
Toward the Genetic Improvement of Drought Tolerance in Conifers: An Integrated Approach


1
Research and Innovation Centre, Fondazione Edmund Mach (FEM), 38098 San Michele all’Adige, Trento, Italy
2
EFI Project Centre on Mountain Forests (MOUNTFOR), 38098 San Michele all’Adige, Trento, Italy
*
Author to whom correspondence should be addressed.
Forests 2022, 13(12), 2016; https://doi.org/10.3390/f13122016
Submission received: 26 October 2022
/
Revised: 25 November 2022
/
Accepted: 27 November 2022
/
Published: 29 November 2022
(This article belongs to the Special Issue Tree Genetics: Molecular and Functional Characterization of Genes)
Abstract
:The constant rise in the global temperature and unpredictable shifts in precipitation patterns are two of the main effects of climate change. Therefore, there is an increasing amount of interest in the identification of tree species, provenances and genotypes capable of withstanding more arid conditions and tolerating drought stress. In the present review, we focus our attention on generally more susceptible conifers and describe the different strategies that plants adopt to respond to drought stress. We describe the main approaches taken in studies of conifer adaptations to low water availability, the advantages and limitations of each, and the main results obtained with each of these approaches in the recent years. Then we discuss how the increasing amount of morphological, physiological and genetic data may find practical applications in forest management, and in particular in next-generation breeding programs. Finally, we provide some recommendations for future research. In particular, we suggest extending future studies to a broader selection of species and genera, increasing the number of studies on adult plants, in particular those on gene expression, and distinguishing between the different types of drought stress that a tree can withstand during its life cycle. The integration of data coming from different disciplines and approaches will be a key factor to increasing our knowledge about a trait as complex as drought resistance.
Keywords:
gymnosperms; molecular; next-generation breeding; water stress; genetics; genomic; drought resistance; Picea; Abies; Pinus1. Introduction
According to the FAO’s Global Forest Resources Assessment 2020 (https://www.fao.org/documents/card/en/c/ca8753en (accessed on 2 July 2022)), 4.06 billion hectares of land are covered by forests worldwide, corresponding to over 30% of the total land area. Over the last decades, climate change has increased the risk of drought stress (DS) in many regions, mainly via increased temperatures, altered precipitation patterns and faster snow melt [1,2]. The effects of droughts on forest plants vary, from reduced growth in cases of moderate droughts to mass mortality if severe droughts occur [3,4]. Global simulations predict widespread massive tree mortality under the projected rise in global temperatures and extremes that accompanies drought [5]. Therefore, it is of primary importance to understand plant response and adaptation mechanisms to drought in order to properly manage tree populations and select those individuals or provenances that show a higher resistance level. Drought resistance is a very complex trait with both environmental and genetic components, and different populations of the same species may not respond equally to a given climate [6,7]. Moreover, several distinct aspects can be important when drought tolerance (DT) is concerned. As an example, in regards to wood production, the capability of the plant to grow in low-water conditions might be the most important trait to consider, while for a natural forest, adaptations to highly variable water availability might be more favorable. In all cases, the main target of research studies is to understand what traits are more important to achieving drought resistance and to find the relationship between phenotypes and genotypes [8,9].
Conifers originated more than 300 million years ago and currently dominate many temperate and boreal forests. They have very large genomes (18 to 35 Gb), showing a very different structure and composition when compared with those of angiosperm genomes. Despite an apparently conserved genome structure, conifers demonstrate some competitive capacity, as different taxa are adapted to a wide variety of environmental conditions [10]. Even though angiosperms and gymnosperms share some general principles regarding drought resistance, there are also significant differences. Some studies suggest that angiosperm tree species, in general, tend to be less sensitive to drought than gymnosperm species [11,12]. At first glance, one of the fundamental differences in water transport is the size and function of conifer tracheids and angiosperm vessels [13]. In fact, even though conifers have greater stem hydraulic safety than angiosperms, during drought events, conifers experience more frequent embolisms than angiosperms in distal tissues [14]. Embolism repair is likely driven by sugars that come from nearby parenchyma cells, and conifers have few carbohydrates or parenchyma in their xylems compared to angiosperms. So, even though conifers tend to experience embolism more frequently in their leaves and roots than angiosperms, these organs may act as hydraulic circuit breakers to prevent stem embolism in conifers [15]. Recent ecophysiological studies have suggested the existence of contrasting strategies in angiosperms and gymnosperms regarding traits related to DT, such as the accumulation of non-structural carbohydrates (NSCs) [16,17,18]. Other studies reported that a reduction in precipitation is predicted to have negative effects on the growth of both angiosperms and gymnosperms, while increased temperatures may result in a performance disadvantage for conifers [19]. Gymnosperms, due to their long life cycle (most trees older than 1000 years are gymnosperms), have a high chance of experiencing numerous drought events during their lives [20]. Moreover, even drought-resistant evergreen gymnosperms growing in Mediterranean forests might be strongly impacted if drought periods become more frequent and severe [21]. The present review is focused on conifers and will not describe in detail all the numerous differences between gymnosperms and angiosperms. Such differences have been extensively reviewed elsewhere [22,23,24,25].
In the next paragraphs, we summarize the main approaches that have been used over the last few years to study a complex trait, drought resistance, in conifers. First of all, we define the main aspects of the trait and the different strategies that trees can adopt to increase their DT. Then we discuss some of the more frequently used techniques and review the main findings that have been obtained so far. Finally, we describe how the results can be applied in forest management, tree improvement programs and future research.
2. Plant Strategies to Cope with Drought
Intuitively, a very simple definition of DT is the ability of a plant to survive a prolonged period of water shortage. This can be true in particular environments, such as deserts or semi-arid regions where water availability is low most of the time. For the analysis of DT in more temperate areas, different climatic drivers and levels of variability among tree species should be taken into account [26]. Among other factors, trees’ growth and in particular trees’ short-term responses to extreme drought events are two of the most frequently studied [27]. Plants that are adapted to low water levels can also show a very low growth rate in favorable conditions as a consequence of more conservative resource usage, and therefore they may not be competitive with other, less conservative species [28,29]. Moreover, some studies on Abies concolor, Pinus lambertiana and P. sylvestris L. have correlated slow growth and sudden decreases in growth with a higher mortality [30,31].
Therefore, growth plasticity, the capability of plants to display a high growth rate in good climatic conditions and a low growth rate in drought conditions, can be considered a very favorable trait, especially in variable environments [32].
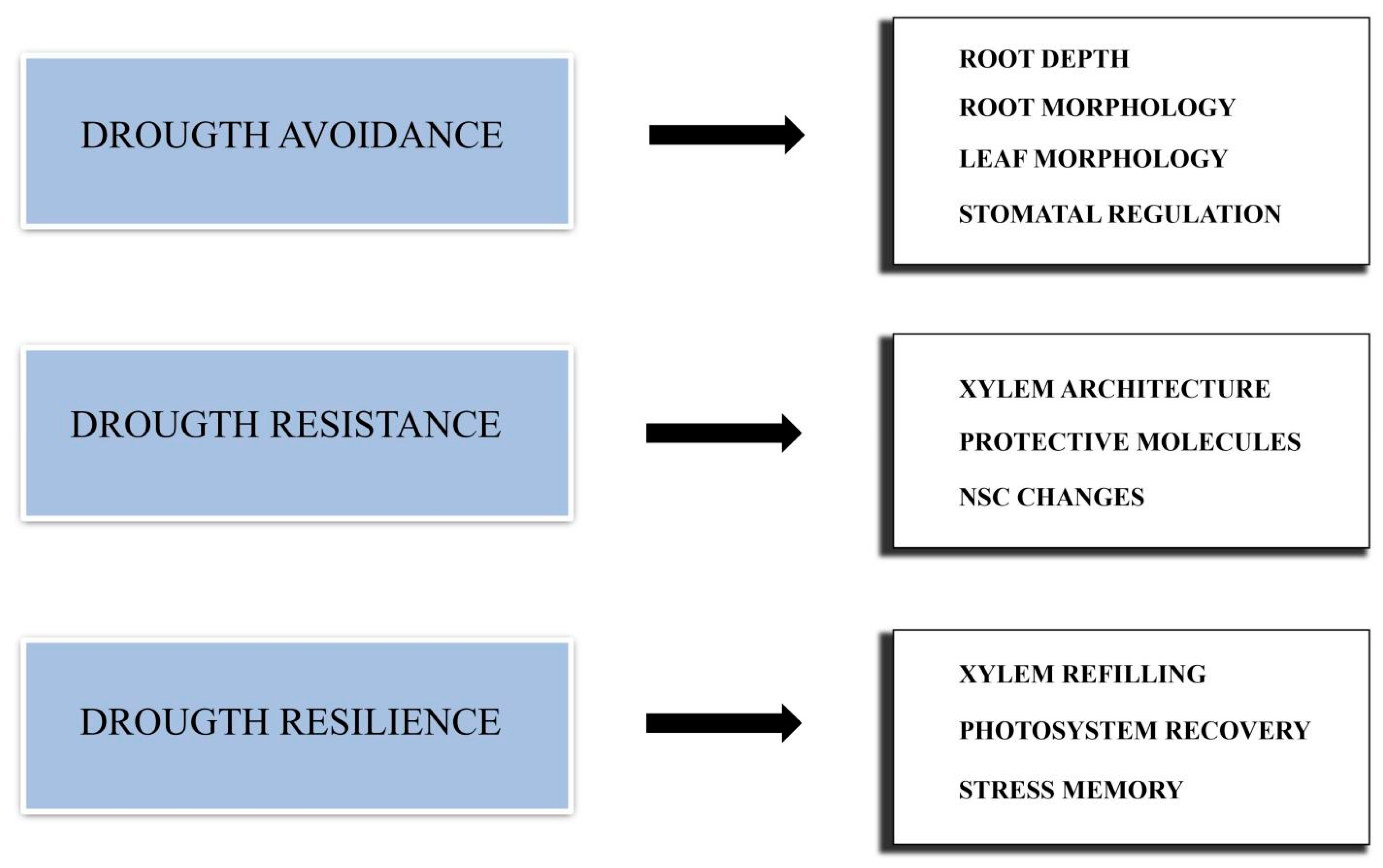
Different types of DS actually exist. A single extreme episode of drought is a different type of stress compared to the stress resulting from low but constant water availability. Moreover, a different kind of stress results from multiple drought episodes occurring during a growing season, and even a single drought event can occur in different periods of a plant’s growing season. Therefore, plant response is very complex, and multiple strategies should be adopted to cope with drought. Such strategies can be divided into three main groups: drought avoidance, drought resistance and drought resilience (Figure 1).
2.1. Drought Avoidance
Drought avoidance is the capability of a tree to minimize water loss and maximize water uptake [33,34]. Avoidance mechanisms are generally characteristic of a species and act at the whole-plant- or organ-level. Plants with a deeper root system are more efficient in reaching subsoil water [35] and less prone to experiencing water stress during the growing season than plants with shallow roots. Root depth can also influence a plant’s hydraulic status. For several conifers (Abies lasiocarpa, Picea engelmannii, Pseudotsuga menziesii), different studies have indicated that deeper water sources, mostly derived from snow melt, are primary reservoirs to support forest water demand [36,37]. Roots with a small diameter and a high ratio between their surface area and volume may present a lower susceptibility to cavitation and therefore an increased drought resistance [38]. On the other hand, drought can induce a severe dieback of the fine roots in Larix decidua, leaving the plant with a very small amount of live roots [39], which in turn can limit the plant’s response to precipitation pulses [40]. In Norway spruce, even a moderate drought was shown to modify the total biomass and the depth distribution of its fine roots [41]. In this species, an extended drought induces the deeper growth of fine roots and fungal hyphae [42].
Root and branch patterns can also influence the timing and severity of water stress, while the total numbers of branches and leaves have an effect on plant transpiration. Conifers can respond to water stress by modifying their needle morphology. In Picea abies, the needle morphology was shown to be affected by DS, but only in the portions of the canopy exposed to sunlight [43]. Another study found a reduction in the needles’ tracheid dimensions, cross-sectional area and xylem area [44]. A strategy to reduce the leaf area index and consequently transpiration is to induce branch die-back during severe droughts [45]. As the water availability decreases, conifers can induce stomatal closure by two distinct mechanisms: isohydric trees use abscisic acid (ABA) as a messenger to induce stomatal closure [46], while anisohydric species, such as many Cupressaceae and some Taxaceae, let the water potential drop and use it as a signal for stomatal closure [33].
2.2. Drought Resistance
When drought avoidance strategies are not enough to overcome a period of low water availability, plants need to cope with DS [34]. Drought resistance is the ability of a plant to withstand such stress, with a series of characteristics that can be constitutive or induced. As an example, the xylem architecture can deeply influence the drought resistance of trees. Tracheids with a larger lumen are more prone to hydraulic failures [47,48]. In fact, conifer species with a higher hydraulic conductivity and photosynthetic capacity were found to be more sensitive to DS and tended to show weaker growth resistance to extreme drought events [49,50]. In contrast, more lignified tracheid walls and smaller inter-tracheid pits can increase their DT [51,52]. The lumen width and cell wall thickness of tracheids were found to vary throughout different seasons in Norway spruce, with those produced in dry seasons being thicker and having a smaller lumen [53,54]. These variations can affect trees’ life and growth for several years, as the xylem, once produced, usually remains functional for multiple seasons [55]. Water loss can cause multiple damages at the cellular level, such as decreased cell turgor, protein denaturation and changes in membrane fluidity [56]. In order to reduce cellular damage, plants can synthesize protective molecules, including some proteins, such as chaperonins and dehydrins [57], the amino acid proline [58], various carbohydrates [59] and isoprenoids [60]. These compounds act as osmotically active molecules, contributing to the increase in cell turgor when the water potential is low. They can also stabilize cellular proteins and prevent membranes from leaking [61,62].
