
Raffaelea quercina sp. nov. Associated with Cork Oak (Quercus suber L.) Decline in Portugal



Abstract
:1. Introduction
2. Materials and Methods
2.1. Beetle Collection and Fungal Isolation
2.2. Fungal Morphology
2.3. DNA Extraction, Amplification and Sequencing
2.4. Phylogenetic Analysis
3. Results
3.1. Fungal Isolation and Identification
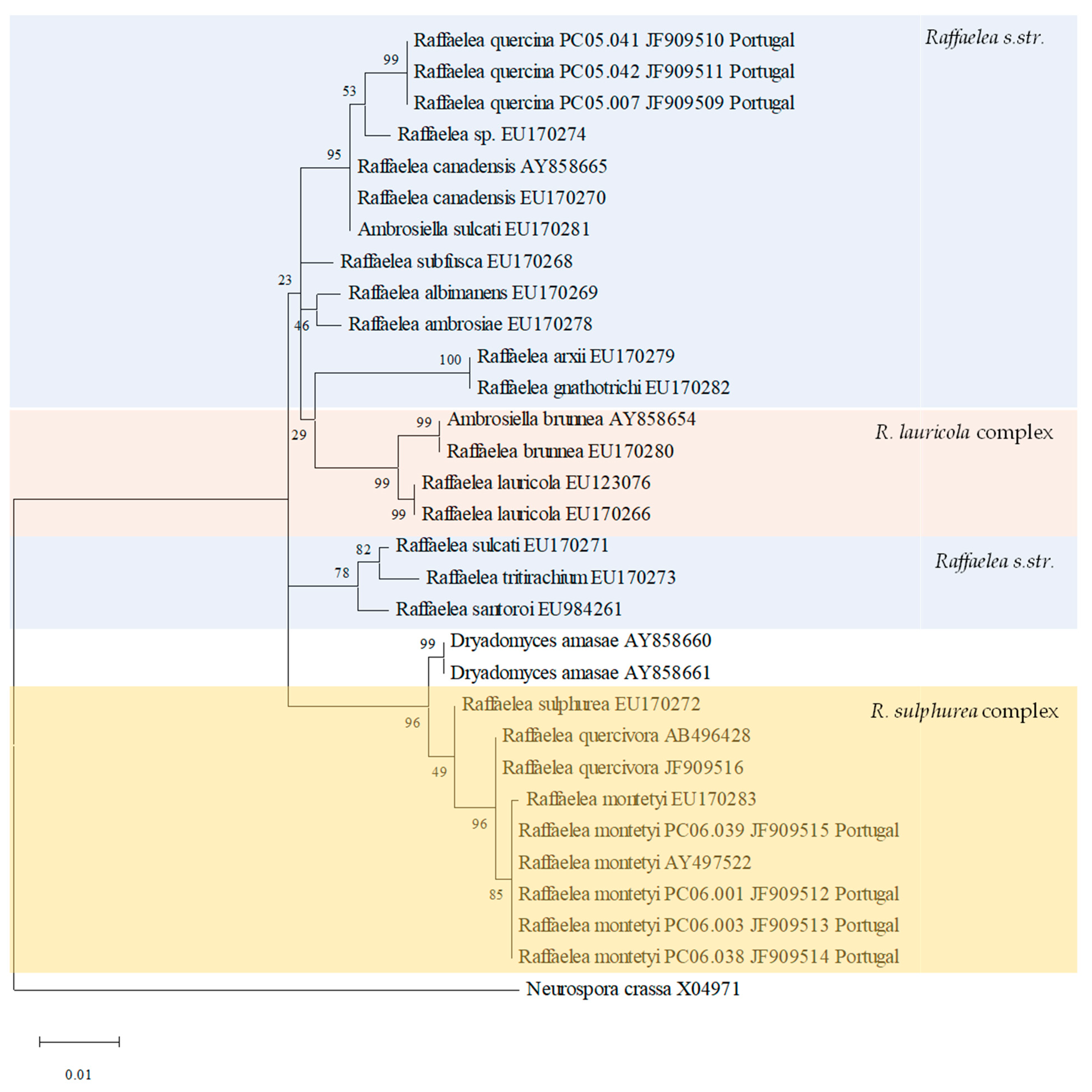
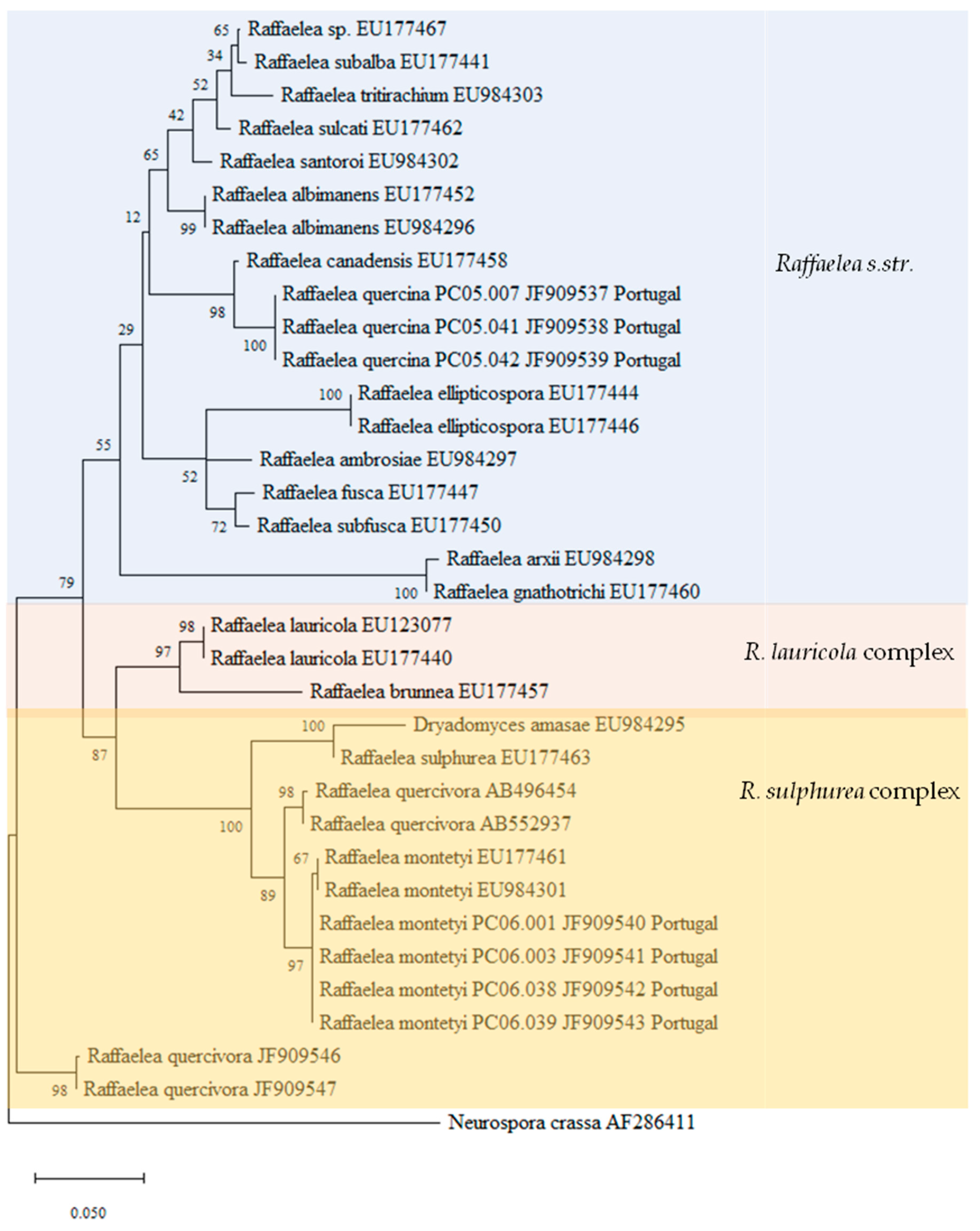
3.2. Phylogenetic Analyses
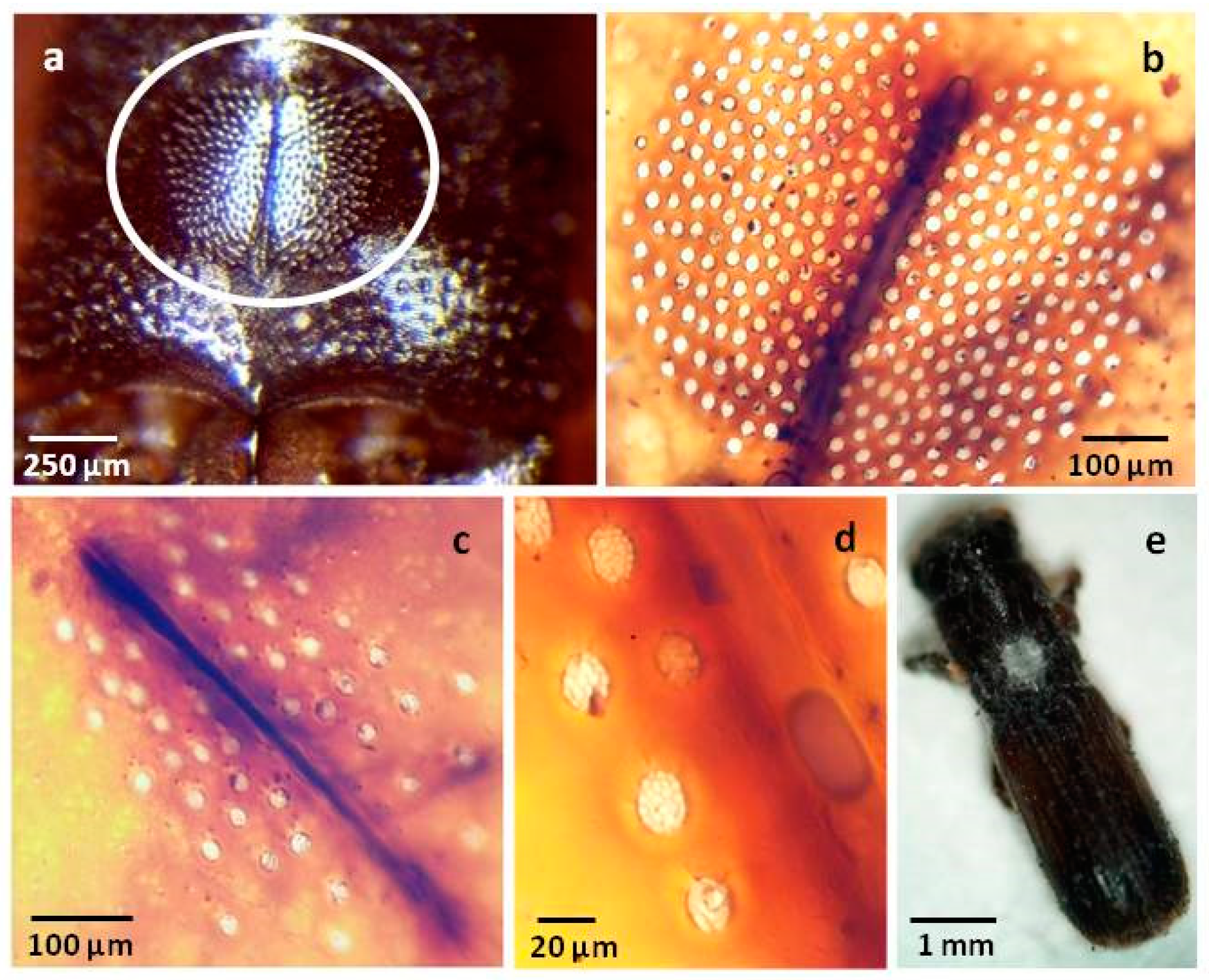
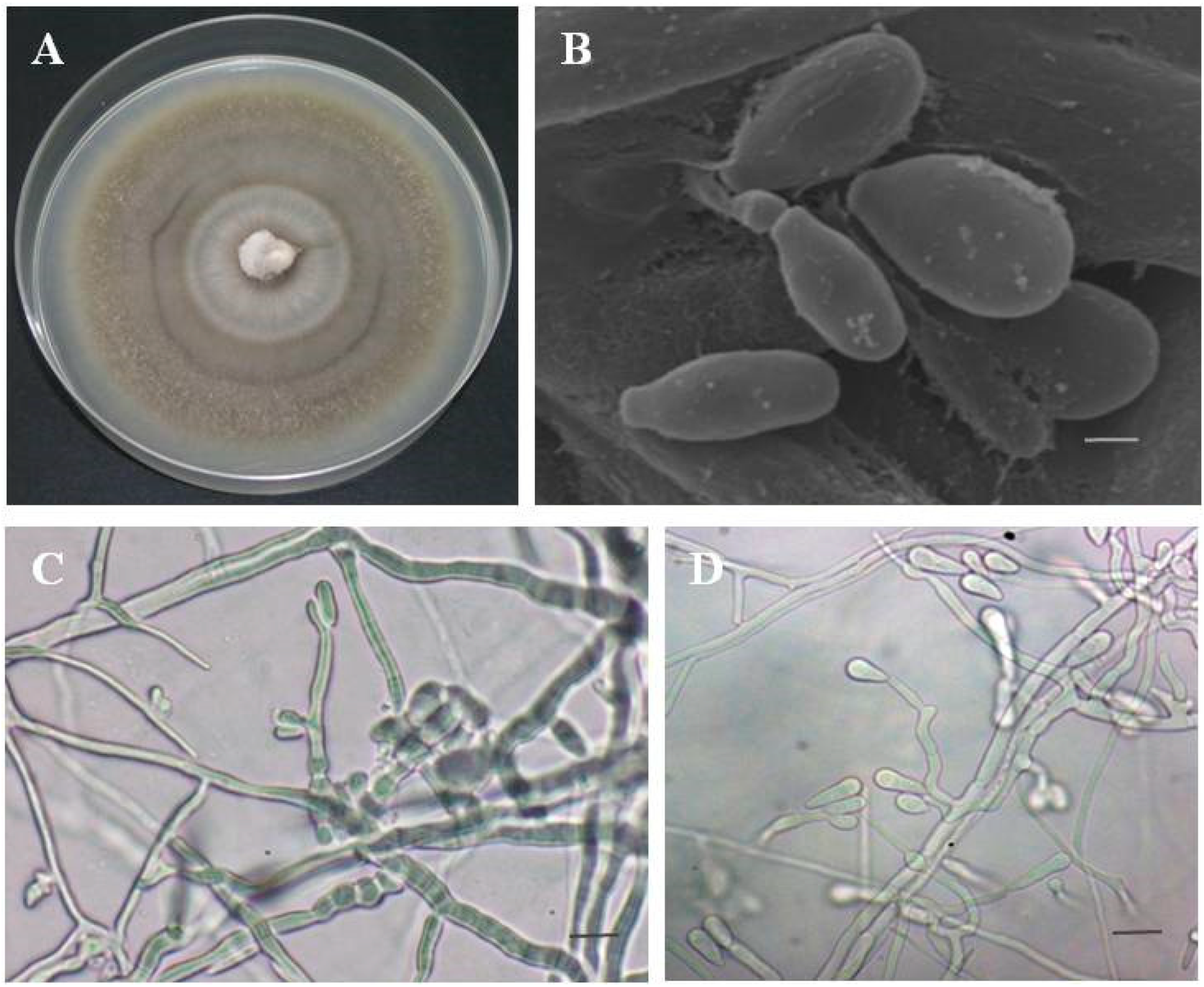
3.3. Morphology and Taxonomy
- (1)
- MycoBank: 838901
- (2)
- Holotype: LISE 96317
- (3)
- Etymology: Named after the host genus from which it was isolated, Quercus
- (4)
- Host/Distribution: On galleries of Quercus suber in Portugal = on mycangia of Playpus cylindrus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICNF. 6° Inventário Florestal Nacional (IFN6), 22 pp, Versão 1.0 2019. Instituto da Conservação da Natureza e das Florestas. Available online: http://www2.icnf.pt/portal/florestas/ifn/ifn6 (accessed on 6 December 2020).
