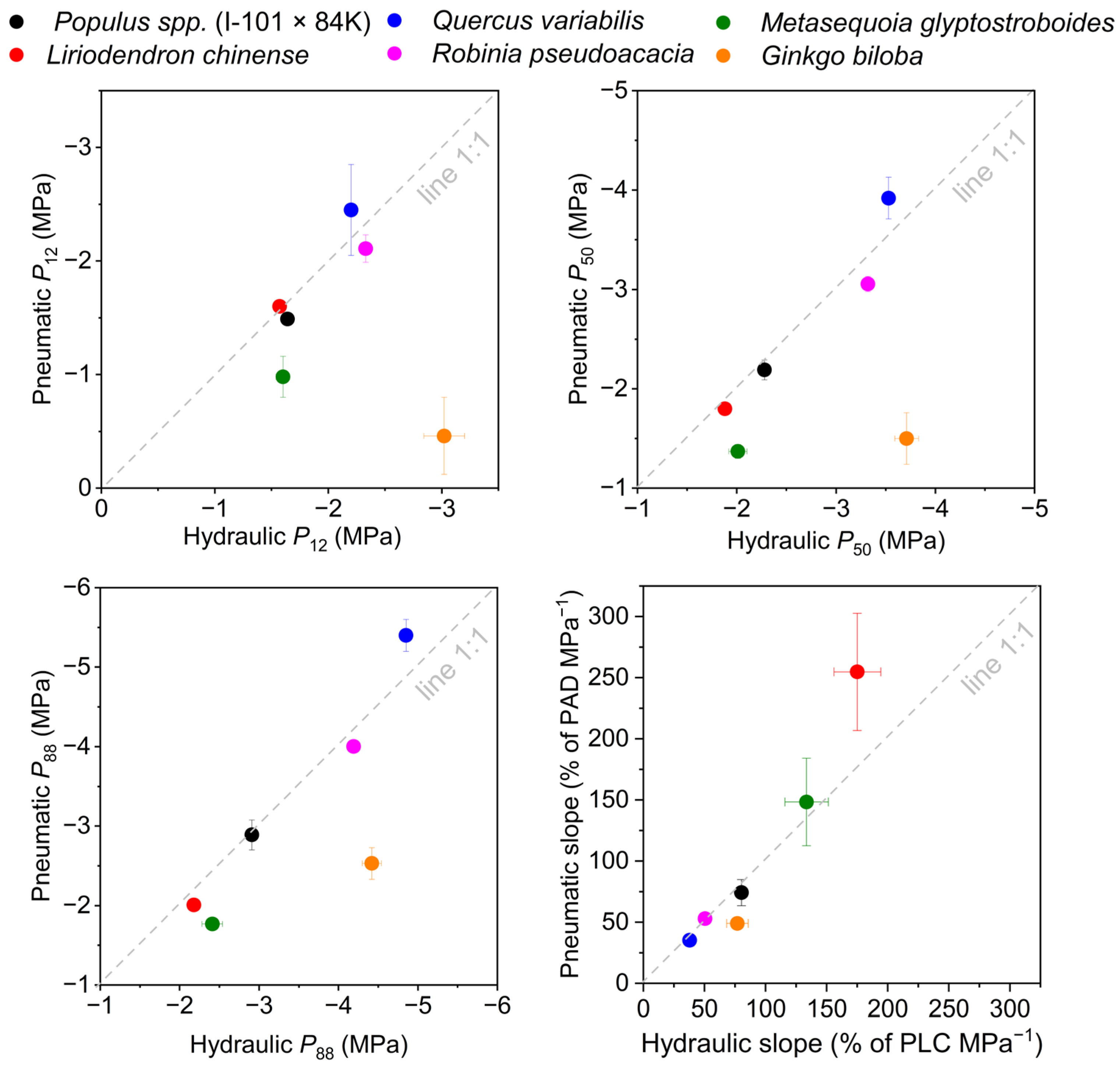
3.3. Reliability Test via Water Relations
4. Discussion
4.1. The Reliability of Hydraulic Methods
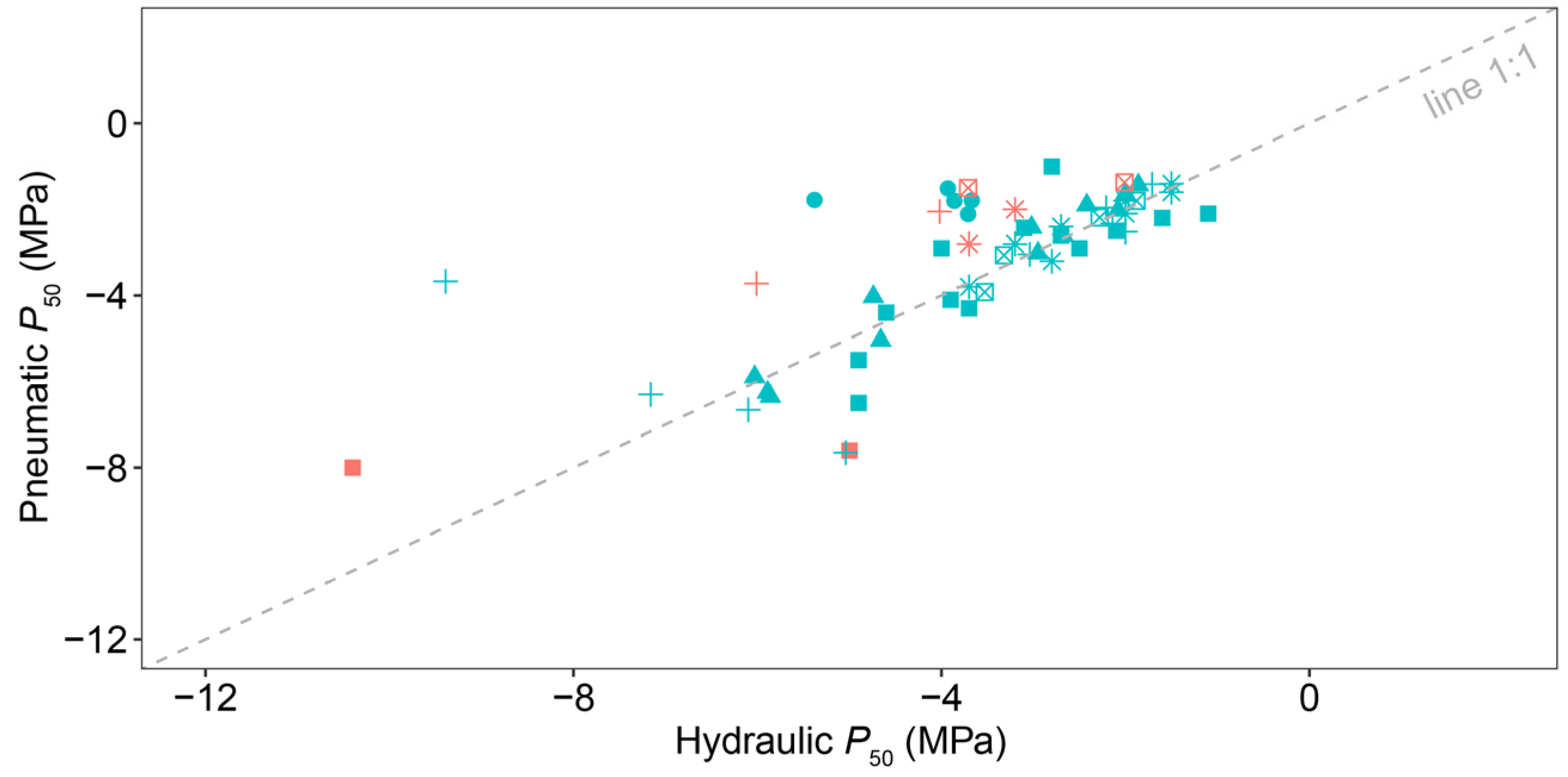
4.2. The Consistency between Pneumatic and Hydraulic Methods
4.3. The Impact of Open Vessels Volume
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; Lopez, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Powers, J.; Cochard, H.; Choat, B. Hanging by a thread? Forests and drought. Science 2020, 368, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Rowland, L.; da Costa, A.C.; Galbraith, D.R.; Oliveira, R.S.; Binks, O.J.; Oliveira, A.A.; Pullen, A.M.; Doughty, C.E.; Metcalfe, D.B.; Vasconcelos, S.S.; et al. Death from drought in tropical forests is triggered by hydraulics not carbon starvation. Nature 2015, 528, 119–122. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2002; pp. i–xiv, 1–283. [Google Scholar]
- Cochard, H.; Badel, E.; Herbette, S.; Delzon, S.; Choat, B.; Jansen, S. Methods for measuring plant vulnerability to cavitation: A critical review. J. Exp. Bot. 2013, 64, 4779–4791. [Google Scholar] [CrossRef]
- Sperry, J.S.; Donnelly, J.R.; Tyree, M.T. A method for measuring hydraulic conductivity and embolism in xylem. Plant Cell Environ. 1988, 11, 35–40. [Google Scholar] [CrossRef]
- Cochard, H.; Cruiziat, P.; Tyree, M.T. Use of Positive Pressures to Establish Vulnerability Curves-Further Support for the Air-Seeding Hypothesis and Implications for Pressure-Volume Analysis. Plant Physiol. 1992, 100, 205–209. [Google Scholar] [CrossRef]
- Pockman, W.T.; Sperry, J.S.; Oleary, J.W. Sustained and Significant Negative Water-Pressure in Xylem. Nature 1995, 378, 715–716. [Google Scholar] [CrossRef]
- Alder, N.N.; Pockman, W.T.; Sperry, J.S.; Nuismer, S. Use of centrifugal force in the study of xylem cavitation. J. Exp. Bot. 1997, 48, 665–674. [Google Scholar] [CrossRef]
- Cochard, H. A technique for measuring xylem hydraulic conductance under high negative pressures. Plant Cell Environ. 2002, 25, 815–819. [Google Scholar] [CrossRef] [Green Version]
- Cochard, H.; Damour, G.; Bodet, C.; Tharwat, I.; Poirier, M.; Améglio, T. Evaluation of a new centrifuge technique for rapid generation of xylem vulnerability curves. Physiol. Plant. 2005, 124, 410–418. [Google Scholar] [CrossRef]
