
Resistance, Recovery and Resilience of Two Co-Occurring Palaeotropical Pinus Species Differing in the Sizes of Their Distribution Areas


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Tree-Ring Analyses
2.3. Cellulose Extraction and Stable Carbon Isotope Analysis
2.4. Climate Data
2.5. Stress Events and Sensitivity Indicators
2.6. Soil Analyses
2.7. Statistical Analyses
3. Results
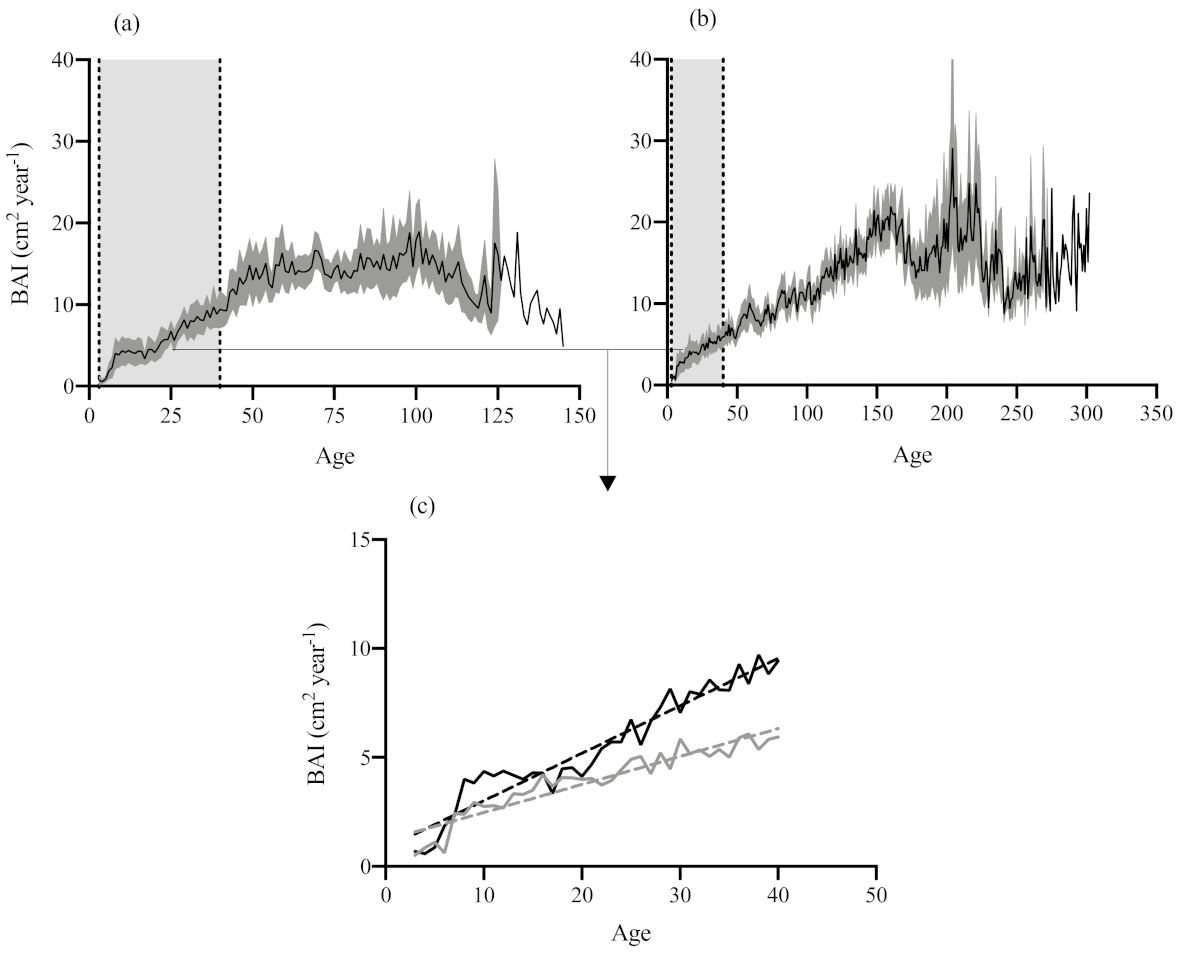
3.1. Stand Structure, Soil Conditions and Tree Growth
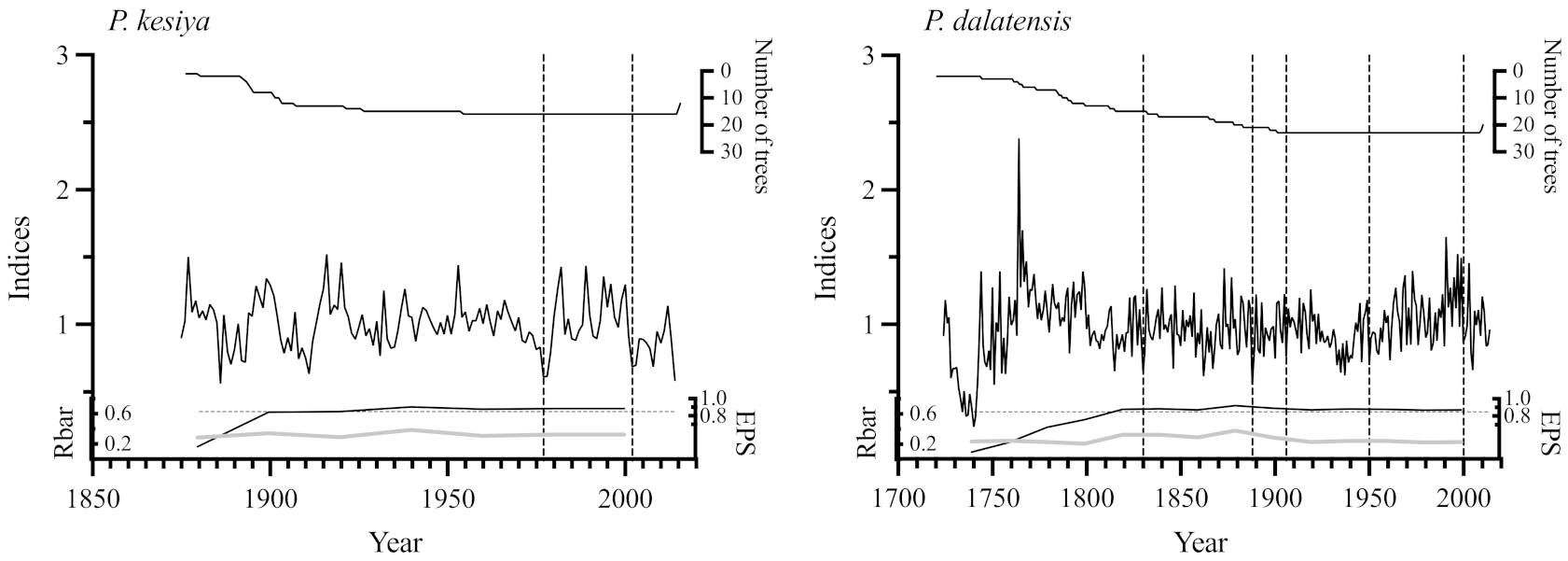
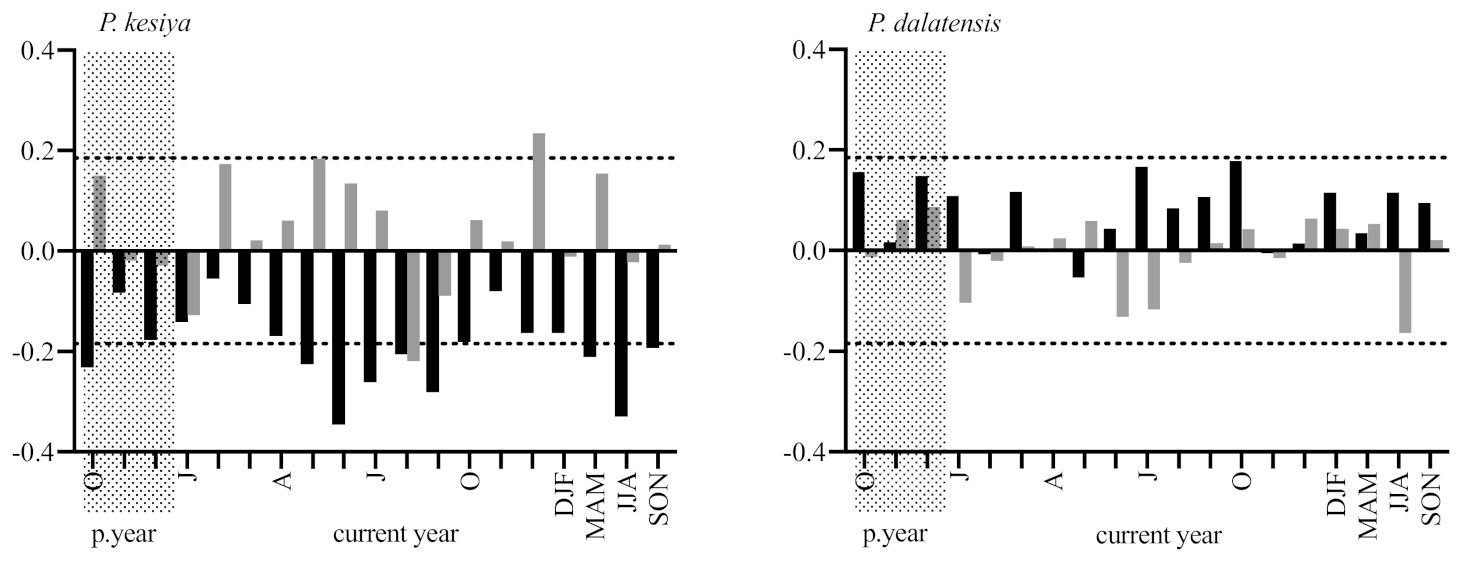
