
Predicting the Potential Distribution of Perennial Plant Coptis chinensis Franch. in China under Multiple Climate Change Scenarios

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
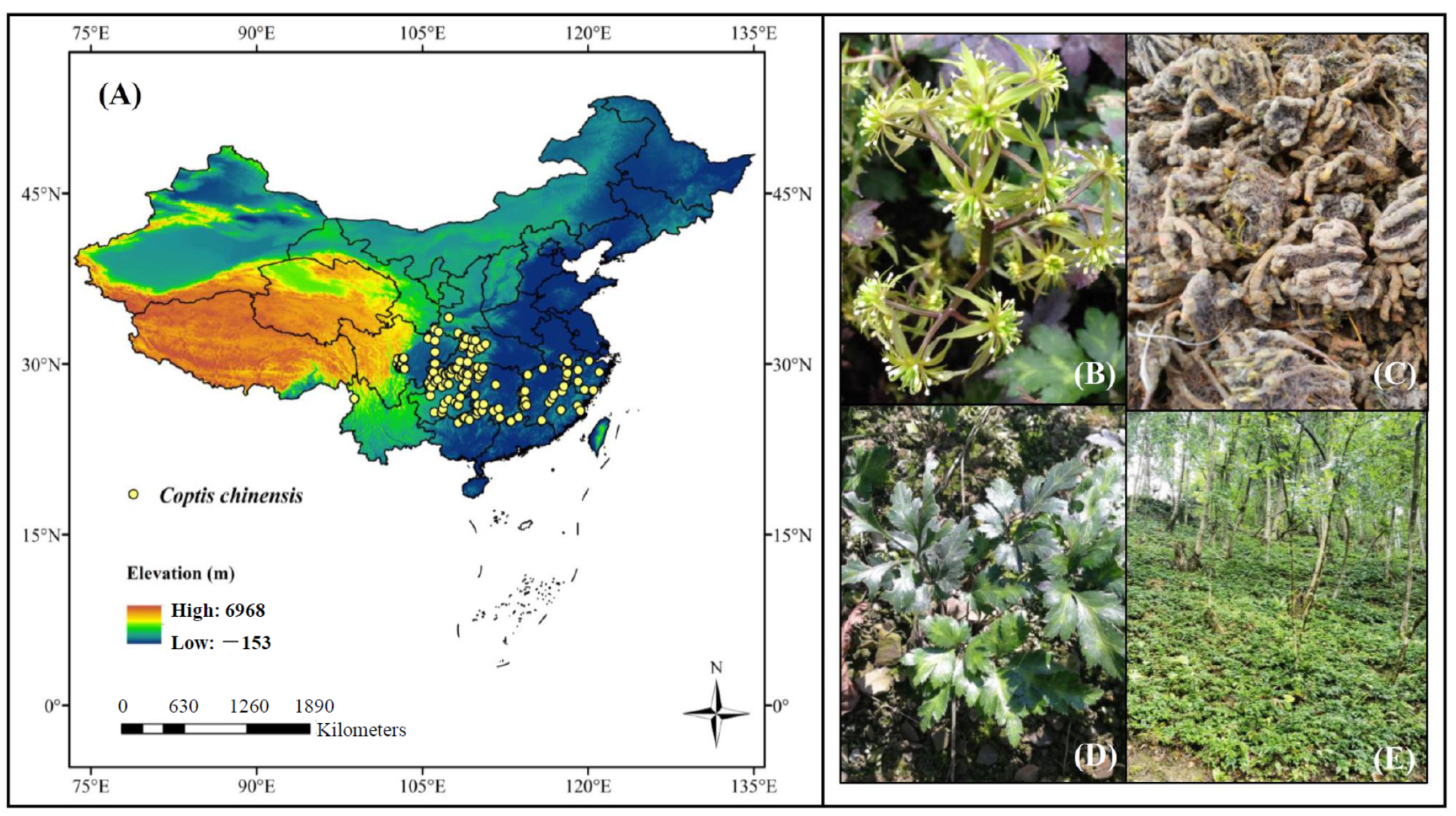
2.1. Study Area
2.2. Species Occurrence Data
2.3. Screening of Environmental Variables
2.4. Model Optimization and Establishment
2.5. Model Reliability Test and