- Li, Y.; Sperry, J.S.; Taneda, H.; Bush, S.E.; Hacke, U.G. Evaluation of centrifugal methods for measuring xylem cavitation in conifers, diffuse- and ring-porous angiosperms. New Phytol. 2008, 177, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; Schuldt, B.; Choat, B. Current controversies and challenges in applying plant hydraulic techniques. New Phytol. 2015, 205, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, J.K.; Huggett, B.A.; Tofte, A.N.; Rockwell, F.E.; Holbrook, N.M. Cutting xylem under tension or supersaturated with gas can generate PLC and the appearance of rapid recovery from embolism. Plant Cell Environ. 2013, 36, 1938–1949. [Google Scholar] [CrossRef] [PubMed]
- Trifilo, P.; Raimondo, F.; Lo Gullo, M.A.; Barbera, P.M.; Salleo, S.; Nardini, A. Relax and refill: Xylem rehydration prior to hydraulic measurements favours embolism repair in stems and generates artificially low PLC values. Plant Cell Environ. 2014, 37, 2491–2499. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Jansen, S.; Choat, B.; McElrone, A.J.; Cochard, H.; Brodribb, T.J.; Badel, E.; Burlett, R.; Bouche, P.S.; Brodersen, C.R.; et al. Direct x-ray microtomography observation confirms the induction of embolism upon xylem cutting under tension. Plant Physiol. 2015, 167, 40–43. [Google Scholar] [CrossRef]
- Ennajeh, M.; Simoes, F.; Khemira, H.; Cochard, H. How reliable is the double-ended pressure sleeve technique for assessing xylem vulnerability to cavitation in woody angiosperms? Physiol. Plant. 2011, 142, 205–210. [Google Scholar] [CrossRef]
- Yin, P.; Cai, J. New possible mechanisms of embolism formation when measuring vulnerability curves by air injection in a pressure sleeve. Plant Cell Environ. 2018, 41, 1361–1368. [Google Scholar] [CrossRef]
- Cochard, H.; Herbette, S.; Barigah, T.; Badel, E.; Ennajeh, M.; Vilagrosa, A. Does sample length influence the shape of xylem embolism vulnerability curves? A test with the Cavitron spinning technique. Plant Cell Environ. 2010, 33, 1543–1552. [Google Scholar] [CrossRef]
- McElrone, A.J.; Brodersen, C.R.; Alsina, M.M.; Drayton, W.M.; Matthews, M.A.; Shackel, K.A.; Wada, H.; Zufferey, V.; Choat, B. Centrifuge technique consistently overestimates vulnerability to water stress-induced cavitation in grapevines as confirmed with high-resolution computed tomography. New Phytol. 2012, 196, 661–665. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, L.; Zhang, S.; Cai, J.; Tyree, M.T. Water relations of Robinia pseudoacacia L.: Do vessels cavitate and refill diurnally or are R-shaped curves invalid in Robinia? Plant Cell Environ. 2014, 37, 2667–2678. [Google Scholar] [CrossRef]