3.2. Tree-Ring Indices and Climate Signals
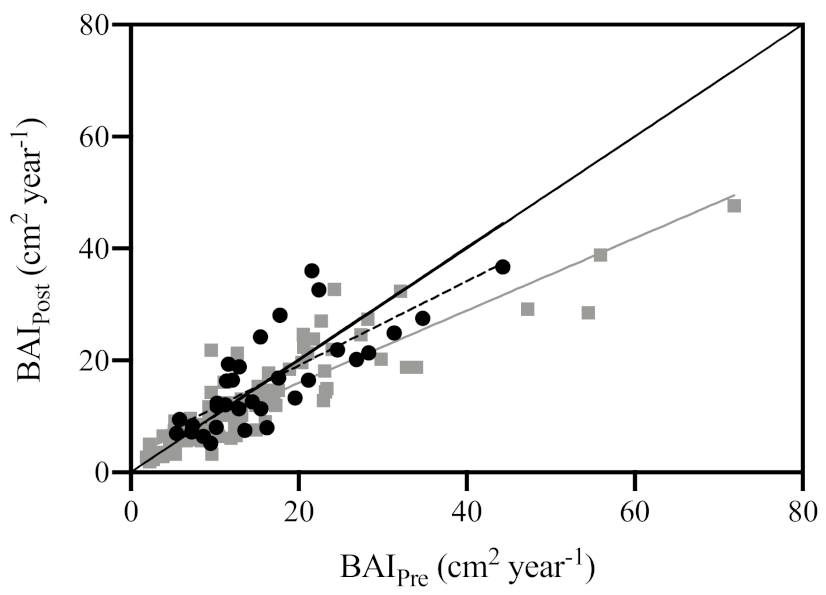
3.3. Sensitivity Indicators
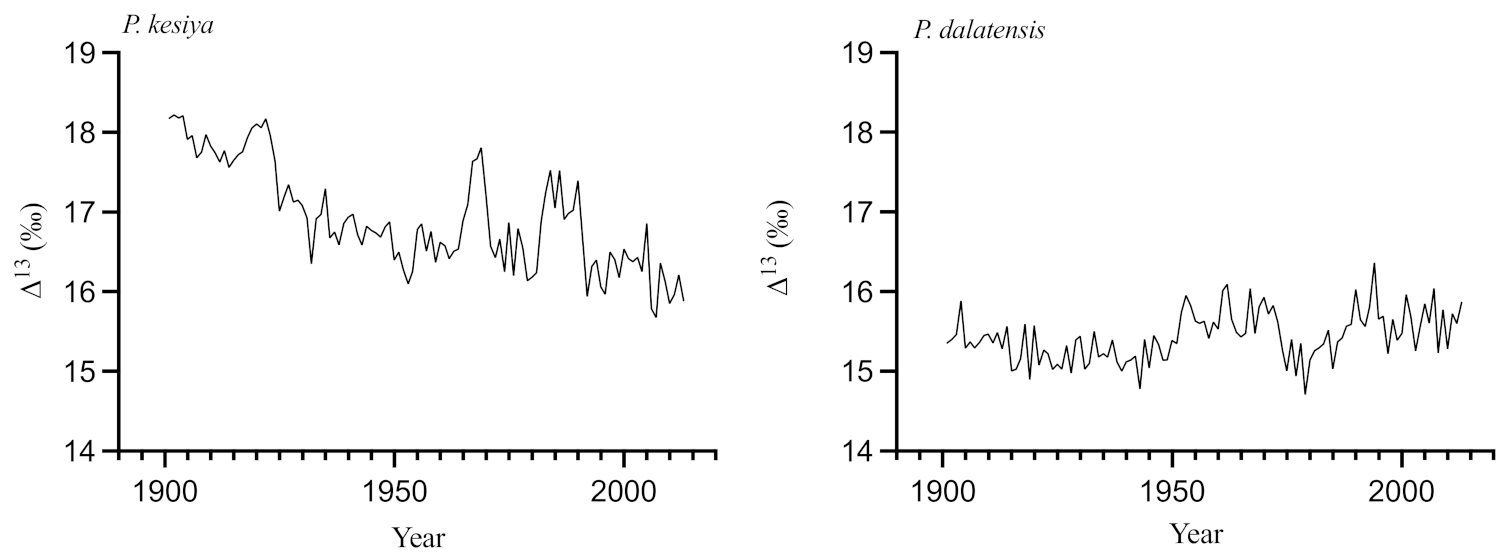
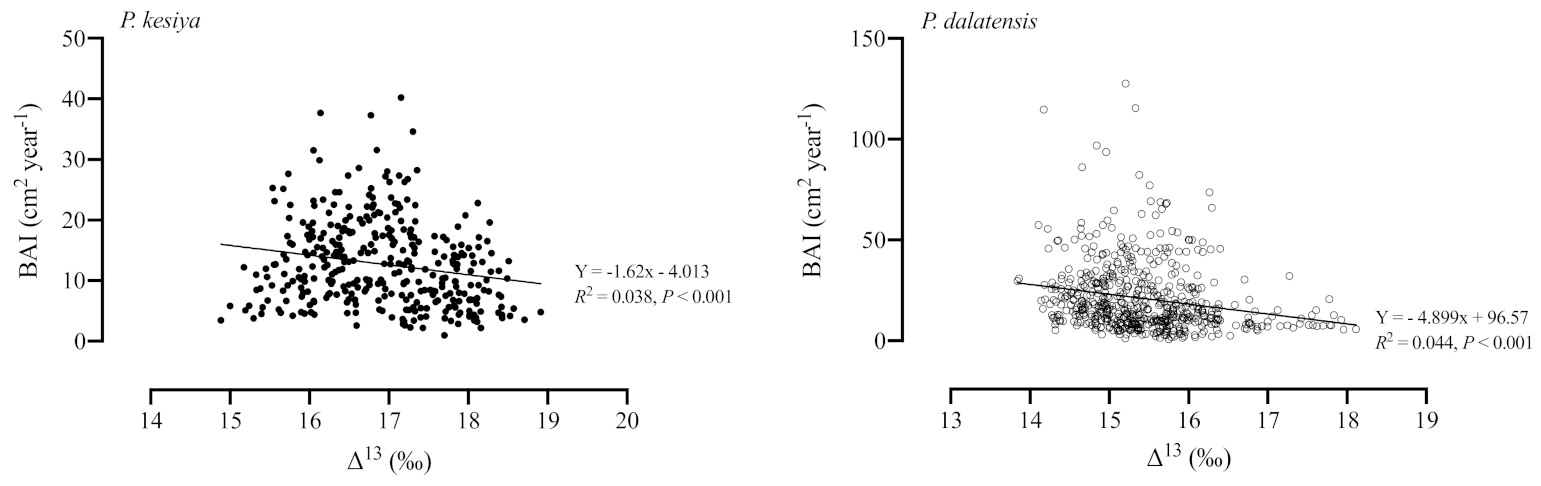
3.4. Stable Carbon Isotope Signals
4. Discussion
4.1. Tree Growth and Climate Signals
4.2. Growth Sensitivity Indicators
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richardson, D.; Rundel, P. Ecology and Biogeography of Pinus: An Introduction pp 3–49 in Richardson DM (ed) Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Luu, N.D.T.; Thomas, P. Conifers of Vietnam: An Illustrated Field Guide for the