NSCs mainly formed by starch and soluble sugars, in addition to protecting cells from dehydration, are an important source of energy for metabolic processes [63]. Changes in NSCs have been reported for conifers during DS, according to modalities that can be different from species to species [64,65,66]. The exact meaning of such changes and their relationship with plant growth and mortality during DS are still unclear [67,68,69,70].
Another consequence of prolonged water deficits is oxidative stress, due to the accumulation of reactive oxygen species (ROS) that in turn can damage important cellular components, such as the photosystem [71]. To cope with that, conifers can produce antioxidant molecules, such as terpenes. During DS, terpene emissions generally increase, even though the response can be modified by severe water stress and the co-occurrence of other stresses [72].
2.3. Drought Resilience
Once the water availability is restored, plants can resume normal growth. The efficiency and speed of this process can be defined as drought resilience, and it is particularly important when wood production is considered [73]. There are several mechanisms that can influence drought resilience. Refilling the xylem following cavitations is one of these mechanisms. Active refilling has been observed for some conifers [74,75], even through water absorption via the needle’s cuticle [76,77]. Nevertheless, multiple cycles of damage and repair may result in conduits that are more vulnerable to embolisms than those freshly produced by the vascular cambium [78,79]. Such a condition is known as cavitation fatigue [49,80]. The fast recovery of the photosystem’s efficiency is another characteristic that can deeply influence a plant’s resilience. This is particularly true for conifers, which are usually evergreen and therefore can be exposed more often to unfavorable water conditions than plants that go dormant [71]. Finally, a sort of stress memory seems to be present in plants. For instance, plants that are exposed to repetitive water stress can maintain a smaller stomatal aperture than that of unstressed ones, even when re-watered [81,82]. Stress memory is thought to rely on post-transcriptional and epigenetic mechanisms and may represent an advantage for plants, especially in highly variable environments [83,84].
3. Studying Drought Tolerance in Conifers
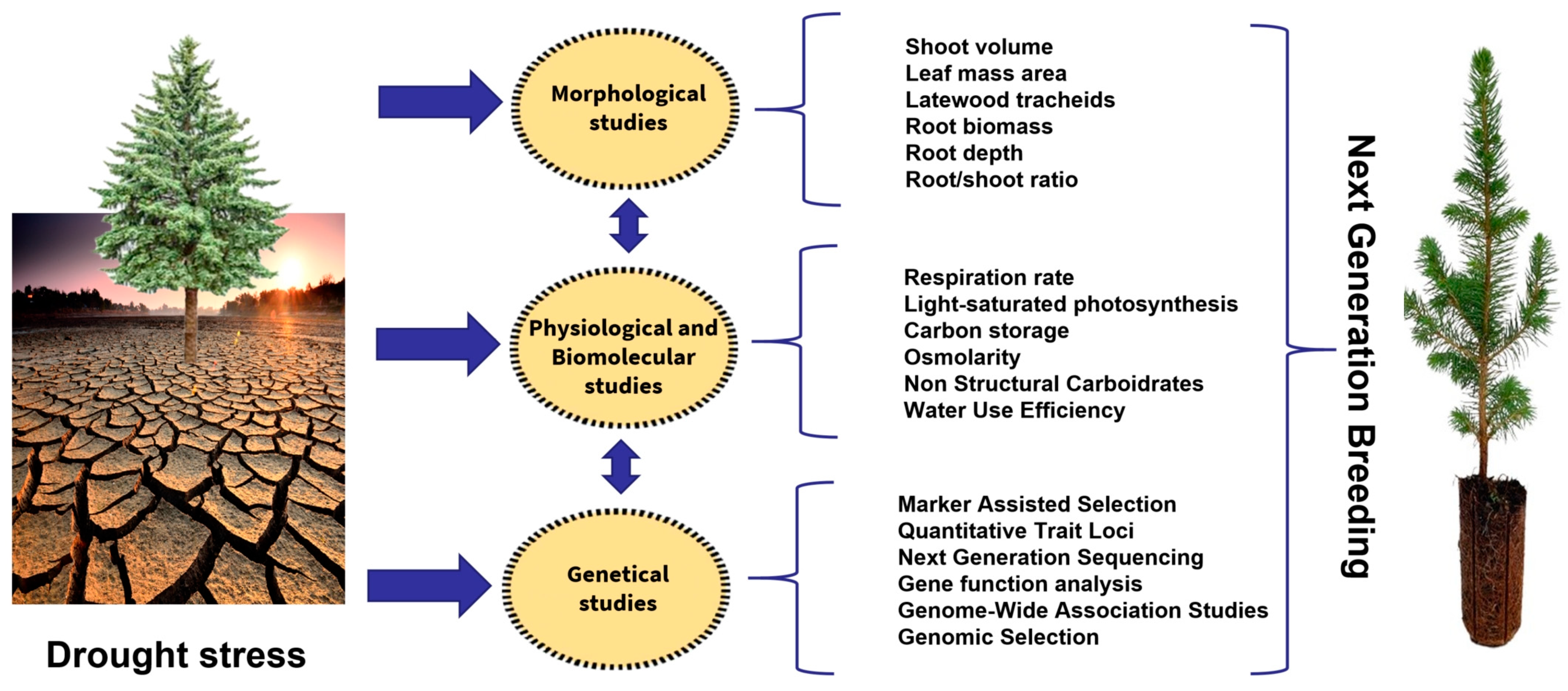
In a context of climate change, in which drought events are predicted to increase in frequency and severity in the near future [85], it is necessary to understand how plants are capable of adapting to low water availability. This is particularly true for conifers, which are often a dominant component of arid zone forests. Plant adaptations to drought can be studied at different levels. The main approaches consider traits at anatomical/morphological, physiological/biochemical and genetic levels.
3.1. Anatomical and Morphological Studies
Morphological studies usually link a specific phenotype to DT. In conifers, tree growth, often measured by the basal area increment or tree ring width, is a widely used parameter that can be correlated to meteorological data to assess plants’ responses to drought events [86]. Tree growth measurements have been applied successfully to understand the effects of forest management on the drought responses of several species, such as Pinus nigra [87], Pinus halepensis [88] and Pinus sylvestris [89,90]. The basal area increment was coupled with shoot elongation measures to compare the drought responses of adult trees and saplings in three different Pinus species, in order to predict future scenarios of relic forests under climate change [91]. In another study, the stem radial growth was used to compare the responses of Pinus edulis (isohydric species) and Juniperus monosperma (anisohydric species) to DS [92]. Tree height can be used as a morphological characteristic to study conifers’ response to drought. In a natural forest, the relationships between dominant trees and smaller ones may vary with climate change. When an ample supply of water is guaranteed, light can be the main limiting factor for plant growth, so the dominant trees are favored. In contrast, when the water availability becomes a limiting factor, the more shaded positions of smaller trees may limit transpiration and therefore compensate for the lack of light [93].
Another important morphological characteristic that is associated with drought resistance is the xylem morphology, in particular the lumen diameter and the thickness of the tracheid walls. These characteristics largely influence the xylem’s water transport efficiency as well as its safety, which influence the probability of tracheid implosions during water stress [94]. Overall, conifers are capable of modifying the xylem’s structure in response to droughts [95]. An arid climate seems to promote xylem efficiency over safety [79,96]. In a recent study on Picea abies, a macroscopic characteristic, namely stem cracks, was associated with water stress [97]. In the same study, the thickness of the tracheid walls, rather than the lumen diameter, was found to be the main anatomical characteristic associated with tracheid collapses. This finding is particularly important for all tree species exploited for wood production, as the cell wall thickness was shown to have quite a high heritability [98]. Cell wall thickness is a parameter determining wood density, and wood density has been negatively correlated to growth in Picea abies [99]. The xylem phenology was also studied in conifers in relationship to drought [100]. Characteristics, such as cell differentiation, cell enlargement and cell wall thickening, seem to be influenced by water availability in several species, such as Abies alba, Pinus sylvestris [101] and Larix decidua [102].
When studying DT in plants, one should take into account that natural populations are not homogeneous but present a certain degree of genetic variation. Morphological traits, such as the basal area increment or tree ring width, can be used to assess the inter- and intraspecific genetic variation. This is usually performed in so-called provenance or common garden studies, in which seedlings from many different regions can be planted and studied in a common environment [103,104]. Provenance studies can be conducted across multiple sites or using multiple treatments in order to estimate the plasticity of traits [105]. The intraspecific growth response to drought was studied in Abies alba using provenances from Bulgaria, Italy, Romania and Czech Republic. The ring width, earlywood width and latewood width were measured and correlated to drought events over a period of over 20 years, in order to find provenances combining a high productivity and drought resistance [106]. In a second study, 43 populations of Picea glauca were evaluated for DT by measuring a series of tree-ring traits. A significant genetic variation was found among populations in response to DS. In particular, the authors found that populations from drier geographical origins showed a higher resilience to extreme drought events when compared to populations from more humid geographical origins, indicating local genetic adaptations [107]. Similar experiments were performed using several phenotypical traits on different conifer species, such as Pinus pinea [108], Pinus ponderosa [109], Pinus sylvestris [110] and Picea abies [111]. An interspecific study was performed comparing the drought responses of Picea abies, Abies alba, Larix decidua and Pseudotsuga menziesii. This latter species had the highest drought resistance, while Abies alba had the best drought recovery. Nonetheless, even the most drought-sensitive species, Picea abies and Larix decidua, showed significant genetic variation within and among populations along their natural geographic areas, enough to justify targeted tree breeding and supportive forest management [112].
3.2. Physiological and Biochemical Studies
Most of the studies using morphological data to assess DT are focused on tree growth. This parameter is a good indicator of plant resilience, and it is very useful when wood production is the final goal. Nevertheless, DT is a very complex trait involving a number of different mechanisms. In order to identify such mechanisms and use them to highlight differences in plant responses to drought, physiological data (Table 1) can be used [113,114]. One of the most studied physiological processes related to water stress is photosynthesis [115,116,117]. In particular, the concentration of chlorophyll a can be obtained via the extrapolation of the emission of refracted light from foliage [118], therefore it is a parameter that can be measured by remote-sensing tools and can be used to monitor large areas [119,120]. Drought can damage Photosystem II (PSII), resulting in changes in fluorescence parameters [121]. The fluorescence measurement was shown to be a very sensitive proxy for DS [122], allowing researchers to assess physiological disturbances even before the appearance of visible symptoms [121]. In the last few years, several studies on conifers have been published [123,124]. In Picea abies, the chlorophyll a concentration and fluorescence parameters were measured together with the tree height to assess seedling performances under water stress. Both physiological parameters were good indicators of plants’ drought sensitivity, even though differences were found depending on the soil type [125]. In Abies alba, the chlorophyll a fluorescence was measured, testing five provenances from different altitudes under mild water stress. Significant differences were found, with provenances from higher altitudes showing better performances under both optimal and low-water conditions, suggesting that there were local adaptations to drought and that fluorescence parameters can be applied during plant selection for resistant seedlings [126].
Another important physiological parameter that has been often used to assess plant DT is water use efficiency (WUE), which is the ratio between the carbon fixed by photosynthesis and the water loss [127,128,129]. Nevertheless, caution must be taken when using WUE as an indicator for DT, because it depends on different mechanisms, such as photosynthesis and transpiration, which can both vary and not always in the same way.
Moreover, several measures of WUE exist and even if they are often correlated, they are not interchangeable [130], so they may lead to contrasting results. One of the most frequently used methods for measuring WUE is the carbon ratio δ13 C [131]. In conifers, some studies have found higher δ13 C in populations originating from drier sites [28], while others have shown the opposite [132]. In Pinus halepensis, individuals from dry sites showed a lower WUE plasticity than those from mesic sites [128], while in another study, a higher average WUE was shown in individuals from drier sources [127]. Finally, it must be noted that, especially if measured during the whole growing season, high water usage due to highly plastic growth can reduce the WUE, even though plastic growth can be considered a desirable characteristic in moderately dry climates [28]. For all these reasons, it is always advisable to integrate WUE with other parameters when dealing with drought resistance [49,133]. As an example, in Pinus mariana, changes in morphological (shoot volume, leaf mass area), physiological (respiration rate, light-saturated photosynthesis) and biochemical (osmolality, NSC) parameters were considered to quantify the impact of temporary drought and re-hydration on its internal carbon dynamics and shoot development [49,134]. More in general, biochemical data can be very useful to integrating physiological studies and understanding conifer responses to DS. Osmolytes, such as proline, chlorophyll a and b, and antioxidants, such as anthocyanins, carotenoids and phenolic compounds, can be measured and correlated to water availability [62]. In Larix decidua, six Carpathian populations were submitted to a month of mild DS under controlled conditions. Several biochemical parameters were measured, looking for the optimal biochemical markers for an early detection of drought symptoms [135]. A lot of attention has been given in the last decade to NSC dynamics, specifically to understanding whether a dehydration-induced carbohydrate limitation (source limitation) or a direct effect of low water availability (sink limitation) is responsible for reducing plant growth under DS [66,75,136]. Some indications seem to suggest that growth is not carbon-limited by drought as growth reductions may occur along with carbohydrate storage increases [68,137]. On the other hand, carbon reserves could be formed at the expense of growth as a strategy to prevent carbon starvation, a very dangerous condition for plants [138]. Moreover, in cases of longer and more severe droughts, a reduction in both the carbon storage and growth was observed [139].