- Cabral, M.T.; Ferreira, M.C.; Moreira, T.; Carvalho, E.C.; Diniz, A.C. Diagnóstico das causas da anormal mortalidade dos sobreiros a Sul do Tejo. Sci. Gerund. 1992, 18, 205–214. [Google Scholar]
- Cabral, M.T.; Lopes, F.; Sardinha, R.M. Determinação das causas da morte do sobreiro nos concelhos de Santiago do Cacém, Grândola e Sines. Silva Lusit. 1993, 1, 7–24. [Google Scholar]
- Inácio, M.L.; Henriques, J.; Lima, A.; Sousa, E.B. Ophiostomatoid fungi associated with cork oak mortality in Portugal. Iobc/Wprs Bull. 2012, 76, 89–92. [Google Scholar]
- Linaldeddu, B.T.; Sirca, C.; Spano, D.; Franceschini, A. Variation of endophytic cork oak-associated fungal communities in relation to plant health and water stress. For. Pathol. 2011, 41, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.N.; Jin, H.Y.; Kwak, M.J.; Khaine, I.; You, H.N.; Lee, T.Y.; Ahn, T.H.; Woo, S.Y. Why does Quercus suber species decline in Mediterranean areas? J. Asia Pac. Biodivers. 2017, 10, 337–341. [Google Scholar] [CrossRef]
- Ragazzi, A.; Vagniluca, S.; Moricca, S. European expansion of oak decline: Involved microorganisms and methodological approaches. Phytopathol. Mediterr. 1995, 34, 207–226. [Google Scholar]
- Ferreira, M.C.; Ferreira, G.W. Platypus cylindrus F. (Coleóptera: Platipodidae) Plaga de Quercus suber. Boletin. Sanid. Veg. Plagas 1989, 4, 301–305. [Google Scholar]
- Sousa, E.M.; Débouzie, D.; Pereira, H. Le rôle de l’insecte Platypus cylindrus F. (Coleoptera, Platypodidae) dans le processus de dépérissement des peuplements de chêne-liège au Portugal. IOBC WPRS Bull. 1995, 18, 24–37. [Google Scholar]
- Cabral, M.T.; Ferreira, M.C. Pragas dos Montados; Estação Florestal Nacional: Lisboa, Portugal, 1999; p. 94. [Google Scholar]
- Félix, C.; Pinto, G.; Amaral, J.; Fernandes, I.; Alves, A.; Esteves, A.C. Strain-related pathogenicity in Diplodia corticola. For. Pathol. 2017, 47, e12366. [Google Scholar] [CrossRef]
- Sousa, E.; Tomaz, I.L.; Moniz, F.A.; Basto, S. La répartition spatiale des champignons associés à Platypus cylindrus Fab. (Coleoptera: Platypodidae). Phytopathol. Mediterr. 1997, 36, 145–153. [Google Scholar]
- Santos, M.N.; Machado, M.H.; Bragança, M.H.; Ramos, H.; Sousa, E.; Tomaz, I. Mycoflora associated with cork oak (Quercus suber L.) in Portugal. IOBC WPRS Bull. 1999, 22, 25–28. [Google Scholar]
- Luque, J.; Parladé, J.; Pera, J. Pathogenicity of fungi isolated from Quercus suber in Catalonia NE Spain). For. Pathol. 2000, 30, 247–263. [Google Scholar] [CrossRef]
- Moreira, C. Distribution of Phytophthora cinnamomi in cork oak stands in Portugal. IOBC WPRS Bull. 2002, 25, 41–48. [Google Scholar]
- Franceschini, A.; Linaldeddu, B.T.; Marras, F. Occurrence and distribution of fungal endophytes in declining cork oak forests in Sardinia (Italy). IOBC WPRS Bull. 2005, 28, 67–74. [Google Scholar]
- Luque, J.; Pera, J.; Parladé, J. Evaluation of fungicides for the control of Botryosphaeria corticola on cork oak in Catalania (NE Spain). For. Pathol. 2008, 38, 147–155. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Sirca, C.; Spano, D.; Franceschini, A. Physiological responses of cork oak and holm oak to infection by fungal pathogens involved in oak decline. For. Pathol. 2009, 39, 232–238. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Scanu, B.; Maddau, L.; Franceschini, A.; Belbahri, L. Diplodia corticola and Phytophthora cinnamomi: The main pathogens involved in oak decline on Caprera Island (Italy). For. Pathol. 2014, 44, 191–200. [Google Scholar] [CrossRef]