- Zhao, H.; Jiang, Z.; Ma, J.; Cai, J. What causes the differences in cavitation resistance of two shrubs? Wood anatomical explanations and reliability testing of vulnerability curves. Physiol. Plant. 2020, 169, 156–168. [Google Scholar] [CrossRef]
- Pereira, L.; Bittencourt, P.R.L.; Oliveira, R.S.; Junior, M.B.M.; Barros, F.V.; Ribeiro, R.V.; Mazzafera, P. Plant pneumatics: Stem air flow is related to embolism-new perspectives on methods in plant hydraulics. New Phytol. 2016, 211, 357–370. [Google Scholar] [CrossRef]
- Pereira, L.; Bittencourt, P.R.L.; Pacheco, V.S.; Miranda, M.T.; Zhang, Y.; Oliveira, R.S.; Groenendijk, P.; Machado, E.C.; Tyree, M.T.; Jansen, S.; et al. The Pneumatron: An automated pneumatic apparatus for estimating xylem vulnerability to embolism at high temporal resolution. Plant Cell Environ. 2020, 43, 131–142. [Google Scholar] [CrossRef]
- Trabi, C.L.; Pereira, L.; Guan, X.; Miranda, M.T.; Bittencourt, P.R.L.; Oliveira, R.S.; Ribeiro, R.V.; Jansen, S. A User Manual to Measure Gas Diffusion Kinetics in Plants: Pneumatron Construction, Operation, and Data Analysis. Front. Plant Sci. 2021, 12, 633595. [Google Scholar] [CrossRef]
- Sergent, A.S.; Varela, S.A.; Barigah, T.S.; Badel, E.; Cochard, H.; Dalla-Salda, G.; Delzon, S.; Fernández, M.E.; Guillemot, J.; Gyenge, J.; et al. A comparison of five methods to assess embolism resistance in trees. For. Ecol. Manag. 2020, 468, 118175. [Google Scholar] [CrossRef]
- Zhang, Y.; Lamarque, L.J.; Torres-Ruiz, J.M.; Schuldt, B.; Karimi, Z.; Li, S.; Qin, D.W.; Bittencourt, P.; Burlett, R.; Cao, K.F.; et al. Testing the plant pneumatic method to estimate xylem embolism resistance in stems of temperate trees. Tree Physiol. 2018, 38, 1016–1025. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Maenpuen, P.; Zhang, Y.-J.; Barai, K.; Katabuchi, M.; Gao, H.; Kaewkamol, S.; Tao, L.-B.; Zhang, J.-L. Quantifying vulnerability to embolism in tropical trees and lianas using five methods: Can discrepancies be explained by xylem structural traits? New Phytol. 2021, 229, 805–819. [Google Scholar] [CrossRef]