{kind=link}
{kind=link}
Table 1.
Physiological traits relevant for drought stress.
| Plant Traits | Species | Relevant Effects | Modulation under Stress | References |
|---|---|---|---|---|
| Photosynthetic activity | Pinus spp., Picea spp., Pseudotsuga menziesii, Abies alba, | Modulation of Calvin cycle and photosystem damage. | Reduction under stress. | [50,114,118,119,122,123,125,126] |
| Carbon allocation and water-use efficiency | Pinus spp., Picea mariana | Compensation of reduced photosynthesis by increased remobilization. | Growth reduction for carbon investments in the maintenance of respiration and osmoprotection. | [18,28,49,68,127,132,134,137] |
| Stomatal conductance | Picea abies, Larix decidua, Pinus radiata | Water consumption and storage. Leaf temperature regulation. | Increased stomatal resistance may help to prevent the xylem embolism. | [46,82,117] |
| Xylem cavitation and embolism | Pinus spp., Picea spp., Abies spp., Juniperus spp., Pseudotsuga menziesii, Larix decidua, Taxus baccata | Cavitation. Hydraulic conductivity. Growth capacity. Drought-induced mortality. | Isolation of embolized tracheids. Changes in carbohydrate contents. Osmotic adjustments. | [14,48,78,79] |
| Phenological phases | Pinus sylvestris, Abies alba, Larix decidua | Growth effects. Production and morphology of latewood tracheids. | Reduced growth. Phenological shifts to compensate for DS. | [100,101,102,133] |
| Root morphology and depth | Picea abies, Larix sibirica | Higher/lower tapping of soil water resources. | Reduced root biomass, increased root/shoot ratio, fine-root production decreased. Root depth increased. | [39,41,42] |
| Protection against active oxygen species | Pinus spp., Picea spp., Pseudotsuga menziesii, Larix laricina, Abies balsamea | Oxidative stress | Production of protective molecules. | [57,58,59,60,72] |
3.3. Molecular Genetics Studies
To further investigate drought responses and link morphological and physiological traits to the genes and/or genomic regions that are responsible for these characteristics, molecular genetics can be used [140]. One of the most applied approaches is a gene expression analysis. Although there are a lot of techniques to study gene expression, in the most recent publications, whole-transcriptome approaches [141,142,143,144,145,146] and/or quantitative real-time PCR (qRT-PCR) [143,144,147] have been used more frequently. In contrast to RNAseq and other whole-transcriptome techniques, qRT-PCR is very sensitive but in most cases it is used to study one or a few specific target genes [148] or to confirm the gene expression results obtained with less sensitive methods [144,149]. Via expression analyses, a range of genes have been identified that might be involved in the drought responses in conifers (Table 2).
Genes related to signaling and transcription factors, including AP2/ERF, bZIP, TCP, WRKY and MYB, have been found to be regulated during water stress in several conifer species, such as Larix kaempferi [150], Pinus massoniana [151], Abies alba [149] and Pinus taeda [152]. All these genes, due to their regulatory function, are usually expressed quite early during DS and therefore they could be considered good indicators of an efficient plant response. Nevertheless, this is not always the case. In Pinus pinaster, it was found that tolerant individuals can be pre-adapted to cope with drought, constitutively expressing stress-related genes; in contrast, in more sensitive individuals, these are induced by the onset of stress [142]. So, care must be taken when considering gene expression, and distinguishing between the different drivers of observed differences is of primary importance. Another important factor that must be considered when studying gene expression during drought is the type of treatment used. In some studies, DS was induced by stopping irrigation [145]; in others, a chemically-induced stress was used [153]. In some cases, the water was withheld for a given period [153]; in others, it was withheld until the needles wilted [152]. Thus, there is always the possibility of methodological artifacts [154]. Abscisic acid (ABA), a plant hormone involved in stomatal closure, shoot growth and water uptake, was shown to regulate many structural genes in conifers [144,145,151] even though ABA-independent pathways also exist in many species [155]. Other genes regulated during drought include those encoding for antioxidants [141,150], protective molecules, such as late embyogenesis abundant (LEA) proteins, which are thought to stabilize proteins and membranes [156], genes involved in lignin and sugar biosynthesis [150], flavonoid and terpenoid biosynthesis [141], aquaporins, which can affect the water permeability of membranes [151] and even pathogen resistance genes, such as nucleotide-binding, leucine-rich repeat proteins [157,158].
All the genes identified by expression analyses may have a role in drought responses, but it is not always easy to link a gene’s expression, which is specific for the given time point in question, to more complex morphological and physiological traits, which are the result of different processes acting over a longer period of time. To do so, molecular markers can be used for quantitative trait locus (QTL), genome scan and genotype association studies. All these approaches aim at linking genes or genomic regions to a trait. Classic QTL studies exploit segregating populations to identify genomic regions linked to a polygenic trait but have had very limited success in conifers [130,159,160]. The main reason is the huge amount of space and time necessary to cross parental individuals and obtain enough progeny. Moreover, conifers have very large genomes, usually with a low linkage disequilibrium (LD), so high-resolution maps are needed for QTL detections and candidate gene identification [161]. The development of high-throughput systems, such as next-generation sequencing and single nucleotide polymorphism (SNP) arrays, is gradually overcoming such problems, allowing for association-mapping studies that exploit evolutionary recombinations in natural populations [162]. Nonetheless, until very recently, studying the molecular basis of heritable trait variation was still considered quite challenging in conifers and most of the studies were focused on a limited number of candidate genes [163,164,165,166,167,168]. This approach, although effective, is very limiting for the identification of new loci. To overcome this problem, genome-wide association studies (GWASs) can be used. As an example, a GWAS was performed on Sequoiadenron giganteum and Sequoia sempervirens, trying to link 10 drought-related morphological and physiological traits to SNP markers. Seventy-eight new marker x trait associations were found in Sequoia sempervirens and six in Sequoiadenron giganteum, identifying a wide range of candidate genes involved in different biological processes, such as signaling, stress response and growth [169]. The genome size of conifers, which can range between 8 and 34 Gb [170], may still represent a problem. With some techniques, tens of thousands of SNPs can be obtained, covering most of the genome [171,172,173]. Nonetheless, most of such SNPs will be in noncoding regions. This, on one hand, is good for the identification of regulatory regions, but on the other hand, it may limit the number of gene associations detected. With the latest advances in sequencing and bioinformatics, it is possible to limit SNP analyses to the protein-coding region of genomes, the so-called exome [174]. With such an approach, the complexity of the conifer genome can be massively reduced, and even though the exome represents only a small fraction of the total genome, gene association is greatly facilitated [175]. Examples of exome-derived SNP association studies exist for several conifer species, such as Pseudotsuga menziesii [176] and Pinus taeda [177].
Table 2.
Main categories of genes related to drought responses in conifers.
| Main Category | Subcategory | Species | Reference |
|---|---|---|---|
| Protective molecules | Dehydrins | 5, 9, 10, 11 | [141,148,149,152,156] |
| LEA | 5, 9, 10, 11 | [141,149,152,156] | |
| HSP | 5, 11 | [152,156] | |
| Antioxidant | 1, 5, 9, 11 | [141,145,146,151,152,156] | |
| Other | 1, 5, 10, 11 | [146,149,151,152] | |
| Gene transcription and signalling | TF | 1, 2, 3, 4, 5, 9, 10, 11 | [141,144,145,146,149,150,151,152,156,164] |
| Hormones | 1, 2, 3, 5, 7, 9, 10, 11 | [141,144,145,146,147,149,150,151,152,156] | |
| Protein kinases | 2, 3, 5, 9, 10 | [141,144,149,150,156] | |
| Other | 2, 5 | [144,146,156] | |
| Metabolic activity | Sugar | 1, 3, 4, 5, 9, 10, 11 | [141,145,149,150,152,156,164] |
| Lipid | 1, 5, 9, 10 | [141,145,149,156] | |
| Cell wall | 5, 9, 10, 11 | [141,146,149,152] | |
| Flavonoids and phenylpropanoids | 1, 3, 4, 7, 9 | [141,145,147,150,164] | |
| Other | 1, 9 | [141,145] | |
| Protein degradation | 5, 9 | [141,156] | |
| Photosynthesis | 1, 5, 6, 7, 9 | [71,141,145,146] | |
| Transport | 1, 5, 9, 10, 11 | [141,146,149,151,152,156] | |
| Biotic stress | 1, 4, 5, 8, 10, 11 | [149,151,152,156,158] |
Species: 1: Pinus massoniana; 2: Pinus tabuliformis; 3: Larix kaempferi; 4: Picea glauca; 5: Pinus pinaster; 6: Picea abies; 7: Pinus sylvestris; 8: Pinus koraiensis; 9: Pinus halepensis; 10: Abies alba; 11: Pinus taeda.
4. Next Generation Breeding
The increasing amount of morphological, physiological and molecular data on conifer drought resistance can have practical applications in improving breeding programs [178,179,180,181] (Figure 2).
As a matter of fact, traditional breeding programs for long-living trees, such as conifers, face a number of challenges. The most important is the great amount of space and time necessary to obtain significant achievements. Even though several breeding programs started in the 1950s, many of them are only in their first or second cycle of selection, with the most advanced ones, such as the loblolly pine (Pinus taeda L.) breeding program in the USA, in the third or fourth cycles [182]. Moreover, due to the high costs, tree breeding programs have been developed mainly for commercially important species and only recently has it become evident that it is important to also protect natural forests, which are often threatened by climate change [183]. In cases of drought, plant resistance or tolerance is controlled by multiple genes, each with a minor additive effect, so it is particularly challenging to achieve and maintain the desired level of resistance [184].
Another important factor that must be taken into account, especially when considering natural forests, is genetic diversity. Resistant populations developed by breeding programs have to maintain all the adaptive traits typical of the species or provenance, such as growth rate, cold resistance and pathogen resistance. Therefore, it is necessary to exclude the presence of negative correlations between drought resistance and other adaptive traits [182].
New emerging technologies can be efficiently used to link plant genome to phenotypic data and therefore could be used to improve the efficiency of traditional breeding and accelerate the selection of valuable genotypes. Nowadays, high-throughput technologies can be applied not only to genotyping but also to phenotyping, improving both data acquisition and analytical pipelines, as well as possibly leading to an unprecedented revolution in the way we have studied agriculture till now [185]. As an example, a number of portable and user-friendly chlorophyll fluorimeters have been introduced in the last few years. These, together with more sophisticated high-throughput hyperspectral imaging systems, have enhanced the accuracy of chlorophyll measurements. In this way, large phenotyping platforms have been implemented with an automatic control and data analysis system, allowing researchers to keep a large number of plants in parallel under constant monitoring, over long periods of time and in different environmental conditions [186]. These high-throughput phenotyping technologies are generally employed in controlled environments, such as growth chambers and greenhouses, so there are some limitations when predicting the performance of a plant under field conditions. Nonetheless, these systems can be very useful in studying how phenotypes change among different genotypes under uniform stress conditions [187].
A large amount of genomic resources is available for conifers. Despite their size and complexity, several conifer genomes have been sequenced [188,189,190,191,192,193]. A complete list of genomic resources of conifers, with the year of release, the Genebank storage and the accession number, has been reported by Traversari et al. [194]. These data, together with transcriptome studies, have provided useful information for the development of high-density genetic maps [195,196,197,198,199] and SNP arrays [200,201,202,203,204], which could be used in next-generation breeding approaches. To date, marker-assisted selection (MAS) has shown only a limited application in forest breeding [205,206]. The main reason is that MAS is quite reliable and easy to use in cases of monogenic characteristics, but it is much harder to be applied in cases of more complex traits, such as drought, as several markers linked to genetic loci involved in the control of the resistance must be identified. As already stated (see previous paragraph), classic QTL mapping had little success in conifers for the identification of genetic loci linked to traits of interest. Moreover, each locus can control a relatively small percentage of the resistance, so several loci should be screened at the same time in order to reach the desired level of resistance and should be validated using different populations before being routinely used for assisted selection [104,207]. Overall, it was shown that QTLs are not effectively suitable for MAS in forest trees, as they do not explain enough genetic variations for complex traits, such as drought resistance [208]. To date, one of the most promising approaches for next-generation breeding is genomic selection (GS) [209,210,211]. GS typically uses a large genome-wide panel of molecular markers (usually SNPs) whose effects on the desired trait are estimated in a so-called training population, which in forest trees is generally composed of one to a few thousand individuals. The molecular markers are used to build prediction models that can be later applied to any candidate resistant tree for which only the genotype is known, while the phenotype can be predicted by the models [206]. This is an obvious advantage, as with GS, previous information on the phenotype-marker linkage or localization of QTLs on the genome and their relative effect on the phenotypic variance becomes unnecessary [212]. The breeding cycle could be significantly shortened, as with GS, marker-based selection is performed on seedlings at a very early stage, eliminating the need for expensive and time-consuming field testing [211]. As for traditional phenotypic selection, to increase GS accuracy, it is advisable to test the population in an environment that is similar to the target environment in which plants are expected to live, in order to avoid genotype x environment interactions [210,213]. GS is currently used routinely for livestock breeding but its application to forest breeding is still very limited. Nonetheless, several works exploring the potential of GS in conifer breeding already exist, mostly focused on traits, such as growth and wood quality. In Pinus taeda, genomic breeding values were estimated on 149 individuals of a cloned progeny from 13 crosses, using 3406 markers simultaneously. The accuracy varied according to the considered trait, ranging between 0.61 and 0.83 for the lignin and cellulose content, while it was lower (0.30 to 0.68) for growth [214]. In Pinus pinaster, 2500 SNPs were used for genotyping 661 individuals from two different generations. The average prediction accuracy using different statistical models and validation scenarios was between 0.4 and 0.5, with small differences according to the considered trait [212]. Other studies evaluated the accuracy of GS in several conifer species, such as Picea mariana [215], Picea abies [216,217] and Picea glauca [218]. Overall, the results obtained with GS in conifers are encouraging, even though significant differences can be found depending on the number of markers [217,219] and types of population used [208,220]. GS studies on DT in trees are still very rare. To our knowledge, the first one was published on a Eucalyptus hybrid population, evaluating 1130 clones with 3303 SNPs [221], but there is no doubt that in the near future research on this area will expand.