- Harrington, T.C.; Aghayeva, D.N.; Fraedrich, S.W. New combinations in Raffaelea, Ambrosiella, and Hyalorhinocladiella, and four new species from the redbay ambrosia beetle, Xyleborus glabratus. Mycotaxon 2010, 111, 337–361. [Google Scholar] [CrossRef] [Green Version]
- De Beer, Z.W.; Wingfield, B.D.; Wingfield, M. The Ophiostoma piceae complex in the Southern Hemisphere: A phylogenetic study. Mycol. Res. 2003, 107, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Musvuugwa, T.; de Beer, Z.W.; Duong, T.A.; Dreyer, L.L.; Oberlander, K.C.; Roets, F. New species of Ophiostomatales from Scolytinae and Platypodinae beetles in the Cape Floristic Region, including the discovery of the sexual state of Raffaelea. Antonie Van Leeuwenhoek J. Microbiol. 2015, 108, 933–950. [Google Scholar] [CrossRef] [PubMed]
- Kubono, T.; Ito, S. Raffaelea quercivora sp. nov. associated with mass mortality of Japanese oak, and the ambrosia beetle (Platypus quercivorus). Mycoscience 2002, 43, 255–260. [Google Scholar] [CrossRef]
- Kim, K.H.; Choi, Y.J.; Seo, S.T.; Shin, H.D. Raffaelea quercus-mongolicae sp. nov. associated with Platypus koryoensis on oak in Korea. Mycotaxon 2009, 110, 189–197. [Google Scholar] [CrossRef]
- Harrington, T.C.; Fraedrich, S.W.; Aghayeva, D.N. Raffaelea lauricola, a new ambrosia beetle symbiont and pathogen on the Lauraceae. Mycotaxon 2008, 104, 399–404. [Google Scholar]
- Gebhardt, H.; Begerow, D.; Oberwinkler, F. Identification of the ambrosia fungus of Xyleborus monographus and X. dryographus (Coleoptera: Curculionidae, Scolytinae). Mycol. Prog. 2004, 3, 95–102. [Google Scholar] [CrossRef]
- Matsuda, Y.; Kimura, K.; Ito, S. Genetic characterization of Raffaelea quercivora isolates collected from areas of oak wilt in Japan. Mycoscience 2010, 51, 310–316. [Google Scholar] [CrossRef]
- Saccardo, P.A. Chromotaxia seu Nomenclator Colorum Polyglottus Adclitis Speciminibus Coloratis ad Botanicorum et Zoologorum; Patavii, Italy, 1891; p. 22. [Google Scholar]
- Riddell, R.W. Permanent stained mycological preparations obtained by slide culture. Mycologia 1950, 42, 265–270. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.-J.; Fung, S.; Breuil, C. A PCR-RFLP marker distinguishing Ophiostoma clavigerum from morphologically similar Leptographium species associated with bark beetles. Can. J. Bot. 2003, 81, 1104–1112. [Google Scholar] [CrossRef]
- Massoumi Alamouti, S.; Tsui, C.M.; Breuil, C. Multigene phylogeny of filamentous ambrosia fungi associated with ambrosia and bark beetles. Mycol. Res. 2009, 113, 822–835. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acid Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clamp, M.; Cuff, J.; Searle, S.M.; Barton, G.J. The Jalview java alignment editor. Bioinformatics 2004, 20, 426–427. [Google Scholar] [CrossRef] [Green Version]
- Gebhardt, H.; Oberwinkler, F. Conidial development in selected ambrosial species of the genus Raffaelea. Antonie Van Leeuwenhoek 2005, 88, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.R.; Zhang, Y.Z.; Qiao, H.J.; Shi, W.F.; Murphy, R.W.; Zhu, C.-D. Outgroup selection in tree reconstruction: A case of the family Halictidae (Hymenoptera: Apoidea). Acta Entomol. Sin. 2010, 53, 192–201. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Morelet, M. Une espèce nouvelle de Raffaelea, isolée de Platypus cylindrus, coléoptère xylomycetophage des chênes. Ann. De La Société Des. Sci. Nat. De Toulon Et Du Var. 1998, 50, 185–193. [Google Scholar]