Most Important Forest Trees; Darwin Initiative: Hanoi, Vietnam, 2004; p. 86. [Google Scholar]
- Schorcht, M.; Roloff, A.; Weisgerber, H.; Lang, U. Pinus Kesiya Royle ex Gordon; Wiley VCH: Weinheim, Germany, 1997; pp. 1–8. [Google Scholar]
- Sterling, E.; Hurley, M.; Minh, L. A Natural History of Vietnam; Yale University Express: New Haven, CT, USA, 2006. [Google Scholar]
- Silba, J. Encyclopaedia Coniferae; Moldenke, H.N., Moldenke, A.L., Eds.; New York Botanical Garden: Corvallis, OR, USA, 1986. [Google Scholar]
- Thomas, P.; Sengdala, K.; Lamxay, V.; Khou, E. New Records of Conifers in Cambodia and Laos. Edinb. J. Bot. 2007, 64, 37–44. [Google Scholar] [CrossRef]
- Rácz, I.; Huyen, D. Study of a low-elevation occurrence of Pinus dalatensis Ferré (Pinaceae) in Gia Lai province, Vietnam. Studia Bot Hung 2007, 38, 133–142. [Google Scholar]
- Businsky, R. Study of Pinus dalatensis Ferre and of the enigmatic “Pin du Moyen Annam”. Candollea 1999, 54, 125–144. [Google Scholar]
- Thomas, P.; Phan, K.L. Pinus dalatensis. The IUCN Red List of Threatened Species. 2013. Available online: https://www.iucnredlist.org/species/32803/2823679 (accessed on 1 February 2021).