5. Conclusions and Future Directions
Plant adaptations to DS are likely to become progressively more important in the near future, as most of the recent climate models suggest an increase in the length and intensity of dry periods in many regions [222]. Conifer response to low water availability is a very complex trait, involving a variety of different physiological processes and molecular mechanisms. Nonetheless, in the last decade, substantial progress has been made in understanding drought responses, especially with the availability of new technologies that allow high-throughput phenotyping and genotyping. Many genes and pathways involved in the control of drought responses have been discovered and analyzed in detail, such as those related to ABA, signaling, carbohydrate metabolism and the production of protecting molecules [140]. The availability of a great number of low-cost molecular markers, mostly SNPs, is making it possible to apply new advanced approaches to tree breeding [84], so that techniques, such as GS, could be applied routinely to substantially reduce breeding cycles [211]. However, not all the traits that are thought to be involved in drought resistance have been thoroughly understood, because conifers include a great number of species, both isohydric and anisohydric, so the strategies to adapt and survive in the face of water stress might be different between families, genera and species. Till now, most of the efforts in studying drought responses were performed on species with a high economical value, especially those grown on plantations. This is understandable, but it means that many ecologically important taxa, such as Cupressaceae, should receive more attention, as they could provide valuable information, being remarkably drought-tolerant [46]. Efforts have been made to integrate different approaches to better understand conifer adaptations to water stress [49,66,133,135,145,164]; most of the studies, especially those looking at gene expression, were performed on seedlings or young ramets [142,144,145,150,223]. This, although important to understanding the molecular mechanisms involved in plant responses to drought, does not necessarily reflect the behavior of an adult tree when subjected to the same conditions. It is an example the remarkably different requirements present in several species between seedlings and mature trees in relation to their shade tolerance [224]. Therefore, it would be advisable to also focus on mature plants. Another factor that should be taken into account is the type of DS considered. A single extreme episode of drought is different from a low but constant water availability or multiple stress episodes during the growing season. Plant strategies to cope with different types of stress could be different, so it is important to try to match, as much as possible, the range of conditions that the studied plant may face in the natural environment, as well as in cultivation. Only integrating data from multiple disciplines and considering all the variables that could influence plant drought responses in a natural context, including the type of soil [225,226] and microbiome [227,228,229], will a more thorough comprehension of this complex trait in conifers be possible.
Author Contributions
Conceptualization, P.B. and N.L.P.; methodology, P.B. and N.L.P.; validation, P.B. and N.L.P.; formal analysis, P.B. and N.L.P.; investigation, P.B. and N.L.P.; resources, P.B. and N.L.P.; data curation, P.B. and N.L.P.; writing—original draft preparation, P.B. and N.L.P.; writing—review and editing, P.B. and N.L.P.; visualization, P.B. and N.L.P.; supervision, P.B. and N.L.P.; project administration, P.B. and N.L.P.; funding acquisition, P.B. and N.L.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by FEM projects with the grant numbers P1611006I and P1611034.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bussotti, F.; Pollastrini, M. Revisiting the concept of stress in forest trees at the time of global change and issues for stress moni-toring. Plant Stress 2021, 2, 100013. [Google Scholar] [CrossRef]
- Caballero, C.B.; Ruhoff, A.; Biggs, T. Land use and land cover changes and their impacts on surface-atmosphere interactions in Brazil: A systematic review. Sci. Total. Environ. 2022, 808, 152134. [Google Scholar] [CrossRef] [PubMed]
- Lloret, F.; Jaime, L.A.; Margalef-Marrase, J.; Perez-Navarro, M.A.; Batllori, E. Short-term forest resilience after drought-induced die-off in Southwestern European forests. Sci. Total Environ. 2022, 806, 150940. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Gonzalez-Moreno, P.; Ruiz-Gomez, F.J.; Sanchez-Cuesta, R.; Gazol, A.; Camarero, J.J. Drought stress and pests increase defoliation and mortality rates in vulnerable Abies pinsapo forests. For. Ecol. Manag. 2022, 504, 119824. [Google Scholar] [CrossRef]
- McDowell, N.G.; Williams, A.P.; Xu, C.; Pockman, W.T.; Dickman, L.T.; Sevanto, S.; Pangle, R.; Limousin, J.; Plaut, J.; Mackay, D.S.; et al. Multi-scale predictions of massive conifer mortality due to chronic temperature rise. Nat. Clim. Change 2016, 6, 295–300. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Leites, L.P.; St Clair, J.B.; Jaquish, B.C.; Saenz-Romero, C.; Lopez-Upton, J.; Joyce, D.G. Comparative genetic re-sponses to climate in the varieties of Pinus ponderosa and Pseudotsuga menziesii: Clines in growth potential. For. Ecol. Manag. 2014, 324, 138–146. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Tchebakova, N.M.; Parfenova, Y.I.; Wykoff, W.R.; Kuzmina, N.A.; Milyutin, L.I. Intraspecific responses to climate in Pinus sylvestris. Glob. Change Biol. 2002, 8, 912–929. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, K.; Germino, M.; Kueppers, L.M.; Domec, J.-C.; Mitton, J. Linking carbon and water relations to drought-induced mortality in Pinus flexilis seedlings. Tree Physiol. 2015, 35, 771–782. [Google Scholar] [CrossRef] [Green Version]
- Sevanto, S.; McDowell, N.G.; Dickman, L.T.; Pangle, R.; Pockman, W.T. How do trees die? A test of the hydraulic failure and car-bon starvation hypotheses. Plant Cell Environ. 2014, 37, 153–161. [Google Scholar] [CrossRef]
- Diaz-Sala, C.; Cabezas, J.A.; Fernández de Simón, B.; Abarca, D.; Guevara, M.Á.; de Miguel, M.; Cadahía, E.; Aranda, I.; Cervera, M.T. The uniqueness of conifers. In From Plant Genomics to Plant Biotechnology, 1st ed.; Poltronieri, P., Burbulis, N., Fogher, C., Eds.; Woodhead Publishing: Sawston, UK, 2013; pp. 67–96. [Google Scholar] [CrossRef]
- Mitchell, P.J.; O’Grady, A.P.; Tissue, D.T.; Worledge, D.; Pinkard, E.A. Co-ordination of growth, gas exchange and hydraulics define the carbon safety margin in tree species with contrasting drought strategies. Tree Physiol. 2014, 34, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Wolf, A. Pervasive drought legacies in forest eco-systems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperry, J.S.; Hacke, U.G.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosner, S.; Johnson, D.M.; Voggeneder, K.; Domec, J.C. The conifer-curve: Fast prediction of hydraulic conductivity loss and vulnerability to cavitation. Ann. For. Sci. 2019, 76, 82. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.M.; McCulloh, K.A.; Woodruff, D.R.; Meinzer, F.C. Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different? Plant Sci. 2012, 195, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Carnicer, J.; Barbeta, A.; Sperlich, D.; Coll, M.; Peñuelas, J. Contrasting trait syndromes in angiosperms and conifers are associat-ed with different responses of tree growth to temperature on a large scale. Front. Plant Sci. 2013, 4, 409. [Google Scholar] [CrossRef] [Green Version]
- Meinzer, F.C.; Woodruff, D.R.; Eissenstat, D.M.; Lin, H.S.; Adams, T.S.; McCulloh, K.A. Above- and belowground controls on water use by trees of different wood types in an eastern US deciduous forest. Tree Physiol. 2013, 33, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Rehschuh, R.; Ruehr, N.K. Diverging responses of water and carbon relations during and after heat and hot drought stress in Pinus sylvestris. Tree Physiol. 2021, 42, 1532–1548. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; García-Valdés, R.; Ruíz-Benito, P.; Zavala, M.A. Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change. Glob. Change Biol. 2011, 17, 2400–2414. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xia, S.; Zeng, D.; Liu, C.; Li, Y.; Yang, W.; Yang, B.; Zang, J.; Slik, F.; Lindenmayer, D.B. Age and spatial distribution of the world’s oldest trees. Conserv. Biol. 2022, 36, e13907. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Vicente-Serrano, S.M.; Sánchez-Salguero, R.; Gutiérrez, E.; de Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; Tíscar, P.A.; et al. Forest resilience to drought varies across biomes. Glob. Change Biol. 2018, 24, 2143–2158. [Google Scholar] [CrossRef]
- Pereira, L.; Domingues, A.P.; Jansen, S.; Choat, B.; Mazzafera, P. Is embolism resistance in plant xylem associated with quantity and characteristics of lignin? Trees-Struct. Funct. 2018, 32, 349–358. [Google Scholar] [CrossRef]
- Hasan, M.M.; Gong, L.; Nie, Z.F.; Li, F.P.; Ahammed, G.J.; Fang, X.W. ABA-induced stomatal movements in vascular plants during dehydration and rehydration. Environ. Exp. Bot. 2021, 186, 104436. [Google Scholar] [CrossRef]
- Pandey, S. Climatic influence on tree wood anatomy: A review. J. Wood Sci. 2021, 67, 24. [Google Scholar] [CrossRef]
- Yang, G.S.; Huang, L.; Shi, Y.F. Magnitude and determinants of plant root hydraulic redistribution: A global synthesis analysis. Front. Plant Sci. 2022, 13, 918585. [Google Scholar] [CrossRef]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Change Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef]
- Eilmann, B.; Rigling, A. Tree-growth analyses to estimate tree species’ drought tolerance. Tree Physiol. 2012, 32, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Kerr, K.L.; Meinzer, F.C.; McCulloh, K.A.; Woodruff, D.R.; Marias, D.E. Expression of functional traits during seedling establish-ment in two populations of Pinus ponderosa from contrasting climates. Tree Physiol. 2015, 35, 535–548. [Google Scholar] [CrossRef] [Green Version]
- Montwé, D.; Spiecker, H.; Hamann, A. Five decades of growth in a genetic field trial of Douglas-fir reveal trade-offs between productivity and drought tolerance. Tree Genet. Genomes 2015, 11, 29. [Google Scholar] [CrossRef]
- Das, A.J.; Battles, J.J.; Stephenson, N.L.; van Mantgem, P.J. The relationship between tree growth patterns and likelihood of mortality: A study of two tree species in the Sierra Nevada. Can. J. For. Res. 2007, 37, 580–597. [Google Scholar] [CrossRef]
- Hereş, A.M.; Martínez-Vilalta, J.; Claramunt López, B. Growth patterns in relation to drought-induced mortality at two Scots pine (Pinus sylvestris L.) sites in NE Iberian Peninsula. Trees 2012, 26, 621–630. [Google Scholar] [CrossRef]
- Santos-Del-Blanco, L.; Bonser, S.P.; Valladares, F.; Chambel, M.R.; Climent, J. Plasticity in reproduction and growth among 52 range-wide populations of a Mediterranean conifer: Adaptive responses to environmental stress. J. Evol. Biol. 2013, 26, 1912–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Valladares, F.; Aranda, I. Mediterranean trees coping with severe drought: Avoidance might not be safe. Environ. Exp. Bot. 2018, 155, 529–540. [Google Scholar] [CrossRef]
- Pirtel, N.L.; Hubbard, R.M.; Bradford, J.B.; Kolb, T.E.; Litvak, M.E.; Abella, S.R.; Porter, S.L.; Petrie, M.D. The aboveground and belowground growth characteristics of juvenile conifers in the southwestern United States. Ecosphere 2021, 12, e03839. [Google Scholar] [CrossRef]
- Martin, J.; Looker, N.; Hoylman, Z.; Jencso, K.; Hu, J. Differential use of winter precipitation by upper and lower elevation Douglas fir in the Northern Rockies. Glob. Change Biol. 2018, 24, 5607–5621. [Google Scholar] [CrossRef]
- Berkelhammer, M.; Still, C.J.; Ritter, F.; Winnick, M.; Anderson, L.; Carroll, R.; Carbone, M.; Williams, K.H. Persistence and plas-ticity in conifer water-use strategies. J. Geophys. Res. Biogeosci. 2020, 125, e2018JG004845. [Google Scholar] [CrossRef]