- Paligi, S.S.; Link, R.M.; Isasa, E.; Bittencourt, P.; Cabral, J.S.; Jansen, S.; Oliveira, R.S.; Pereira, L.; Schuldt, B. Accuracy of the pneumatic method for estimating xylem vulnerability to embolism in temperate diffuse-porous tree species. BioRxiv 2021. [Google Scholar] [CrossRef]
- Guan, X.; Pereira, L.; McAdam, S.A.M.; Cao, K.F.; Jansen, S. No gas source, no problem: Proximity to pre-existing embolism and segmentation affect embolism spreading in angiosperm xylem by gas diffusion. Plant Cell Environ. 2021, 44, 1329–1345. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, Y.; Oya, T.; Marcati, C.R.; Pereira, L.; Jansen, S. Root xylem in three woody angiosperm species is not more vulnerable to embolism than stem xylem. Plant Soil 2020, 450, 479–495. [Google Scholar] [CrossRef]
- Martin-StPaul, N.; Delzon, S.; Cochard, H. Plant resistance to drought depends on timely stomatal closure. Ecol. Lett. 2017, 20, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, U.; Windt, C.W.; Ponomarenko, A.; Zhang, Y.-J.; Gersony, J.; Rockwell, F.E.; Holbrook, N.M. Stomatal Closure, Basal Leaf Embolism, and Shedding Protect the Hydraulic Integrity of Grape Stems. Plant Physiol. 2017, 174, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Holbrook, N.M. Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol. 2003, 132, 2166–2173. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, J.K.; Sperry, J.S.; Hacke, U.G.; Hoang, N. Inter-vessel pitting and cavitation in woody Rosaceae and other vesselled plants: A basis for a safety versus efficiency trade-off in xylem transport. Plant Cell Environ. 2005, 28, 800–812. [Google Scholar] [CrossRef]
- Franklin, G.L. Preparation of thin sections of synthetic resins and wood-resin composites, and a new macerating method for wood. Nature 1945, 155, 51. [Google Scholar] [CrossRef]
- Tyree, M.T.; Hammel, H.T. The Measurement of the Turgor Pressure and the Water Relations of Plants by the Pressure-bomb Technique. J. Exp. Bot. 1972, 23, 267–282. [Google Scholar] [CrossRef]
- Lopez, R.; Nolf, M.; Duursma, R.A.; Badel, E.; Flavel, R.J.; Cochard, H.; Choat, B. Mitigating the open vessel artefact in centrifuge-based measurement of embolism resistance. Tree Physiol. 2019, 39, 143–155. [Google Scholar] [CrossRef]
- Cai, J.; Hacke, U.; Zhang, S.; Tyree, M.T. What happens when stems are embolized in a centrifuge? Testing the cavitron theory. Physiol. Plant. 2010, 140, 311–320. [Google Scholar] [CrossRef]