- Sousa, E.M. Contribution à l’étude de la biologie de populations de Platypus cylindrus (Coleóptera: Platypodidae) dans des peuplements de chênes liège au Portugal. In Thèse de Doctorat; Université de Lyon: Lyon, France, 1996; p. 153. [Google Scholar]
- Baker, J.M. Ambrosia beetle and their fungi, with particular reference to Platypus cylindrus Fab. Symp. Soc. Gen. Microbiol. 1963, 13, 323–354. [Google Scholar]
- Von Arx, J.A.; Hennebert, G.L. Deux champignons ambrosia. Mycopathol. Et. Mycol. Appl. 1965, 25, 309–315. [Google Scholar] [CrossRef]
- Funk, A. Fungal symbionts of the ambrosia beetle Gnathotrichus sulcatus. Can. J. Bot. 1970, 48, 1445–1448. [Google Scholar] [CrossRef]
- Harrington, T.C.; Yun, H.Y.; Lu, S.; Goto, H.; Aghayeva, D.; Fraedrich, S. Isolations from the redbay ambrosia beetle, Xyleborus glabratus, confirm that the laurel wilt pathogen, Raffaelea lauricola, originated in Asia. Mycologia 2011, 103, 1028–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batra, L.R. Ecology of ambrosia fungi and their dissemination by beetles. Trans. Kans. Acad. Sci. 1963, 66, 213–236. [Google Scholar] [CrossRef]
- Simmons, D.R.; de Beer, Z.W.; Huang, Y.-T.; Bateman, C.; Campbell, A.S.; Dreaden, T.J.; Li, Y.; Ploetz, R.C.; Black, A.; Li, H.-F.; et al. New Raffaelea species (Ophiostomatales) from the USA and Taiwan associated with ambrosia beetles and plant hosts. IMA Fungus 2016, 7, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Vanderpool, D.; Bracewell, R.R.; McCutcheon, J.P. Know your farmer: Ancient origins and multiple independent domestications of ambrosia beetle fungal cultivars. Mol. Ecol. 2017, 27, 2077–2094. [Google Scholar] [CrossRef] [PubMed]
- Mayers, C.G.; Harrington, T.C.; Mcnewa, D.L.; Roeperb, R.A.; Biedermann, P.; Masuyad, H.; Bateman, C.C. Four mycangium types and four genera of ambrosia fungi suggest a complex history of fungus farming in the ambrosia beetle tribe Xyloterini. Mycologia 2020, 112, 1104–1137. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, Y.T.; Kasson, M.T.; Macias, A.M.; Skelton, J.; Carlson, P.S.; Yin, M.; Hulcr, J. Specific and promiscuous ophiostomatalean fungi associated with Platypodinae ambrosia beetles in the southeastern United States. Fungal Ecol. 2018, 35, 42–50. [Google Scholar] [CrossRef]
- Fraedrich, S.W.; Harrington, T.C.; Rabaglia, R.J.; Ulyshen, M.D.; Mayfield, A.E.; Hanula, J.L.; Eickwort, J.M.; Miller, D.R. A fungal symbiont of the redbay ambrosia beetle causes a lethal wilt in redbay and other Lauraceae in the southeastern United States. Plant Dis. 2008, 92, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskalen, A.; McDonald, V. First Report of Raffaelea canadensis Causing Laurel Wilt Disease Symptoms on Avocado in California. Plant Dis. 2011, 95, 1189. [Google Scholar] [CrossRef] [PubMed]
- Procter, M.; Nel, W.J.; Marincowitz, S.; Crous, P.W.; Wingfield, M.J. A new species of Raffaelea from beetle-infested Leucaena leucocephala. Fungal Syst. Evol. 2020, 6, 305–314. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection No. a | Country | Host/Source | GenBank Accession Nrs. b | |
|---|---|---|---|---|---|
| SSU sDNA | LSU rDNA | ||||
| R. quercina quercina sp. nov. | MEAN 1290 = PC05.007 = C2512 = CBS 147554 | Portugal (Santarém) | Platypus cylindrus | JF909509 | JF909537 |
| R. quercina quercina sp. nov. | MEAN 1291 = PC05.041 = C2516 | Portugal (Santarém) | P. cylindrus | JF909510 | JF909538 |
| R. quercina quercina sp. nov. | MEAN 1292 = PC05.042 = C2513 = CBS 147555 | Portugal (Santarém) | Quercus suber, P. cylindrus | JF909511 | JF909539 |
| R. albimanens | C2223 = CBS 271.70 | S. Africa | P. externedentatus | EU170269 | |