- Phillips, R.P.; Ibáñez, I.; D’Orangeville, L.; Hanson, P.J.; Ryan, M.G.; McDowell, N.G. A belowground perspective on the drought sensitivity of forests: Towards improved understanding and simulation. For. Ecol. Manag. 2016, 380, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Chenlemuge, T.; Hertel, D.; Dulamsuren, C.; Khishigjargal, M.; Leuschner, C.; Hauck, M. Extremely low fine root biomass in Larix sibirica forests at the southern drought limit of the boreal forest. Flora 2013, 208, 488–496. [Google Scholar] [CrossRef]
- Plaut, J.A.; Wadsworth, W.D.; Pangle, R.; Yepez, E.A.; McDowell, N.G.; Pockman, W.T. Reduced transpiration response to precip-itation pulses precedes mortality in a piñon-juniper woodland subject to prolonged drought. New Phytol. 2013, 200, 375–387. [Google Scholar] [CrossRef]
- Konôpka, B.; Lukac, M. Moderate drought alters biomass and depth distribution of fine roots in Norway spruce. For. Pathol. 2013, 43, 115–123. [Google Scholar] [CrossRef]
- Børja, I.; Godbold, D.; Světlík, J.; Nagy, N.E.; Gebauer, R.; Urban, J.; Volařik, D.; Lange, H.; Krokene, P.; Čermák, P.; et al. Norway spruce fine roots and fungal hyphae grow deeper in forest soils after extended drought. In Soil Biological Communities and Ecosystem Resilience. Book Series: Sustainability in Plant and Crop Protection; Lukac, M., Grenni, P., Gamboni, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 123–142. [Google Scholar] [CrossRef]
- Gebauer, R.; Volarik, D.; Urban, J.; Borja, I.; Nagy, N.E.; Eldhuset, T.D.; Krokene, P. Effects of mild drought on the morphology of sun and shade needles in 20-year-old Norway spruce trees. Iforest 2019, 12, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Gebauer, R.; Volařík, D.; Urban, J.; Børja, I.; Nagy, N.E.; Eldhuset, T.D.; Krokene, P. Effects of prolonged drought on the anatomy of sun and shade needles in young Norway spruce trees. Ecol. Evol. 2015, 5, 4989–4998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koepke, D.F.; Kolb, T.E.; Adams, H.D. Variation in woody plant mortality and dieback from severe drought among soils, plant groups, and species within a northern Arizona ecotone. Oecologia 2010, 163, 1079–1090. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; McAdam, S.A.M.; Jordan, G.J.; Martins, S.C.V. Conifer species adapt to low-rainfall climates by following one of two divergent pathways. Proc. Natl. Acad. Sci. USA 2014, 111, 14489–14493. [Google Scholar] [CrossRef] [Green Version]
- Dulamsuren, C.; Abilova, S.B.; Bektayeva, M.; Eldarov, M.; Schuldt, B.; Leuschner, C.; Hauck, M. Hydraulic architecture and vul-nerability to drought-induced embolism in southern boreal tree species of Inner Asia. Tree Physiol. 2019, 9, 463–473. [Google Scholar] [CrossRef]
- Feng, F.; Losso, A.; Tyree, M.; Zhang, S.; Mayr, S. Cavitation fatigue in conifers: A study on eight European species. Plant Physiol. 2021, 186, 1580–1590. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.M.; Eckert, D.; Carter, K.R.; Persson, M.; Warren, J.M. Springtime drought shifts carbon partitioning of recent photo-synthates in 10-year old Picea mariana trees, Causing restricted canopy development. Front. For. Glob. Change 2021, 3, 601046. [Google Scholar] [CrossRef]
- Duarte, A.G.; Katata, G.; Hoshika, Y.; Hossain, M.; Kreuzwieser, J.; Arneth, A.; Ruehr, N.K. Immediate and potential long-term effects of consecutive heat waves on the photosynthetic performance and water balance in Douglas-fir. J. Plant Physiol. 2016, 205, 57–66. [Google Scholar] [CrossRef]
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef]
- Held, M.; Ganthaler, A.; Lintunen, A.; Oberhuber, W.; Mayr, S. Tracheid and pit dimensions hardly vary in the xylem of Pinus sylvestris under contrasting growing conditions. Front. Plant Sci. 2021, 12, 786593. [Google Scholar] [CrossRef]
- Eldhuset, T.D.; Nagy, N.E.; Volařík, D.; Børja, I.; Gebauer, R.; Yakovlev, I.A.; Krokene, P. Drought affects tracheid structure, de-hydrin expression, and above- and belowground growth in 5-year-old Norway spruce. Plant Soil. 2013, 366, 305–320. [Google Scholar] [CrossRef]
- Xu, J.; Lu, J.; Evans, R.; Downes, G. Relationship between ring width and tracheid characteristics in Picea crassifolia: Implication in dendroclimatology. Bioresources 2014, 9, 2203–2213. [Google Scholar] [CrossRef] [Green Version]
- Chano, V.; Collada, C.; Soto, A. Transcriptomic analysis of wound xylem formation in Pinus canariensis. BMC Plant Biol. 2017, 17, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.L.; Roddy, A.B.; Brodersen, C.R.; McElrone, A.J.; Johnson, D.M. Anatomical and hydraulic responses to desiccation in emergent conifer seedlings. Am. J. Bot. 2020, 107, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Sena, J.S.; Giguere, I.; Rigault, P.; Bousquet, J.; Mackay, J. Expansion of the dehydrin gene family in the Pinaceae is associated with considerable structural diversity and drought-responsive expression. Tree Physiol. 2018, 38, 442–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigala, J.A.; Uscola, M.; Oliet, J.A.; Jacobs, D.F. Drought tolerance and acclimation in Pinus ponderosa seedlings: The influence of nitrogen form. Tree Physiol. 2020, 40, 1165–1177. [Google Scholar] [CrossRef]
- Lintunen, A.; Paljakka, T.; Jyske, T.; Peltoniemi, M.; Sterck, F.; von Arx, G.; Cochard, H.; Copini, P.; Caldeira, M.C.; Delzon, S.; et al. Osmolality and non-structural carbohydrate composition in the secondary phloem of trees across a latitudinal gradient in Europe. Front. Plant Sci. 2016, 7, 726. [Google Scholar] [CrossRef] [Green Version]
- Junker-Frohn, L.; Kleiber, A.; Jansen, K.; Gessler, A.; Kreuzwieser, J.; Ensminger, I. Differences in isoprenoid-mediated energy dissipation pathways between coastal and interior Douglas-fir seedlings in response to drought. Tree Physiol. 2019, 39, 1750–1766. [Google Scholar] [CrossRef]
- Kartashov, A.V.; Zlobin, I.E.; Pashkovskiy, P.P.; Pojidaeva, E.S.; Ivanov, Y.V.; Mamaeva, A.S.; Fesenko, I.A.; Kuznetsov, V.V. Quantitative analysis of differential dehydrin regulation in pine and spruce seedlings under water deficit. Plant Physiol. Biochem. 2021, 162, 237–246. [Google Scholar] [CrossRef]
- Papú, S.; Berli, F.; Piccoli, P.; Patón, D.; Ortega Rodriguez, D.R.; Roig, F.A. Physiological, biochemical, and anatomical responses of Araucaria araucana seedlings to controlled water restriction. Plant Physiol. Biochem. 2021, 165, 47–56. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees–from what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderegg, W.R.; Anderegg, L.D. Hydraulic and carbohydrate changes in experimental drought-induced mortality of saplings in two conifer species. Tree Physiol. 2013, 33, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earles, J.M.; Stevens, J.T.; Sperling, O.; Orozco, J.; North, M.P.; Zwieniecki, M.A. Extreme mid-winter drought weakens tree hy-draulic-carbohydrate systems and slows growth. New Phytol. 2018, 219, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Urrutia-Jalabert, R.; Lara, A.; Barichivich, J.; Vergara, N.; Rodriguez, C.G.; Piper, F.I. Low growth sensitivity and fast replenish-ment of non-structural carbohydrates in a long-lived endangered conifer after drought. Front. Plant Sci. 2020, 11, 905. [Google Scholar] [CrossRef] [PubMed]
- Galiano, L.; Timofeeva, G.; Saurer, M.; Siegwolf, R.; Martínez-Vilalta, J.; Hommel, R.; Gessler, A. The fate of recently fixed carbon after drought release: Towards unravelling C storage regulation in Tilia platyphyllos and Pinus sylvestris. Plant Cell Environ. 2017, 40, 1711–1724. [Google Scholar] [CrossRef]
- Piper, F.I.; Fajardo, A.; Hoch, G. Single-provenance mature conifers show higher non-structural carbohydrate storage and re-duced growth in a drier location. Tree Physiol. 2017, 37, 1001–1010. [Google Scholar] [CrossRef]
- Delgado, M.; Zúñiga-Feest, A.; Piper, F. Does carbon storage confer waterlogging tolerance? Evidence from four evergreen species of a temperate rainforest. Aust. J. Bot. 2018, 66, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Piper, F.; Paula, S. The role of nonstructural carbohydrates storage in forest resilience under climate change. Curr. For. Rep. 2020, 6, 1–13. [Google Scholar] [CrossRef]
- Zlobin, I.E.; Kartashov, A.V.; Pashkovskiy, P.P.; Ivanov, Y.V.; Kreslavski, V.D.; Kuznetsov, V.V. Comparative photosynthetic responses of Norway spruce and Scots pine seedlings to prolonged water deficiency. J. Photochem. Photobiol. B 2019, 201, 111659. [Google Scholar] [CrossRef]
- Kopaczyk, J.M.; Wargula, J.; Jelonek, T. The variability of terpenes in conifers under developmental and environmental stimuli. Environ. Exp. Bot. 2020, 180, 104197. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, Z.; Shi, Z.; Pan, L.; Kwon, S.; Yang, X.; Liu, Y. Tree characteristics and drought severity modulate the growth re-silience of natural Mongolian pine to extreme drought episodes. Sci. Total Environ. 2022, 830, 154742. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Cohen, S.; Paudel, I.; Preisler, Y.; Rotenberg, E.; Yakir, D. Diurnal dynamics of water transport, storage and hydraulic conductivity in pine trees under seasonal drought. iForest 2016, 9, 710–719. [Google Scholar] [CrossRef] [Green Version]
- Tomasella, M.; Häberle, K.-H.; Nardini, A.; Hesse, B.; Machlet, A.; Matyssek, R. Post-drought hydraulic recovery is accompanied by non-structural carbohydrate depletion in the stem wood of Norway spruce saplings. Sci. Rep. 2017, 7, 14308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laur, J.; Hacke, U.G. Exploring Picea glauca aquaporins in the context of needle water uptake and xylem refilling. New Phytol. 2014, 203, 388–400. [Google Scholar] [CrossRef]
- Earles, J.; Sperling, O.; Silva, L.; McElrone, A.; Brodersen, C.; North, M.; Zwieniecki, M. Bark water uptake promotes localized hydraulic recovery in coastal redwood crown. Plant Cell Environ. 2015, 39, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Torres-Ruiz, J.M.; Cochard, H.; Mencuccini, M.; Delzon, S.; Badel, E. Direct observation and modelling of embolism spread be-tween xylem conduits: A case study in Scots pine. Plant Cell Environ. 2016, 39, 2774–2785. [Google Scholar] [CrossRef]
- Guérin, M.; von Arx, G.; Martin-Benito, D.; Andreu-Hayles, L.; Griffin, K.L.; McDowell, N.G.; Pockman, W.; Gentine, P. Distinct xylem responses to acute vs prolonged drought in pine trees. Tree Physiol. 2020, 40, 605–620. [Google Scholar] [CrossRef] [Green Version]
- Klein, T.; Torres-Ruiz, J.M.; Albers, J.J. Conifer desiccation in the 2021 NW heatwave confirms the role of hydraulic damage. Tree Physiol. 2022, 42, 722–726. [Google Scholar] [CrossRef]
- Virlouvet, L.; Fromm, M. Physiological and transcriptional memory in guard cells during repetitive dehydration stress. New Phytol. 2015, 205, 596–607. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.L.; Speich, M.; Pappas, C.; Kahmen, A.; Von Arx, G.; Pannatier, E.G.; Steppe, K.; Treydte, K.; Stritih, A.; Fonti, P. Contrasting stomatal sensitivity to temperature and soil drought in mature alpine conifers. Plant Cell Environ. 2019, 42, 1674–1689. [Google Scholar] [CrossRef]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and promises of epigenetics for forest trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Matallana-Ramirez, L.P.; Whetten, R.W.; Sanchez, G.M.; Payn, K.G. Breeding for Climate Change Resilience: A Case Study of Loblolly Pine (Pinus taeda L.) in North America. Front. Plant Sci. 2021, 12, 606908. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Proutsos, N.; Tigkas, D. Growth response of endemic black pine trees to meteorological variations and drought episodes in a mediterranean region. Atmosphere 2020, 11, 554. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Andivia, E.; Candel-Pérez, D.; Linares, J.; Camarero, J. Long term forest management drives drought resilience in Mediterranean black pine forest. Trees 2021, 35, 1651–1662. [Google Scholar] [CrossRef]
- Manrique-Alba, À.; Beguería, S.; Molina, A.J.; González-Sanchis, M.; Tomàs-Burguera, M.; del Campo, A.D.; Colangelo, M.; Camarero, J.J. Long-term thinning effects on tree growth, drought response and water use efficiency at two Aleppo pine plantations in Spain. Sci. Total Environ. 2020, 728, 138536. [Google Scholar] [CrossRef]