- Hiep, N.T.; Loc, P.K.; Luu, N.D.T.; Thomas, P.; Farjon, A.; Averyanov, L.V.; Regalado, J., Jr. Vietnam Conifers: Conservation Status Review 2004; Fauna & Flora International, Vietnam Programme: Hanoi, Vietnam, 2004. [Google Scholar]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Gazol, A.; Camarero, J.J.; Anderegg, W.R.L.; Vicente-Serrano, S.M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Glob. Ecol. Biogeogr. 2017, 26, 166–176. [Google Scholar] [CrossRef]
- Vitali, V.; Buntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, N.; Schall, P.; Ammer, C.; Leder, B.; Vor, T. Drought sensitivity and stem growth variation of nine alien and native tree species on a productive forest site in Germany. Agric. Forest Meteorol. 2018, 256, 431–444. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schutze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. (Stuttg.) 2013, 15, 483–495. [Google Scholar] [CrossRef]
- Rais, A.; van de Kuilen, J.-W.G.; Pretzsch, H. Growth reaction patterns of tree height, diameter, and volume of Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco) under acute drought stress in Southern Germany. Eur. J. Forest Res. 2014, 133, 1043–1056. [Google Scholar] [CrossRef]
- Cavin, L.; Jump, A.S. Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial range edge. Glob. Chang. Biol. 2017, 23, 362–379. [Google Scholar] [CrossRef] [Green Version]
- Gazol, A.; Camarero, J.J.; Sangüesa-Barreda, G.; Vicente-Serrano, S.M. Post-drought resilience after forest die-off: Shifts in regeneration, composition, growth and productivity. Front. Plant Sci. 2018, 9, 1546. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Vicente-Serrano, S.M.; Sánchez-Salguero, R.; Gutiérrez, E.; de Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; Tíscar, P.A. Forest resilience to drought varies across biomes. Glob. Chang. Biol. 2018, 24, 2143–2158. [Google Scholar] [CrossRef] [PubMed]
- Marqués, L.; Camarero, J.J.; Gazol, A.; Zavala, M.A. Drought impacts on tree growth of two pine species along an altitudinal gradient and their use as early-warning signals of potential shifts in tree species distributions. Forest Ecol. Manag. 2016, 381, 157–167. [Google Scholar] [CrossRef]
- Rubio-Cuadrado, Á.; Camarero, J.J.; Aspizua, R.; Sánchez-González, M.; Gil, L.; Montes, F. Abiotic factors modulate post-drought growth resilience of Scots pine plantations and rear-edge Scots pine and oak forests. Dendrochronologia 2018, 51, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Vitasse, Y.; Bottero, A.; Cailleret, M.; Bigler, C.; Fonti, P.; Gessler, A.; Lévesque, M.; Rohner, B.; Weber, P.; Rigling, A. Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob. Chang. Biol. 2019, 25, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Anh, P.L.; Vladimirov, A.; Kruk, N.; Polyakov, G.; Ponomarchuk, V.; Hoa, T.T.; Phuong, N.T.; Kuibida, M.; Annikova, I.Y.; Pavlova, G. Stanniferrous granites of Vietnam: Rb-Sr and Ar-Ar isotope age, composition, sources, and geodynamic formation conditions. Dokl. Earth Sci. 2021, 1, 839–845. [Google Scholar] [CrossRef]
- Singh, N.D.; Venugopal, N. Cambial activity and annual rhythm of xylem production of Pinus kesiya Royle ex. Gordon (Pinaceae) in relation to phenology and climatic factors growing in sub-tropical wet forest of North East India. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 198–204. [Google Scholar] [CrossRef]