- De Baerdemaeker, N.J.F.; Arachchige, K.N.R.; Zinkernagel, J.; Van den Bulcke, J.; Van Acker, J.; Schenk, H.J.; Steppe, K. The stability enigma of hydraulic vulnerability curves: Addressing the link between hydraulic conductivity and drought-induced embolism. Tree Physiol. 2019, 39, 1646–1664. [Google Scholar] [CrossRef]
- Cochard, H.; Petit, G.; Torres-Ruiz, J.M.; Sylvain, D. XIM4 meeting report, Sept. 25-27 2019, Padua (Italy)The hydraulic community has taken a step forward towards non-invasive measurements. For. Ecol. Manag. 2019, 6, 118175. [Google Scholar]
- Cochard, H.; Breda, N.; Granier, A. Whole tree hydraulic conductance and water loss regulation in Quercus during drought: Evidence for stomatal control of embolism? Ann. For. Sci. 1996, 53, 197–206. [Google Scholar] [CrossRef]
- Pereira, L.; Bittencourt, P.R.L.; Rowland, L.; Brum, M.; Miranda, M.T.; Pacheco, V.S.; Oliveira, R.S.; Machado, E.C.; Jansen, S.; Ribeiro, R.V. Using the Pneumatic method to estimate embolism resistance in species with long vessels: A commentary on the article “A comparison of five methods to assess embolism resistance in trees”. For. Ecol. Manag. 2021, 479, 118547. [Google Scholar] [CrossRef]
- Yang, D.; Pereira, L.; Peng, G.; Ribeiro, R.V.; Kaack, L.; Jansen, S.; Tyree, M.T. A Unit Pipe Pneumatic model to simulate gas kinetics during measurements of embolism in excised angiosperm xylem. Tree Physiol. 2021, 43, 88–101. [Google Scholar] [CrossRef]
- Chen, Y.J.; Maenpuen, P.; Zhang, J.L.; Zhang, Y.J. Remaining uncertainties in the Pneumatic method. New Phytol. 2023, 237, 384–391. [Google Scholar] [CrossRef]
- Brum, M.; Pereira, L.; Ribeiro, R.V.; Jansen, S.; Bittencourt, P.R.L.; Oliveira, R.S.; Saleska, S.R. Reconciling discrepancies in measurements of vulnerability to xylem embolism with the pneumatic method: A comment on Chen et al. (2021) ‘Quantifying vulnerability to embolism in tropical trees and lianas using five methods: Can discrepancies be explained by xylem structural traits?’: A comment on Chen et al. (2021) ‘Quantifying vulnerability to embolism in tropical trees and lianas using five methods: Can discrepancies be explained by xylem structural traits?’. New Phytol. 2023, 237, 374–383. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Wood Type | Lmean (cm) | Dmean (μm) | VD (n mm−2) |
|---|---|---|---|---|