- Duque Lazo, J.; Navarro-Cerrillo, R.; Sanchez-Salguero, R.; Rodriguez Vallejo, C. Is thinning an alternative when trees could die in response to drought? The case of planted Pinus nigra and P. Sylvestris stands in southern Spain. Forest Ecol. Manag. 2019, 433, 313–324. [Google Scholar] [CrossRef]
- Sun, S.; Lei, S.; Jia, H.; Li, C.; Zhang, J.; Meng, P. Tree-ring analysis reveals density-dependent vulnerability to drought in planted mongolian pines. Forests 2020, 11, 98. [Google Scholar] [CrossRef]
- Andivia, E.; Ruiz-Benito, P.; Díaz-Martínez, P.; Carro-Martínez, N.; Zavala, M.A.; Madrigal-González, J. Inter-specific tolerance to recurrent droughts of pine species revealed in saplings rather than adult trees. Forest Ecol. Manag. 2020, 459, 117848. [Google Scholar] [CrossRef]
- Manrique-Alba, À.; Sevanto, S.; Adams, H.D.; Collins, A.D.; Dickman, L.T.; Chirino, E.; Bellot, J.; McDowell, N.G. Stem radial growth and water storage responses to heat and drought vary between conifers with differing hydraulic strategies. Plant Cell Environ. 2018, 41, 1926–1934. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Schütze, G.; Biber, P. Drought can favour the growth of small in relation to tall trees in mature stands of Norway spruce and European beech. For. Ecosyst. 2018, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Venturas, M.D.; Sperry, J.S.; Hacke, U.G. Plant xylem hydraulics: What we understand, current research, and future challenges. J. Integr. Plant Biol. 2017, 59, 356–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagneri, D.; Fonti, P.; von Arx, G.; Carrer, M. How does climate influence xylem morphogenesis over the growing season? Insights from long-term intra-ring anatomy in Picea abies. Ann. Bot. 2017, 119, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Martin-Benito, D.; Anchukaitis, K.J.; Evans, M.N.; Del Río, M.; Beeckman, H.; Cañellas, I. Effects of drought on xylem anatomy and water-use efficiency of two co-occurring pine species. Forests 2017, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Rosner, S.; Gierlinger, N.; Klepsch, M.; Karlsson, B.; Evans, R.; Lundqvist, S.-O.; Světlík, J.; Børja, I.; Dalsgaard, L.; Andreassen, K.; et al. Hydraulic and mechanical dysfunction of Norway spruce sapwood due to extreme summer drought in Scandinavia. Forest Ecol. Manag. 2018, 409, 527–540. [Google Scholar] [CrossRef]
- Chen, Z.; Karlsson, B.; Mörling, T.; Olsson, L.; Mellerowicz, E.; Wu, H.; Lundqvist, S.-O.; Garcia Gil, R. Genetic analysis of fiber dimensions and their correlation with stem diameter and solid-wood properties in Norway spruce. Tree Genet. Genomes 2016, 12, 123. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Garcia Gil, R.; Karlsson, B.; Lundqvist, S.-O.; Olsson, L.; Wu, H. Inheritance of growth and solid wood quality traits in a large Norway spruce population tested at two locations in southern Sweden. Tree Genet. Genomes 2014, 10, 1291–1303. [Google Scholar] [CrossRef]
- Fajstavr, M.; Bednářová, E.; Nezval, O.; Giagli, K.; Gryc, V.; Vavrčík, H.; Horacek, P.; Urban, J. How needle phenology indicates the changes of xylem cell formation during drought stress in Pinus sylvestris L. Dendrochronologia 2019, 56, 125600. [Google Scholar] [CrossRef]
- Larysch, E.; Stangler, D.F.; Nazari, M.; Seifert, T.; Kahle, H.P. Xylem phenology and growth response of European beech, Silver fir and Scots pine along an elevational gradient during the extreme drought year 2018. Forests 2021, 12, 75. [Google Scholar] [CrossRef]
- Saderi, M.; Rathgeber, C.; Rozenberg, P.; Fournier, M. Phenology of wood formation in larch (Larix decidua Mill.) trees growing along a 1000-m elevation gradient in the French Southern Alps. Ann. For. Sci. 2019, 76, 89. [Google Scholar] [CrossRef]
- Leites, L.P.; Rehfeldt, G.E.; Robinson, A.P.; Crookston, N.L.; Jaquish, B. Possibilities and limitations of using historic provenance tests to infer forest species growth responses to Climate Change. Nat. Resour. Model. 2012, 25, 409–433. [Google Scholar] [CrossRef]
- Capblancq, T.; Lachmuth, S.; Fitzpatrick, M.C.; Keller, S.R. From common gardens to candidate genes: Exploring local adaptation to climate in red spruce. New Phytol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, G.E.; Jaquish, B.; Saenz-Romero, C.; Joyce, D.; Leites, L.; Clair, J.; Upton, J. Comparative genetic responses to climate in the varieties of Pinus ponderosa and Pseudotsuga menziesii: Reforestation. Forest Ecol. Manag. 2014, 324, 147–157. [Google Scholar] [CrossRef]
- Mihai, G.; Alexandru, A.; Stoica, E.; Birsan, M.V. Intraspecific growth response to drought of Abies alba in the southeastern Car-pathians. Forests 2021, 12, 387. [Google Scholar] [CrossRef]
- Depardieu, C.; Girardin, M.P.; Nadeau, S.; Lenz, P.; Bousquet, J.; Isabel, N. Adaptive genetic variation to drought in a widely distributed conifer suggests a potential for increasing forest resilience in a drying climate. New Phytol. 2020, 227, 427–439. [Google Scholar] [CrossRef] [Green Version]
- Pardos, M.; Calama, R. Responses of Pinus pinea seedlings to moderate drought and shade: Is the provenance a differential factor? Photosynthetica 2017, 56, 786–798. [Google Scholar] [CrossRef]
- Warwell, M.V.; Shaw, R.G. Phenotypic selection on ponderosa pine seed and seedling traits in the field under three experimen-tally manipulated drought treatments. Evol. Appl. 2019, 12, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Vizcaíno-Palomar, N.; González-Muñoz, N.; González-Martínez, S.; Alia, R.; Garzón, M. Most southern Scots pine populations are locally adapted to drought for tree height growth. Forests 2019, 10, 555. [Google Scholar] [CrossRef]
- Budeanu, M.; Apostol, E.; Besliu, E.; Crișan, V.; Petritan, A.M. Phenotypic Variability and differences in the drought response of Norway spruce pendula and pyramidalis half-sib families. Forests 2021, 12, 947. [Google Scholar] [CrossRef]
- Schueler, S.; George, J.-P.; Karanitsch-Ackerl, S.; Mayer, K.; Klumpp, R.T.; Grabner, M. Evolvability of drought response in four native and non-native conifers: Opportunities for forest and genetic resource management in Europe. Front. Plant Sci. 2021, 12, 1304. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Ghazghazi, H.; Riahi, L.; Yangui, I.; Messaoud, C.; Rzigui, T.; Nasr, Z. Effect of drought stress on physio-biochemical traits and secondary metabolites production in the woody species Pinus halepensis Mill. at a juvenile development stage. J. Sustain. Forest 2022, 41, 878–894. [Google Scholar] [CrossRef]
- Chaves, M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2008, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, D.E.; Chaitanya, V. Photosynthesis and antioxidative defense mechanisms in deciphering drought stress tolerance of crop plants. Biol. Plant. 2016, 60, 201–218. [Google Scholar] [CrossRef]
- Drake, J.E.; Power, S.A.; Duursma, R.A.; Medlyn, B.E.; Aspinwall, M.J.; Choat, B.; Creek, D.; Eamusc, D.; Maier, C.; Pfautsch, S.; et al. Stomatal and non-stomatal limitations of photosynthesis for four tree species under drought: A comparison of model formulations. Agric. For. Meteorol. 2017, 247, 454–466. [Google Scholar] [CrossRef]
- Malenovský, Z.; Homolova, L.; Zurita-Milla, R.; Lukeš, P.; Kaplan, V.; Hanus, J.; Gastellu-Etchegorry, J.P.; Schaepman, M. Re-trieval of spruce leaf chlorophyll content from airborne image data using continuum removal and radiative transfer. Remote Sens. Environ. 2013, 131, 85–102. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Wang, S.Q.; Chen, B.; Li, Y.; Amir, M.; Ma, L.; Zhu, K.; Yang, F.T.; Wang, X.B.; Liu, Y.Y.; et al. Comparative analysis on the estimation of diurnal solar-induced chlorophyll fluorescence dynamics for a subtropical evergreen coniferous forest. Remote Sens. 2021, 13, 3143. [Google Scholar] [CrossRef]
- Lausch, A.; Borg, E.; Bumberger, J.; Dietrich, P.; Heurich, M.; Huth, A.; Jung, A.; Klenke, R.; Knapp, S.; Mollenhauer, H.; et al. Un-derstanding forest health with remote sensing, Part III: Requirements for a scalable multi-source forest health monitoring net-work based on data science approaches. Remote Sens. 2018, 10, 1120. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Bigras, F. Photosynthetic response of white spruce families to drought stress. New For. 2005, 29, 135–148. [Google Scholar] [CrossRef]
- Maseyk, K.; Lin, T.; Cochavi, A.; Schwartz, A.; Yakir, D. Quantification of leaf-scale light energy allocation and photoprotection processes in a Mediterranean pine forest under extensive seasonal drought. Tree Physiol. 2019, 39, 1767–1782. [Google Scholar] [CrossRef] [PubMed]
- Zuromski, L.M.; Bowling, D.R.; Kohler, P.; Frankenberg, C.; Goulden, M.L.; Blanken, P.D.; Lin, J.C. Solar-induced fluorescence detects interannual variation in gross primary production of coniferous forests in the western United States. Geophys. Res. Lett. 2018, 45, 7184–7193. [Google Scholar] [CrossRef]
- Matisons, R.; Krisans, O.; Jansons, A.; Kondratovičs, T.; Elferts, D.; Ievinsh, G. Norway spruce seedlings from an eastern Baltic provenance show tolerance to simulated drought. Forests 2021, 12, 82. [Google Scholar] [CrossRef]
- Konôpková, A.; Húdoková, H.; Ježík, M.; Kurjak, D.; Jamnická, G.; Ditmarová, Ľ.; Gomory, D.; Longauer, R.; Tognetti, R.; Pšidová, E. Origin rather than mild drought stress influenced chlorophyll a fluorescence in contrasting silver fir (Abies alba Mill.) prove-nances. Photosynthetica 2020, 58, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Voltas, J.; Chambel, M.; Prada, A.; Ferrio, J.P. Climate-related variability in carbon and oxygen stable isotopes among popula-tions of Aleppo pine grown in common-garden tests. Trees 2008, 22, 759–769. [Google Scholar] [CrossRef]
- Klein, T.; Matteo, G.; Rotenberg, E.; Cohen, S.; Yakir, D. Differential ecophysiological response of a major Mediterranean pine species across a climatic gradient. Tree Physiol. 2012, 33, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhang, F.; Gou, X.H.; Fonti, P.; Xia, J.Q.; Cao, Z.Y.; Liu, J.G.; Wang, Y.F.; Zhang, J.Z. Seasonal variations in leaf-level photosynthesis and water use efficiency of three isohydric to anisohydric conifers on the Tibetan Plateau. Agric. For. Meteorol. 2021, 308, 108581. [Google Scholar] [CrossRef]
- De Miguel, M.; Cabezas, J.-A.; de María, N.; Sánchez-Gómez, D.; Guevara, M.-Á.; Vélez, M.-D.; Sáez-Laguna, E.; Díaz, L.-M.; Man-cha, J.-A.; Barbero, M.-C.; et al. Genetic control of functional traits related to photosynthesis and water use efficiency in Pinus pi-naster Ait. drought response: Integration of genome annotation, allele association and QTL detection for candidate gene identification. BMC Genom. 2014, 15, 464. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Guy, R.D.; Holowachuk, D.L. Population differences in stable carbon isotope ratio of Pinus contorta Dougl. ex Loud.: Relationship to environment, climate of origin, and growth potential. Can. J. Bot. 2001, 79, 274–283. [Google Scholar] [CrossRef]
- Csilléry, K.; Buchmann, N.; Fady, B. Adaptation to drought is coupled with slow growth, but independent from phenology in marginal silver fir (Abies alba Mill.) populations. Evol. Appl. 2020, 13, 2357–2376. [Google Scholar] [CrossRef] [PubMed]
- Birami, B.; Gattmann, M.; Heyer, A.G.; Grote, R.; Arneth, A.; Ruehr, N.K. Heat waves alter carbon allocation and increase mortality of Aleppo pine under dry conditions. Front. For. Glob. Change 2018, 1, 8. [Google Scholar] [CrossRef]
- Plesa, I.; Al Hassan, M.; González Orenga, S.; Sestras, A.; Vicente, O.; Prohens, J.; Boscaiu, M.; Sestras, R. Responses to drought in seedlings of European larch (Larix decidua Mill.) from several carpathian provenances. Forests 2019, 10, 511. [Google Scholar] [CrossRef] [Green Version]
- Palacio, S.; Hoch, G.; Sala, A.; Körner, C.; Millard, P. Does carbon storage limit tree growth? New Phytol. 2014, 201, 1096–1100. [Google Scholar] [CrossRef] [Green Version]
- Klein, T.; Hoch, G.; Yakir, D.; Körner, C. Drought stress, growth and nonstructural carbohydrate dynamics of pine trees in a semi-arid forest. Tree Physiol. 2014, 34, 981–992. [Google Scholar] [CrossRef] [Green Version]