| R. albimanens | CBS271.70 | S. Africa | EU984296 | ||
| R. ambrosiae | C2225 = CBS 185.64 | UK | P. cylindrus | EU170278 | EU984297 |
| R. amasae | TUB F4290 | Germany | Amasa aff. glaber | AY858661 | |
| R. arxii | C2218 = CBS 273.70 | South Africa | Xyleborus torquatus | EU170279 | EU984298 |
| R. brunnea | C2229 = CBS378.68 | USA | Monarthrum sp. | EU170280 | EU177457 |
| R. canadensis | C2233 = CBS 168.66 | Canada | P. wilsonni | EU170270 | EU177458 |
| R. canadensis | CBS 168.66 | Canada | P. wilsonni | AY858665 | |
| A. sulcati | C592 = CBS 805.70 | Canada | Gnathotrichus sulcatus | EU170281 | |
| R. ellipticospora | C2346 | USA | X. glabratus | EU177444 | |
| R. fusca | C2254 | USA | X. glabratus | EU177447 | |
| R. gnathotrichi | C2219 = CBS 379.68 | USA | G. retusus | EU170282 | EU177460 |
| R. lauricola | C2203 | USA | X. glabratus | EU123076 | EU123077 |
| R. lauricola | C2339 = CBS 121567 | USA | X. glabratus | EU177440 | |
| R. montetyi | PC06.001, C2506 | Portugal (Alentejo) | P. cylindrus | JF909512 | JF909540 |
| R. montetyi | PC07.003, C2505 | Portugal (Alentejo) | Quercus suber, P. cylindrus | JF909513 | JF909541 |
| R. montetyi | PC06.038, C2504 | Portugal (Alentejo) | P. cylindrus | JF909514 | JF909542 |
| R. montetyi | PC06.039, C2514 | Portugal (Alentejo) | Quercus suber, P. cylindrus | JF909515 | JF909543 |
| Species | Collection No. a | Country | Host/Source | GenBank accession nrs. b | |
| SSU sDNA | LSU rDNA | ||||
| C2220 = CBS451.94 | Portugal (Alentejo) | P. cylindrus | - | EU177461 | |
| CBS451.94 | Portugal | P. cylindrus | EU984301 | ||
| C2221 = CBS463.94 | France | P. cylindrus | EU170283 | ||
| TUB F4264 | Germany | X. dryographus | AY497522 | ||
| R. quercivora | PC10.919, MAFF919 | Japan | P. quercivorus | JF909517 | JF909547 |
| PC10.921, MAFF921 | Japan | P. quercivorus | JF909516 | JF909546 | |
| JCM 15683 | Japan | P. quercivorus | AB552937 | ||
| MAFF410918 | Japan | Q. crispula/P. quercivorus | AB496428 | AB496454 | |
| R. santoroi | CBS 399.67 | Argentina | M. mutatus | EU984261 | EU984302 |
| R. subalba | C2368 | USA | X. glabratus | EU177441 | |
| R. subfusca | C2253 | USA | X. glabratus | EU170268 | |
| R. sulcati | C2234 = CBS806.70 | Canada | G. sulcatus | EU170271 | EU177462 |
| R. sulphurea | C593 = CBS380.68 | USA | X. saxeseni | EU170272 | EU177463 |
| R. tritirachium | C2222 = CBS 726.69 | USA | M. mali | EU170273 | EU984303 |
| Raffaelea sp. | C1941 | USA | unknown | EU170274 | |
| Raffaelea sp. | C2224 = CBS326.70 | S. Africa | P. externedentatus | EU177467 | |
| R. amasae | CBS 116694 | Taiwan | A. concitatus | AY858660 | |
| R. brunnea | CBS378.68 | unknown | Monarthrum sp. | AY858654 | |
| R. lauricola | C2204 | USA | X. glabratus | EU170266 | |
| R. subfusca | C2335 = CBS121571 | USA | X. glabratus | EU177450 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inácio, M.L.; Marcelino, J.; Lima, A.; Sousa, E.; Nóbrega, F. Raffaelea quercina sp. nov. Associated with Cork Oak (Quercus suber L.) Decline in Portugal. Forests 2021, 12, 513. https://doi.org/10.3390/f12040513
Inácio ML, Marcelino J, Lima A, Sousa E, Nóbrega F. Raffaelea quercina sp. nov. Associated with Cork Oak (Quercus suber L.) Decline in Portugal. Forests. 2021; 12(4):513. https://doi.org/10.3390/f12040513
Chicago/Turabian StyleInácio, Maria L., José Marcelino, Arlindo Lima, Edmundo Sousa, and Filomena Nóbrega. 2021. "Raffaelea quercina sp. nov. Associated with Cork Oak (Quercus suber L.) Decline in Portugal" Forests 12, no. 4: 513. https://doi.org/10.3390/f12040513