| Populus. spp. (I-101 × 84K) | Diffuse-porous | 4.57 ± 0.45 | 31.87 ± 1.22 | 187 ± 13 |
| Liriodendron chinense | Diffuse-porous | 3.74 ± 0.24 | 30.04 ± 0.52 | 351 ± 8 |
| Quercus variabilis | Ring-porous | 38.08 ± 2.80 | 47.86 ± 2.38 | 90 ± 6 |
| Robinia pseudoacacia | Ring-porous | 15.59 ± 0.65 | 56.09 ± 1.27 | 45 ± 6 |
| Metasequoia glyptostroboides | Non-porous | 0.15 ± 0.01 | 13.61 ± 0.46 | 2260 ± 77 |
| Ginkgo biloba | Non-porous | 0.19 ± 0.01 | 16.23 ± 0.33 | 1585 ± 62 |
| Species | Methods | P12 (MPa) | P50 (MPa) | P88 (MPa) | Slope (% of PLC MPa−1) |
|---|---|---|---|---|---|
| Populus. spp. (I-101 × 84K) | Hydraulic | −1.64 ± 0.06 | −2.28 ± 0.07 | −2.91 ± 0.03 | 80.29 ± 4.72 |
| Pneumatic | −1.49 ± 0.01 | −2.19 ± 0.10 | −2.89 ± 0.19 | 74.23 ± 10.69 | |
| Liriodendron chinense | Hydraulic | −1.57 ± 0.05 | −1.88 ± 0.03 | −2.18 ± 0.04 | 175.04 ± 19.18 |
| Pneumatic | −1.60 ± 0.03 | −1.80 ± 0.04 | −2.01 ± 0.06 | 254.80 ± 47.96 | |
| Quercus variabilis | Hydraulic | −2.2 | −3.53 | −4.85 | 37.77 |
| Pneumatic | −2.45 ± 0.4 | −3.92 ± 0.21 | −5.4 ± 0.20 | 35.38 ± 4.85 | |
| Robinia pseudoacacia | Hydraulic | −2.33 | −3.32 | −4.19 | 50.49 |
| Pneumatic | −2.11 ± 0.12 | −3.06 ± 0.08 | −4.00 ± 0.04 | 53.09 ± 2.86 | |
| Metasequoia glyptostroboides | Hydraulic | −1.69 ± 0.06 | −2.01 ± 0.09 | −2.41 ± 0.13 | 133.53 ± 17.78 |
| Pneumatic | −0.98 ± 0.18 * | −1.37 ± 0.07 * | −1.77 ± 0.07 * | 148.34 ± 35.76 | |
| Ginkgo biloba | Hydraulic | −3.02 ± 0.18 | −3.71 ± 0.12 | −4.42 ± 0.12 | 76.90 ± 8.79 |
| Pneumatic | −0.46 ± 0.34 * | −1.50 ± 0.26 * | −2.53 ± 0.20 * | 49.18 ± 4.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Li, Y.; Liao, S.; Jiang, Z.; Cai, J. Further Test of Pneumatic Method in Constructing Vulnerability Curves Using Six Tree Species with Contrasting Xylem Anatomy. Forests 2023, 14, 293. https://doi.org/10.3390/f14020293
Zhao H, Li Y, Liao S, Jiang Z, Cai J. Further Test of Pneumatic Method in Constructing Vulnerability Curves Using Six Tree Species with Contrasting Xylem Anatomy. Forests. 2023; 14(2):293. https://doi.org/10.3390/f14020293
Chicago/Turabian StyleZhao, Han, Yueyang Li, Suhui Liao, Zaimin Jiang, and Jing Cai. 2023. "Further Test of Pneumatic Method in Constructing Vulnerability Curves Using Six Tree Species with Contrasting Xylem Anatomy" Forests 14, no. 2: 293. https://doi.org/10.3390/f14020293