- Sala, A.; Woodruff, D.; Meinzer, F. Carbon dynamics in trees: Feast or famine? Tree Physiol. 2012, 32, 764–775. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Moran, E.; Lauder, J.; Musser, C.; Stathos, A.; Shu, M. The genetics of drought tolerance in conifers. New Phytol. 2017, 216, 1034–1048. [Google Scholar] [CrossRef]
- Fox, H.; Doron-Faigenboim, A.; Kelly, G.; Bourstein, R.; Attia, Z.; Zhou, J.; Moshe, Y.; Moshelion, M.; David-Schwartz, R. Tran-scriptome analysis of Pinus halepensis under drought stress and during recovery. Tree Physiol. 2018, 38, 423–441. [Google Scholar] [CrossRef] [Green Version]
- De María, N.; Guevara, M.Á.; Perdiguero, P.; Vélez, D.; Cabezas, J.A.; López-Hinojosa, M.; Li, Z.; Díaz, L.M.; Pizarro, A.; Mancha, J.; et al. Molecular study of drought response in the Mediterranean conifer Pinus pinaster Ait.: Differential transcriptomic profiling reveals constitutive water deficit-independent drought tolerance mechanisms. Ecol. Evol. 2020, 10, 9788–9807. [Google Scholar] [CrossRef]
- Li, J.; West, J.B.; Hart, A.; Wegrzyn, J.L.; Smith, M.A.; Domec, J.-C.; Loopstra, C.A.; Casola, C. Extensive variation in drought-induced gene expression changes between loblolly pine genotypes. Front. Genet. 2021, 12, 661440. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, T.; Liu, S.-W.; Uddin, S.; Amjid, M.W.; Niu, S.-H.; Wu, H.X. The transcriptional landscape and hub genes associated with physiological responses to drought stress in Pinus tabuliformis. Int. J. Mol. Sci. 2021, 22, 9604. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Zhao, Y.; Wang, X.R.; Liu, Q.; Ran, J. Transcriptome analysis of needle and root of Pinus massoniana in response to con-tinuous drought stress. Plants 2021, 10, 769. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hu, X.G.; Zheng, B.; Li, Y.; Wang, T.; Sharma, A.; Yuan, H.W.; Mao, J.F. Transcriptome-wide Identification and Characterization of microRNAs and Their Targets in a Highly Adaptable Conifer Platycladus orientalis. J. Am. Soc. Hortic. Sci. 2022, 147, 7–17. [Google Scholar] [CrossRef]
- Carvalho, A.; Pavia, I.; Fernandes, C.; Pires, J.; Correia, C.; Bacelar, E.; Moutinho-Pereira, J.; Gaspar, M.J.; Bento, J.; Silva, M.E.; et al. Differential physiological and genetic responses of five European Scots pine provenances to induced water stress. J. Plant Physiol. 2017, 215, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Conde, T.; Yakovlev, I.; Majada, J.; Aranda, I.; Johnsen, Ø. Dehydrins in maritime pine (Pinus pinaster) and their expres-sion related to drought stress response. Tree Genet. Genomes 2012, 8, 957–973. [Google Scholar] [CrossRef] [Green Version]
- Behringer, D.; Zimmermann, H.; Ziegenhagen, B.; Liepelt, S. Differential gene expression reveals candidate genes for drought stress response in Abies alba (Pinaceae). PLoS ONE 2015, 10, e0124564. [Google Scholar] [CrossRef]
- Li, W.; Lee, J.; Yu, S.; Wang, F.; Lv, W.; Zhang, X.; Li, C.; Yang, J. Characterization and analysis of the transcriptome response to drought in Larix kaempferi using PacBio full-length cDNA sequencing integrated with de novo RNA-seq reads. Planta 2021, 253, 28. [Google Scholar] [CrossRef]
- Du, M.Z.; Ding, G.; Cai, Q.-Z. The transcriptomic responses of Pinus massoniana to drought stress. Forests 2018, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, W.; Alba, R.; Yu, Y.-S.; Bordeaux, J.; Simões, M.; Dean, J. Microarray analysis and scale-free gene networks identify can-didate regulators in drought-stressed roots of loblolly pine (P. taeda L.). BMC Genom. 2011, 12, 264. [Google Scholar] [CrossRef] [Green Version]
- Perdiguero, P.; Collada, C.; Barbero, M.; Casado, G.; Cervera, M.T.; Soto, A. Identification of water stress genes in Pinus pinaster Ait. by controlled progressive stress and suppression-subtractive hybridization. Plant Physiol. Biochem. 2012, 50, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Watkinson, J.; Sioson, A.; Vasquez-Robinet, C.; Shukla, M.; Kumar, D.; Ellis, M.; Heath, L.; Ramakrishnan, N.; Chevone, B.; Wat-son, L.; et al. Photosynthetic acclimation is reflected in specific patterns of gene expression in drought-stressed loblolly pine. Plant Physiol. 2004, 133, 1702–1716. [Google Scholar] [CrossRef] [Green Version]
- Hamanishi, E.; Campbell, M. Genome-wide responses to drought in forest trees. Forestry 2011, 84, 273–283. [Google Scholar] [CrossRef]
- Perdiguero, P.; Barbero, M.; Cervera, M.T.; Soto, A.; Collada, C. Novel conserved segments are associated with differential ex-pression patterns for Pinaceae dehydrins. Planta 2012, 236, 1863–1874. [Google Scholar] [CrossRef] [PubMed]
- Van Ghelder, C.; Parent, G.J.; Rigault, P.; Prunier, J.; Giguere, I.; Caron, S.; Sena, J.S.; Deslauriers, A.; Bousquet, J.; Esmenjaud, D.; et al. The large repertoire of conifer NLR resistance genes includes drought responsive and highly diversified RNLs. Sci. Rep. 2019, 9, 11614. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Schoettle, A.W.; Sniezko, R.A.; Williams, H.; Zamany, A.; Rancourt, B. Fine dissection of limber pine resistance to Cronartium ribicola using targeted sequencing of the NLR family. BMC Genom. 2021, 22, 567. [Google Scholar] [CrossRef]
- González-Martínez, S.; Krutovsky, K.; Neale, D. Forest-tree population genomics and adaptive evolution. New Phytol. 2006, 170, 227–238. [Google Scholar] [CrossRef]
- Gonzalez-Martinez, S.C.; Wheeler, N.C.; Ersoz, E.; Nelson, C.D.; Neale, D.B. Association genetics in Pinus taeda L. I. Wood prop-erty traits. Genetics 2007, 175, 399–409. [Google Scholar] [CrossRef]
- Neale, D.; Kremer, A. Forest tree genomics: Growing resources and applications. Nature reviews. Genetics 2011, 12, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Myles, S.; Peiffer, J.; Brown, P.; Ersoz, E.; Zhang, Z.; Costich, D.; Buckler, E. Association mapping: Critical considerations shift from genotyping to experimental design. Plant Cell 2009, 21, 2194–2202. [Google Scholar] [CrossRef] [Green Version]
- Cumbie, W.P.; Eckert, A.; Wegrzyn, J.; Whetten, R.; Neale, D.; Goldfarb, B. Association genetics of carbon isotope discrimination, height and foliar nitrogen in a natural population of Pinus taeda L. Heredity 2011, 107, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depardieu, C.; Gérardi, S.; Nadeau, S.; Parent, G.; MacKay, J.; Lenz, P.; Lamothe, M.; Girardin, M.; Bousquet, J.; Isabel, N. Con-necting tree-ring phenotypes, genetic associations, and transcriptomics to decipher the genomic architecture of drought adapta-tion in a widespread conifer. Mol. Ecol. 2021, 30, 3898–3917. [Google Scholar] [CrossRef] [PubMed]
- Eckert, A.; Heerwaarden, J.; Wegrzyn, J.; Nelson, C.; Ross-Ibarra, J.; González-Martínez, S.; Neale, D. Patterns of population structure and environmental associations to aridity across the range of loblolly pine (Pinus taeda L., Pinaceae). Genetics 2010, 185, 969–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peláez, P.; Ortiz Martínez, A.; Montes, R.; Gernandt, D. Population structure, diversifying selection, and local adaptation in Pinus patula. Am. J. Bot. 2020, 107, 1555–1566. [Google Scholar] [CrossRef]
- Trujillo Moya, C.; George, J.-P.; Fluch, S.; Geburek, T.; Grabner, M.; Karanitsch-Ackerl, S.; Konrad, H.; Mayer, K.; Sehr, E.; Wischnitzki, E.; et al. Drought sensitivity of Norway spruce at the species’ warmest fringe: Quantitative and molecular analysis reveals high genetic variation among and within provenances. G3-Genes Genom. Genet. 2018, 8, 1225–1245. [Google Scholar] [CrossRef] [Green Version]
- Vangestel, C.; Vazquez-Lobo, A.; Martínez-García, P.J.; Ćalić, I.; Wegrzyn, J.; Neale, D. Patterns of neutral and adaptive genetic diversity across the natural range of sugar pine (Pinus lambertiana Dougl.). Tree Genet. Genom. 2016, 12, 51. [Google Scholar] [CrossRef]
- De La Torre, A.R.; Sekhwal, M.K.; Puiu, D.; Salzberg, S.L.; Scott, A.D.; Allen, B.; Neale, D.B.; Chin, A.R.O.; Buckley, T.N. Ge-nome-wide association identifies candidate genes for drought tolerance in coast redwood and giant sequoia. Plant J. 2022, 109, 7–22. [Google Scholar] [CrossRef]
- De La Torre, A.; Birol, I.; Bousquet, J.; Ingvarsson, P.; Jansson, S.; Jones, S.; Keeling, C.; MacKay, J.; Nilsson, O.; Ritland, K.; et al. Insights into conifer giga-genomes. Plant Physiol. 2014, 166, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Karam, M.-J.; Lefèvre, F.; Bou Dagher Kharrat, M.; Pinosio, S.; Giovanni Giuseppe, V. Genomic exploration and molecular mark-er development in a large and complex conifer genome using RADseq and mRNAseq. Mol. Ecol. Res. 2014, 15, 601–612. [Google Scholar] [CrossRef]
- Pan, J.; Wang, B.; Pei, Z.Y.; Zhao, W.; Gao, J.; Mao, J.-F.; Wang, X.-R. Optimization of the genotyping-by-sequencing strategy for population genomic analysis in conifers. Mol. Ecol. Res. 2014, 15, 711–722. [Google Scholar] [CrossRef]
- Hrivnák, M.; Krajmerová, D.; Kurjak, D.; Konôpková, A.; Magni, F.; Scaglione, D.; Ditmarová, L.; Jamnická, G.; Marešová, J.; Gömöry, D. Differential associations between nucleotide polymorphisms and physiological traits in Norway spruce (Picea abies Karst.) plants under contrasting water regimes. Forestry 2022, 95, 686–697. [Google Scholar] [CrossRef]
- Neves, L.G.; Davis, J.M.; Barbazuk, W.B.; Kirst, M. Whole-exome targeted sequencing of the uncharacterized pine genome. Plant J. 2013, 75, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Capblancq, T.; Butnor, J.R.; Deyoung, S.; Thibault, E.; Munson, H.; Nelson, D.M.; Fitzpatrick, M.C.; Keller, S.R. Whole-exome se-quencing reveals a long-term decline in effective population size of red spruce (Picea rubens). Evol. Appl. 2020, 13, 2190–2205. [Google Scholar] [CrossRef]
- George, J.-P.; Schueler, S.; Grabner, M.; Karanitsch-Ackerl, S.; Stierschneider, M.; Weissenbacher, L.; Loo, M. Looking for the needle in a downsized haystack: Whole-exome sequencing unravels how selection and gene flow have shaped climatic adaptation in Douglas-fir (Pseudotsuga menziesii). Ecol. Evol. 2021, 17, 8238–8253. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Seeve, C.; Loopstra, C.; Krutovsky, K. Exploring the genetic basis of gene transcript abundance and metabolite levels in loblolly pine (Pinus taeda L.) using association mapping and network construction. BMC Genet. 2018, 19, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plomion, C.; Bousquet, J.; Kole, C. Genetics, Genomics and Breeding of Conifers; CRC Press and Edenbridge Science Publishers: New York, NY, USA, 2011; p. 449. ISBN 978-1-57808-719-8. [Google Scholar]
- Wei, J.; Pei, X.; Hu, X.; Sun, S.; Zhao, C.; Han, R.; Zhao, X. Applications of transcriptome in conifer species. Plant Cell Tissue Organ Cult. 2022, 150, 511–525. [Google Scholar] [CrossRef]
- Cappa, E.P.; Klutsch, J.G.; Sebastian-Azcona, J.; Ratcliffe, B.; Wei, X.; Da Ros, L.; Liu, Y.; Chen, C.; Benowicz, A.; Sadoway, S.; et al. Integrating genomic information and productivity and climate-adaptability traits into a regional white spruce breeding program. PLoS ONE 2022, 17, e0264549. [Google Scholar] [CrossRef]
- Laverdière, J.P.; Lenz, P.; Nadeau, S.; Depardieu, C.; Isabel, N.; Perron, M.; Beaulieu, J.; Bousquet, J. Breeding for adaptation to climate change: Genomic selection for drought response in a white spruce multi-site polycross test. Evol. Appl. 2022, 15, 383–402. [Google Scholar] [CrossRef]
- Mullin, T.; Andersson Gull, B.; Bastien, J.C.; Beaulieu, J.; Burdon, R.; Dvorak, W.; King, J.; Kondo, T.; Krakowski, J.; Lee, S.; et al. Economic importance, breeding objectives and achievements. In Genetics, Genomics and Breeding of Conifers; Plomion, C., Bousquet, J., Kole, C., Eds.; Science Publishers and CRC Press: New York, NY, USA, 2011; pp. 40–127. ISBN 9781578087198. [Google Scholar]
- Reyer, C.; Brouwers, N.; Rammig, A.; Brook, B.; Epila, J.; Grant, R.F.; Holmgren, M.; Langerwisch, F.; Leuzinger, S.; Lucht, W.; et al. Forest resilience and tipping points at different spatio-temporal scales: Approaches and challenges. J. Ecol. 2015, 103, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Zobel, B.; Talbert, J. Applied Forest Tree Improvement; Blackburn Press: Caldwel, NJ, USA, 2003; ISBN 1930665814. [Google Scholar]