- Stokes, M.; Smiley, T. Introduction to Tree-Ring Dating; University of Chicago: Chicago, IL, USA, 1968. [Google Scholar]
- Rinn, F. TSAP-Win: Time Series Analysis and Presentation for Dendrochronology and 409 Related Applications; Frank Rinn: Heidelberg, Germany, 2003. [Google Scholar]
- Schweingruber, F.H. Der Jahrring: Standort, Methodik, Zeit und Klima in der Dendrochronologie; Paul Haupt: Bern, Switzerland; Stuttgart, Germany, 1983. [Google Scholar]
- Esper, J.; Cook, E.R.; Krusic, P.J.; Peters, K.; Schweingruber, F.H. Tests of the RCS method for preserving low-frequency variability in long tree-ring chronologies. Tree Ring Res. 2003, 59, 81–98. [Google Scholar]
- Cook, E.R. A Time Series Analysis Approach to Tree Ring Standardization; University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Wigley, T.M.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Appl. Meteorol. Climatol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Biondi, F.; Qeadan, F. A theory-driven approach to tree-ring standardization: Defining the biological trend from expected basal area increment. Tree Ring Res. 2008, 64, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Loader, N.; Robertson, I.; Barker, A.; Switsur, V.; Waterhouse, J. An improved technique for the batch processing of small wholewood samples to α-cellulose. Chem. Geol. 1997, 136, 313–317. [Google Scholar] [CrossRef]
- McCarroll, D.; Gagen, M.H.; Loader, N.J.; Robertson, I.; Anchukaitis, K.J.; Los, S.; Young, G.H.F.; Jalkanen, R.; Kirchhefer, A.; Waterhouse, J.S. Correction of tree ring stable carbon isotope chronologies for changes in the carbon dioxide content of the atmosphere. Geochim. Cosmochim. Acta 2009, 73, 1539–1547. [Google Scholar] [CrossRef]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- Saurer, M.; Siegwolf, R.T.W. Human Impacts on Tree-Ring Growth Reconstructed from Stable Isotopes. In Terrestrial Ecology; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, pp. 49–62. [Google Scholar]
- Trouet, V.; Van Oldenborgh, G.J. KNMI Climate Explorer: A Web-Based Research Tool for High-Resolution Paleoclimatology. Tree Ring Res. 2013, 69, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations–the CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Schneider, U.; Becker, A.; Finger, P.; Meyer-Christoffer, A.; Ziese, M.; Rudolf, B. GPCC’s new land surface precipitation climatology based on quality-controlled in situ data and its role in quantifying the global water cycle. Theor. Appl. Climatol. 2014, 115, 15–40. [Google Scholar] [CrossRef] [Green Version]
- Steubing, L.; Fangmeier, A. Plant Ecological Training Course: Field and Laboratory Practicals in Terrestrial Plant Ecology; Verlag Eugen: Stuttgart, Germany, 1992. [Google Scholar]
- Murphy, H.T.; VanDerWal, J.; Lovett-Doust, J. Distribution of abundance across the range in eastern North American trees. Glob. Ecol. Biogeogr. 2006, 15, 63–71. [Google Scholar] [CrossRef]
- Lomolino, M.V.; Riddle, B.R.; Whittaker, R.J. Biogeography, 5th ed. J. Bot. Res. Inst. Tex. 2018, 12, 26. [Google Scholar]
- Fréjaville, T.; Vizcaíno-Palomar, N.; Fady, B.; Kremer, A.; Benito Garzón, M. Range margin populations show high climate adaptation lags in European trees. Glob. Chang. Biol. 2020, 26, 484–495. [Google Scholar] [CrossRef]
- Lennon, J.J.; Kunin, W.E.; Corne, S.; Carver, S.; Van Hees, W.W. Are Alaskan trees found in locally more favourable sites in marginal areas? Glob. Ecol. Biogeogr. 2002, 11, 103–114. [Google Scholar] [CrossRef]