- Esposito, S.; Carputo, D.; Cardi, T.; Tripodi, P. Applications and trends of machine learning in genomics and phenomics for next-generation breeding. Plants 2019, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Sun, D.; Cen, H.; Haixia, X.; Haiyong, W.; Yuan, F. Phenotyping of arabidopsis drought stress response using kinetic chlorophyll fluorescence and multicolor fluorescence imaging. Front. Plant Sci. 2018, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Lowe, A.; Harrison, N.; French, A. Hyperspectral image analysis techniques for the detection and classification of the early onset of plant disease and stress. Plant Methods 2017, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Birol, I.; Raymond, A.; Jackman, S.D.; Pleasance, S.; Coope, R.; Taylor, G.A.; Yuen, M.M.; Keeling, C.I.; Brand, D.; Vandervalk, B.P.; et al. Assembling the 20 Gb white spruce (Picea glauca) genome from whole-genome shotgun sequencing data. Bioinformatics 2013, 29, 1492–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nystedt, B.; Street, N.; Wetterbom, A.; Zuccolo, A.; Lin, Y.-C.; Scofield, D.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, K.; Wegrzyn, J.; Zimin, A.; Puiu, D.; Crepeau, M.; Cardeno, C.; Paul, R.; Gonzalez-Ibeaz, D.; Koriabine, M.; Holtz-Morris, A.; et al. Sequence of the sugar pine megagenome. Genetics 2016, 204, 1613–1626. [Google Scholar] [CrossRef] [Green Version]
- Zimin, A.; Stevens, K.A.; Crepeau, M.W.; Holtz-Morris, A.; Koriabine, M.; Marçais, G.; Puiu, D.; Roberts, M.; Wegrzyn, J.L.; de Jong, P.J.; et al. Sequencing and assembly of the 22-Gb loblolly pine genome. Genetics 2014, 196, 875–890. [Google Scholar] [CrossRef] [Green Version]
- Scott, A.D.; Zimin, A.V.; Puiu, D.; Workman, R.; Britton, M.; Zaman, S.; Caballero, M.; Read, A.C.; Bogdanove, A.J.; Burns, E.; et al. A reference genome sequence for giant sequoia. G3-Genes Genom. Genet. 2020, 10, 3907–3919. [Google Scholar] [CrossRef]
- Niu, S.H.; Li, J.; Bo, W.H.; Yang, W.F.; Zuccolo, A.; Giacomello, S.; Chen, X.; Han, F.X.; Yang, J.H.; Song, Y.; et al. The Chinese pine genome and methylome unveil key features of conifer evolution. Cell 2022, 185, 204–217. [Google Scholar] [CrossRef]
- Traversari, S.; Giovannelli, A.; Emiliani, G. Wood formation under changing environment: Omics approaches to elucidate the mechanisms driving the early-to-latewood transition in Conifers. Forests 2022, 13, 608. [Google Scholar] [CrossRef]
- Pavy, N.; Lamothe, M.; Pelgas, B.; Gagnon, F.; Birol, I.; Bohlmann, J.; Mackay, J.; Isabel, N.; Bousquet, J. A high-resolution refer-ence genetic map positioning 8.8K genes for the conifer white spruce: Structural genomics implications and correspondence with physical distance. Plant J. 2017, 90, 189–203. [Google Scholar] [CrossRef] [Green Version]
- Bernhardsson, C.; Vidalis, A.; Wang, X.; Scofield, D.G.; Schiffthaler, B.; Baison, J.; Street, N.R.; Garcia-Gil, M.R.; Ingvarsson, P.K. An ultra-dense haploid genetic map for evaluating the highly fragmented genome assembly of Norway spruce (Picea abies). G3-Genes Genom. Genet. 2019, 9, 1623–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, M.L.; He, Q.W.; Zhao, J.; Zhang, Y.; Yuan, D.S.; Zhang, J.F. Genetic mapping of Prince Rupprecht’s larch (Larix princi-pis-rupprechtii Mayr) by specific-locus amplified fragment sequencing. Genes 2019, 10, 583. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.Q.; Zhao, W.; Nie, S.; Liu, S.S.; El-Kassaby, Y.A.; Wang, X.R.; Mao, J.F. Genome-wide variant identification and high-density genetic map construction using RADseq for Platycladus orientalis (Cupressaceae). G3-Genes Genom. Genet. 2019, 9, 3663–3672. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Schoettle, A.W.; Sniezko, R.A.; Yao, F.P.; Zamany, A.; Williams, H.; Rancourt, B. Limber pine (Pinus flexilis James) genetic map constructed by exome-seq provides insight into the evolution of disease resistance and a genomic resource for ge-nomics-based breeding. Plant J. 2019, 98, 745–758. [Google Scholar] [CrossRef]
- Pavy, N.; Gagnon, F.; Rigault, P.; Blais, S.; Deschenes, A.; Boyle, B.; Pelgas, B.; Deslauriers, M.; Clement, S.; Lavigne, P.; et al. Development of high-density SNP genotyping arrays for white spruce (Picea glauca) and transferability to subtropical and nordic congeners. Mol. Ecol. Resour. 2013, 13, 324–336. [Google Scholar] [CrossRef]
- Telfer, E.; Graham, N.; Macdonald, L.; Li, Y.J.; Klapste, J.; Resende, M.; Neves, L.G.; Dungey, H.; Wilcox, P. A high-density exome capture genotype-by-sequencing panel for forestry breeding in Pinus radiata. PLoS ONE 2019, 14, e02222640. [Google Scholar] [CrossRef] [Green Version]
- Nagano, S.; Hirao, T.; Takashima, Y.; Matsushita, M.; Mishima, K.; Takahashi, M.; Iki, T.; Ishiguri, F.; Hiraoka, Y. SNP Genotyping with target amplicon sequencing using a multiplexed primer panel and its application to genomic prediction in Japanese cedar, Cryptomeria japonica(L.f.) D.Don. Forests 2020, 11, 898. [Google Scholar] [CrossRef]
- Bernhardsson, C.; Zan, Y.J.; Chen, Z.Q.; Ingvarsson, P.K.; Wu, H.X. Development of a highly efficient 50K single nucleotide pol-ymorphism genotyping array for the large and complex genome of Norway spruce (Picea abies L. Karst) by whole genome rese-quencing and its transferability to other spruce species. Mol. Ecol. Resour. 2021, 21, 880–896. [Google Scholar] [CrossRef]
- Kastally, C.; Niskanen, A.K.; Perry, A.; Kujala, S.T.; Avia, K.; Cervantes, S.; Haapanen, M.; Kesalahti, R.; Kumpula, T.A.; Mattila, T.M.; et al. Taming the massive genome of Scots pine with PiSy50k, a new genotyping array for conifer research. Plant J. 2022, 109, 1337–1350. [Google Scholar] [CrossRef]
- Muranty, H.; Jorge, V.; Bastien, C.; Lepoittevin, C.; Bouffier, L.; Sanchez, L. Potential for marker-assisted selection for forest tree breeding: Lessons from 20 years of MAS in crops. Tree Genet. Genomes 2014, 10, 1491–1510. [Google Scholar] [CrossRef] [Green Version]
- Grattapaglia, D.; Silva-Junior, O.B.; Resende, R.T.; Cappa, E.P.; Muller, B.S.F.; Tan, B.Y.; Isik, F.; Ratcliffe, B.; El-Kassaby, Y.A. Quantitative genetics and genomics converge to accelerate forest tree breeding. Front. Plant Sci. 2018, 9, 1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingvarsson, P.K.; Street, N.R. Association genetics of complex traits in plants. New Phytol. 2011, 189, 909–922. [Google Scholar] [CrossRef] [PubMed]
- Grattapaglia, D. Status and perspectives of genomic selection in forest tree breeding. In Genomic Selection for Crop Improvement: New Molecular Breeding Strategies for Crop Improvement; Varshney, R.K., Roorkiwal, M., Sorrells, M.E., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 199–249. [Google Scholar] [CrossRef]
- Meuwissen, T.H.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Isik, F. Genomic selection in forest tree breeding: The concept and an outlook to the future. New For. 2014, 45, 379–401. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Lebedeva, T.N.; Chernodubov, A.I.; Shestibratov, K.A. Genomic selection for forest tree improvement: Methods, achievements and perspectives. Forests 2020, 11, 1190. [Google Scholar] [CrossRef]
- Isik, F.; Bartholomé, J.; Farjat, A.; Chancerel, E.; Raffin, A.; Sanchez, L.; Plomion, C.; Bouffier, L. Genomic selection in maritime pine. Plant Sci. 2016, 242, 108–119. [Google Scholar] [CrossRef]
- Li, Z.Y.; Suontama, M.; Burdon, R.D.; Dungey, H.S. Genotype by environment interactions in forest tree breeding: Review of methodology and perspectives on research and application. Tree Genet. Genom. 2017, 13, 60. [Google Scholar] [CrossRef]
- Zapata-Valenzuela, J.; Whetten, R.; Neale, D.; McKeand, S.; Isik, F. Genomic estimated breeding values using genomic relation-ship matrices in a cloned population of loblolly pine. G3 Genes Genomes Genet. 2013, 3, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Lenz, P.R.N.; Beaulieu, J.; Mansfield, S.D.; Clement, S.; Desponts, M.; Bousquet, J. Factors affecting the accuracy of genomic selec-tion for growth and wood quality traits in an advanced-breeding population of black spruce (Picea mariana). BMC Genom. 2017, 18, 335. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Q.; Baison, J.; Pan, J.; Karlsson, B.; Andersson, B.; Westin, J.; Garcia-Gil, M.R.; Wu, H.X. Accuracy of genomic selection for growth and wood quality traits in two control-pollinated progeny trials using exome capture as the genotyping platform in Norway spruce. BMC Genom. 2018, 19, 946. [Google Scholar] [CrossRef] [Green Version]
- Lenz, P.R.N.; Nadeau, S.; Mottet, M.J.; Perron, M.; Isabel, N.; Beaulieu, J.; Bousquet, J. Multi-trait genomic selection for weevil re-sistance, growth, and wood quality in Norway spruce. Evol. Appl. 2020, 13, 76–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenz, P.R.N.; Nadeau, S.; Azaiez, A.; Gerardi, S.; Deslauriers, M.; Perron, M.; Isabel, N.; Beaulieu, J.; Bousquet, J. Genomic predic-tion for hastening and improving efficiency of forward selection in conifer polycross mating designs: An example from white spruce. Heredity 2020, 124, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thistlethwaite, F.; Gamal El-Dien, O.; Ratcliffe, B.; Klápště, J.; Porth, I.; Chen, C.; Stoehr, M.; Ingvarsson, P.; El-Kassaby, Y. Link-age disequilibrium vs. pedigree: Genomic selection prediction accuracy in conifer species. PLoS ONE 2020, 15, e0232201. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Hayes, B.; Daetwyler, H.D. Genomic selection in crops, trees and forages: A review. Crop Pasture Sci. 2014, 65, 1177–1191. [Google Scholar] [CrossRef]
- Bouvet, J.-M.; Makouanzi Ekomono, C.G.; Brendel, O.; Laclau, J.-P.; Bouillet, J.-P.; Epron, D. Selecting for water use efficiency, wood chemical traits and biomass with genomic selection in a Eucalyptus breeding program. Forest Ecol. Manag. 2020, 465, 118092. [Google Scholar] [CrossRef]
- Ukkola, A.M.; De Kauwe, M.G.; Roderick, M.L.; Abramowitz, G.; Pitman, A.J. Robust future changes in meteorological drought in CMIP6 projections despite uncertainty in precipitation. Geophys. Res. Lett. 2020, 47, e2020GL087820. [Google Scholar] [CrossRef]
- Fernández de Simón, B.; Sanz, M.; Sánchez-Gómez, D.; Cadahía, E.; Aranda, I. Rising [CO(2)] effect on leaf drought-induced metabolome in Pinus pinaster Aiton: Ontogenetic- and genotypic-specific response exhibit different metabolic strategies. Plant Physiol. Biochem. 2020, 149, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, A.J.; Kilgore, J.S. Soil type affects Pinus ponderosa var. scopulorum (Pinaceae) seedling growth in simulated drought experiments. Appl. Plant Sci. 2013, 1, 1300031. [Google Scholar] [CrossRef]
- Serra-Maluquer, X.; Gazol, A.; Igual, J.M.; Camarero, J.J. Silver fir growth responses to drought depend on interactions between tree characteristics, soil and neighbourhood features. Forest Ecol. Manag. 2021, 480, 118625. [Google Scholar] [CrossRef]
- Allsup, C.; Lankau, R. Migration of soil microbes may promote tree seedling tolerance to drying conditions. Ecology 2019, 100, e02729. [Google Scholar] [CrossRef] [PubMed]
- Marias Ulrich, D.; Sevanto, S.; Peterson, S.; Ryan, M.; Dunbar, J. Effects of soil microbes on functional traits of loblolly pine (Pinus taeda) seedling families from contrasting climates. Front. Plant Sci. 2020, 10, 1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Haney, C.H. Drought dampens microbiome development. Nat. Plants 2021, 7, 994–995. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the main strategies adopted by conifers to cope with drought stress.
Figure 1.
Schematic representation of the main strategies adopted by conifers to cope with drought stress.

Figure 2.
Next-generation breeding processes to improve drought stress in conifers.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Baldi, P.; La Porta, N. Toward the Genetic Improvement of Drought Tolerance in Conifers: An Integrated Approach. Forests 2022, 13, 2016. https://doi.org/10.3390/f13122016
AMA Style
Baldi P, La Porta N. Toward the Genetic Improvement of Drought Tolerance in Conifers: An Integrated Approach. Forests. 2022; 13(12):2016. https://doi.org/10.3390/f13122016
Chicago/Turabian StyleBaldi, Paolo, and Nicola La Porta. 2022. "Toward the Genetic Improvement of Drought Tolerance in Conifers: An Integrated Approach" Forests 13, no. 12: 2016. https://doi.org/10.3390/f13122016
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.