- Bigler, C.; Veblen, T.T. Increased early growth rates decrease longevities of conifers in subalpine forests. Oikos 2009, 118, 1130–1138. [Google Scholar] [CrossRef]
- Castagneri, D.; Storaunet, K.O. Age-related growth trends in ancient Norway spruce trees and potential effects on long term growth patterns. Trace Tree Rings Archaeol. Climatol. Ecol. 2011, 9, 58–63. [Google Scholar]
- Büntgen, U.; Krusic, P.J.; Piermattei, A.; Coomes, D.A.; Esper, J.; Myglan, V.S.; Kirdyanov, A.V.; Camarero, J.J.; Crivellaro, A.; Körner, C. Limited capacity of tree growth to mitigate the global greenhouse effect under predicted warming. Nat. Commun. 2019, 10, 1–6. [Google Scholar]
- Bugmann, H.; Bigler, C. Will the CO 2 fertilization effect in forests be offset by reduced tree longevity? Oecologia 2011, 165, 533–544. [Google Scholar] [CrossRef]
- Sillett, S.C.; Van Pelt, R.; Koch, G.W.; Ambrose, A.R.; Carroll, A.L.; Antoine, M.E.; Mifsud, B.M. Increasing wood production through old age in tall trees. For. Ecol. Manag. 2010, 259, 976–994. [Google Scholar] [CrossRef]
- Sheil, D.; Eastaugh, C.S.; Vlam, M.; Zuidema, P.A.; Groenendijk, P.; van der Sleen, P.; Jay, A.; Vanclay, J. Does biomass growth increase in the largest trees? Flaws, fallacies and alternative analyses. Funct. Ecol. 2017, 31, 568–581. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Eckstein, D. Reconstruction of pre-monsoon weather conditions in northwestern Thailand from the tree-ring widths of Pinus merkusii and Pinus kesiya. Trees 2011, 25, 125–132. [Google Scholar] [CrossRef]
- Singh, N.D.; Yadav, R.R.; Venugopal, N.; Singh, V.; Yadava, A.K.; Misra, K.G.; Singh, T.B.; Sanjita, C. Climate control on ring width and intra-annual density fluctuations in Pinus kesiya growing in a sub-tropical forest of Manipur, Northeast India. Trees 2016, 30, 1711–1721. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Wanyaphet, T. Seasonal cambial activity and tree-ring formation of Pinus merkusii and Pinus kesiya in Northern Thailand in dependence on climate. For. Ecol. Manag. 2006, 226, 279–289. [Google Scholar] [CrossRef]
- Ho, L.T.; Schneider, R.; Thomas, F.M. Growth of the tropical Pinus kesiya as influenced by climate and nutrient availability along an elevational gradient. J. Plant Ecol. 2020, 13, 97–106. [Google Scholar] [CrossRef]
- Graham, E.A.; Mulkey, S.S.; Kitajima, K.; Phillips, N.G.; Wright, S.J. Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons. Proc. Natl. Acad. Sci. USA 2003, 100, 572–576. [Google Scholar] [CrossRef] [Green Version]
- Sutton, W.R.; Srivastava, J.P.; Rosegrant, M.; Thurlow, J.; Sebastian, L. Striking a Balance: Managing El Niño and La Niña in Vietnam’s Agriculture; World Bank: Washington, DC, USA, 2019. [Google Scholar]
- Martinez-Vilalta, J.; Lopez, B.C.; Loepfe, L.; Lloret, F. Stand- and tree-level determinants of the drought response of Scots pine radial growth. Oecologia 2012, 168, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Yoder, B.; Ryan, M.; Waring, R.; Schoettle, A.; Kaufmann, M. Evidence of reduced photosynthetic rates in old trees. For. Sci. 1994, 40, 513–527. [Google Scholar]
- Bose, A.K.; Gessler, A.; Bolte, A.; Bottero, A.; Buras, A.; Cailleret, M.; Camarero, J.J.; Haeni, M.; Hereş, A.M.; Hevia, A. Growth and resilience responses of Scots pine to extreme droughts across Europe depend on predrought growth conditions. Glob. Chang. Biol. 2020, 26, 4521. [Google Scholar] [CrossRef]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef]
- Matesanz, S.; Gianoli, E.; Valladares, F. Global change and the evolution of phenotypic plasticity in plants. Ann. N. Y. Acad. Sci. 2010, 1206, 35–55. [Google Scholar] [CrossRef]
- Eckert, A.J.; Hall, B.D. Phylogeny, historical biogeography, and patterns of diversification for Pinus (Pinaceae): Phylogenetic tests of fossil-based hypotheses. Mol. Phylogenetics Evol. 2006, 40, 166–182. [Google Scholar] [CrossRef]
- Xu, X.-H.; Wang, Z.-X.; Yang, G.-L.; Wang, J.; Yang, Y.; Ma, F.-J.; Wang, Q.-J.; Li, R.-Y.; Sun, B.-N. Two Pinus species from the upper Miocene in Zhejiang, China and their palaeobiogeographic significance. Rev. Palaeobot. Palynol. 2015, 215, 68–75. [Google Scholar] [CrossRef]
- Hong, Y.; Song, G.; Rui-Fu, H.; Han-Qiao, J. A preliminary study on genetic variation and relationships of Pinus yunnanensis and its closely related species. J. Integr. Plant Biol. 2000, 42, 107–110. [Google Scholar]
- Potter, L. The onslaught on the forests in South-East Asia. Brookfield and Byron. In South-East Asia’s Environmental Future—The Search for Sustainability; Brookfield, H., Byron, Y., Eds.; United Nations University Press & Oxford University Press: Tokyo, Japan, 1993; pp. 103–123. [Google Scholar]
- Kuznetsov, A.; Kuznetsova, S. Tropical monsoon forests of Vietnam (results of 20 years of phytoecological research). Biol. Bull. 2013, 40, 187–196. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Elevation (m a.s.l.) | Longitude; Latitude | dbh (cm) | Tree Height (m) | Tree Density (Tree ha−1) | Number of Analysed Trees |
|---|---|---|---|---|---|---|
| PIKE | 1487 | N 12°10.531′ E 108°39.863′ | 40.3 ± 2.2 | 24.8 ± 1.0 | 290 | 16 |
| PIDA1 | 1494 | N 12°11.049′ E 108°41.526′ | 60.3 ± 0.5 | 30.0 ± 1.0 | 60 | 5 |
| PIDA2 | 1525 | N 12°10.885′ E 108°40.394′ | 54.7 ± 2.0 | 24.3 ± 1.3 | 70 | 5 |
| PIDA3 | 1535 | N 12°11.453′ E 108°40.762′ | 44.1 ± 1.2 | 23.8 ± 1.3 | 90 | 7 |
| PIDA4 | 1546 | N 12°12.094′ E 108°40.642′ | 57.4 ± 2.1 | 24.9 ± 0.9 | 70 | 6 |
| Species | Resistance (Rt) | Recovery (Rc) | Resilience (Rs) | Relative Resilience (rRs) |
|---|---|---|---|---|
| P. kesiya | 0.70 ± 0.02 | 1.48 ± 0.08 | 1.03 ± 0.06 | 0.33 ± 0.05 |
| P. dalatensis | 0.67 ± 0.02 | 1.51 ± 0.06 | 0.96 ± 0.03 | 0.29 ± 0.03 |
| Species | Fixed Effects | Rt | Rc | Rs | rRs |
|---|---|---|---|---|---|
| P. kesiya | PreEps | −0.09 | −0.16 | −0.16 | −0.15 |
| Age | −0.17 | −0.64 ** | −0.64 ** | −0.69 ** | |
| dbh | −0.04 | −0.36 * | −0.32 * | −0.37 * | |
| P. dalatensis | PreEps | −0.16 | −0.19 | −0.39 * | −0.30 * |
| Age | 0.05 | −0.28 * | −0.23 * | −0.29 * | |
| dbh | −0.08 | −0.23 * | −0.31 * | −0.28 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, L.T.; Hoppe, J.; Thomas, F.M. Resistance, Recovery and Resilience of Two Co-Occurring Palaeotropical Pinus Species Differing in the Sizes of Their Distribution Areas. Forests 2021, 12, 511. https://doi.org/10.3390/f12040511
Ho LT, Hoppe J, Thomas FM. Resistance, Recovery and Resilience of Two Co-Occurring Palaeotropical Pinus Species Differing in the Sizes of Their Distribution Areas. Forests. 2021; 12(4):511. https://doi.org/10.3390/f12040511
Chicago/Turabian StyleHo, Le T., Jana Hoppe, and Frank M. Thomas. 2021. "Resistance, Recovery and Resilience of Two Co-Occurring Palaeotropical Pinus Species Differing in the Sizes of Their Distribution Areas" Forests 12, no. 4: 511. https://doi.org/10.3390/f12040511