Classification of Suitable Regions
2.6. Analysis of Low Impact Area and Spatial Pattern Change
2.7. Core Distributional Shifts
3. Results
3.1. Model Parameter Optimization and Accuracy Analysis
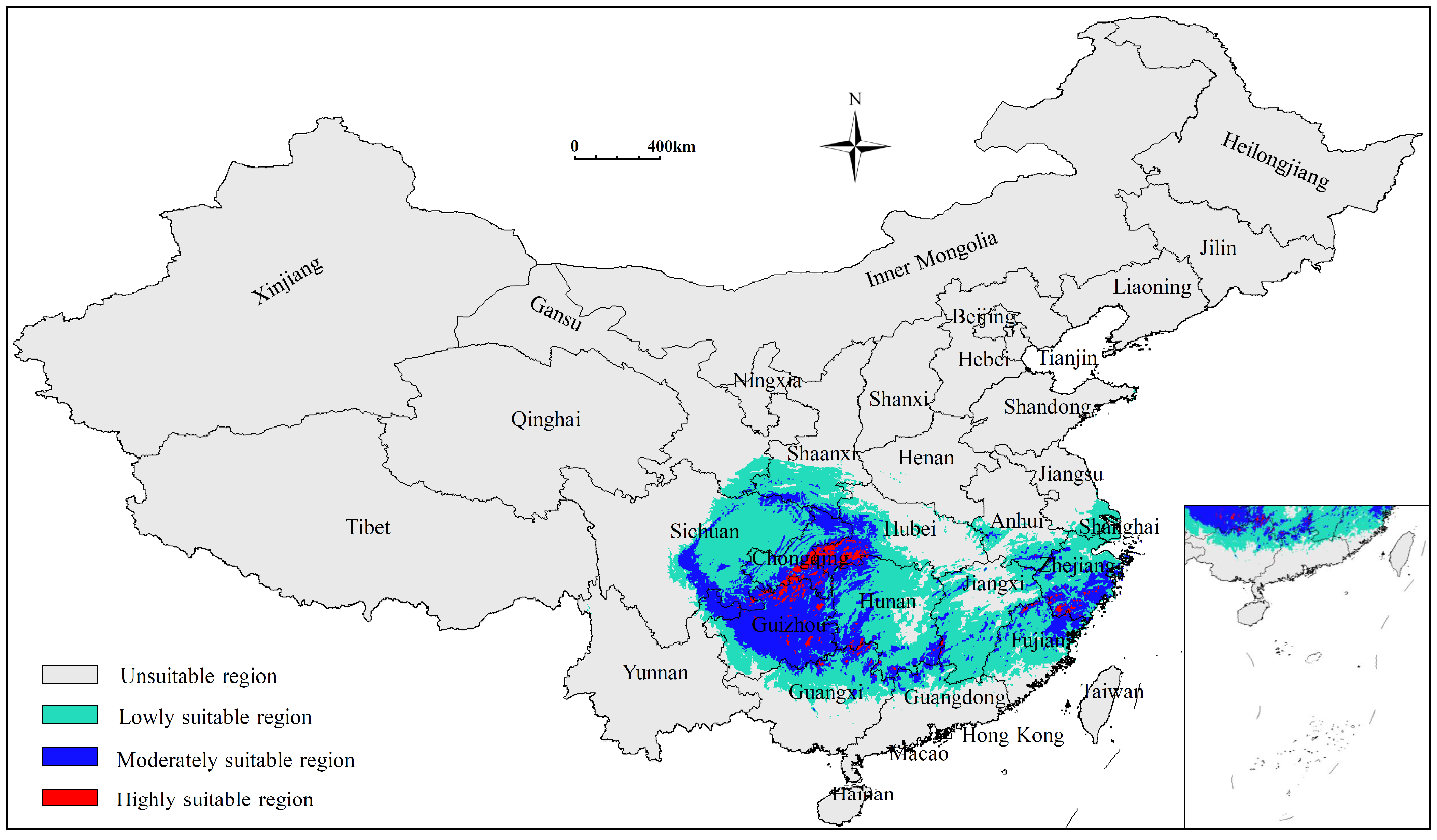
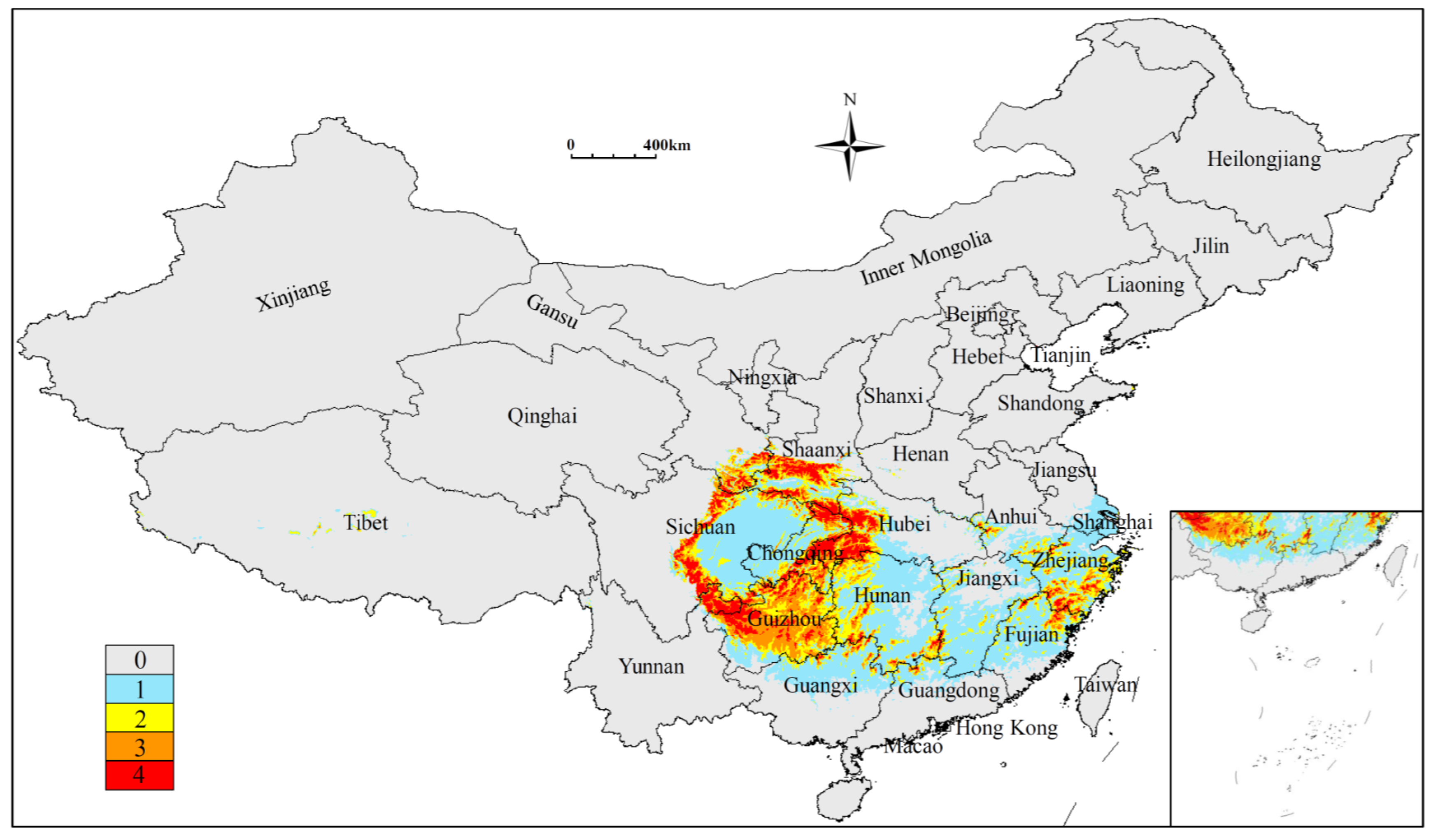
3.2. Current Potentially Suitable Region
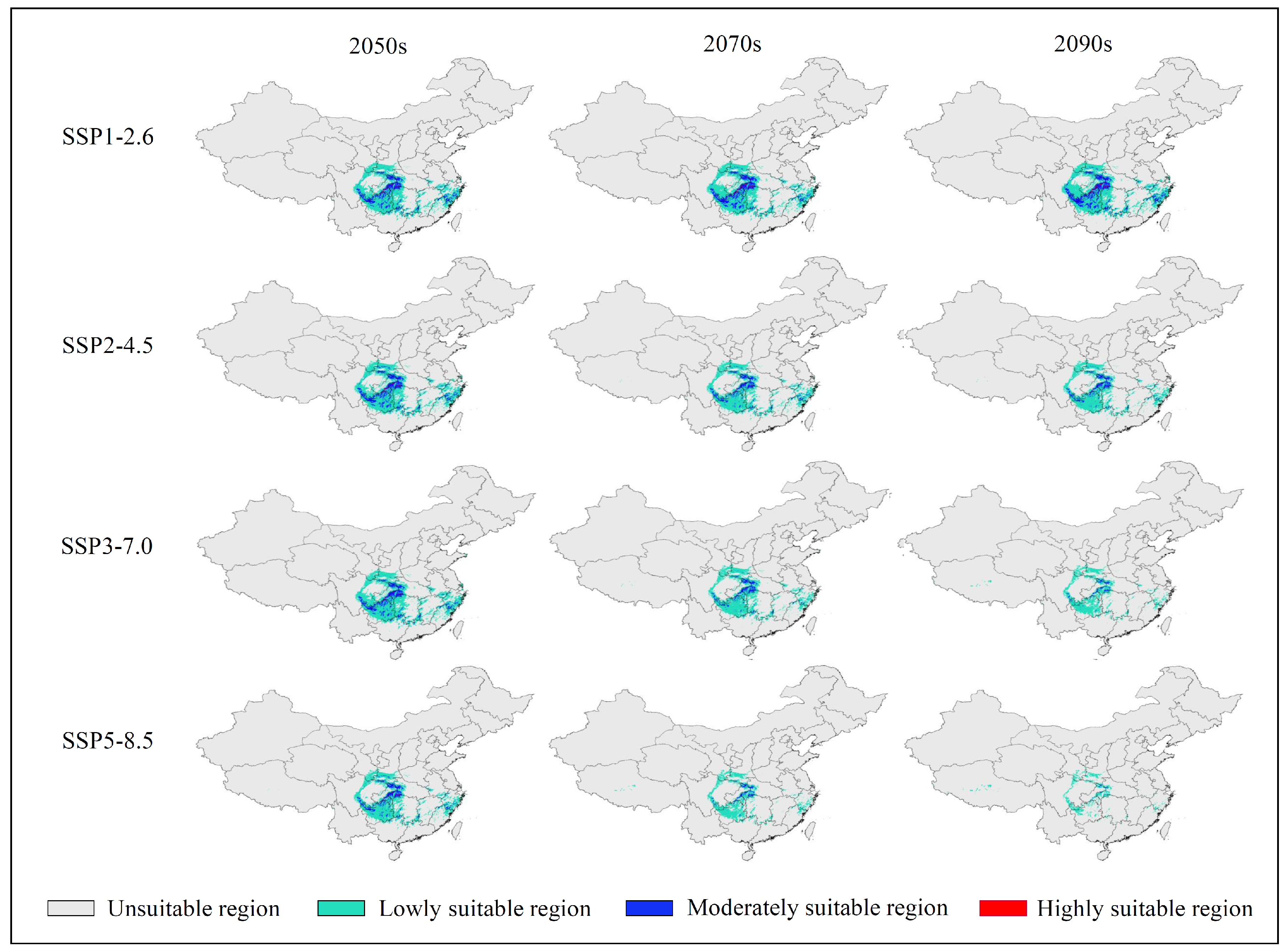
3.3. Future Potentially Suitable Regions
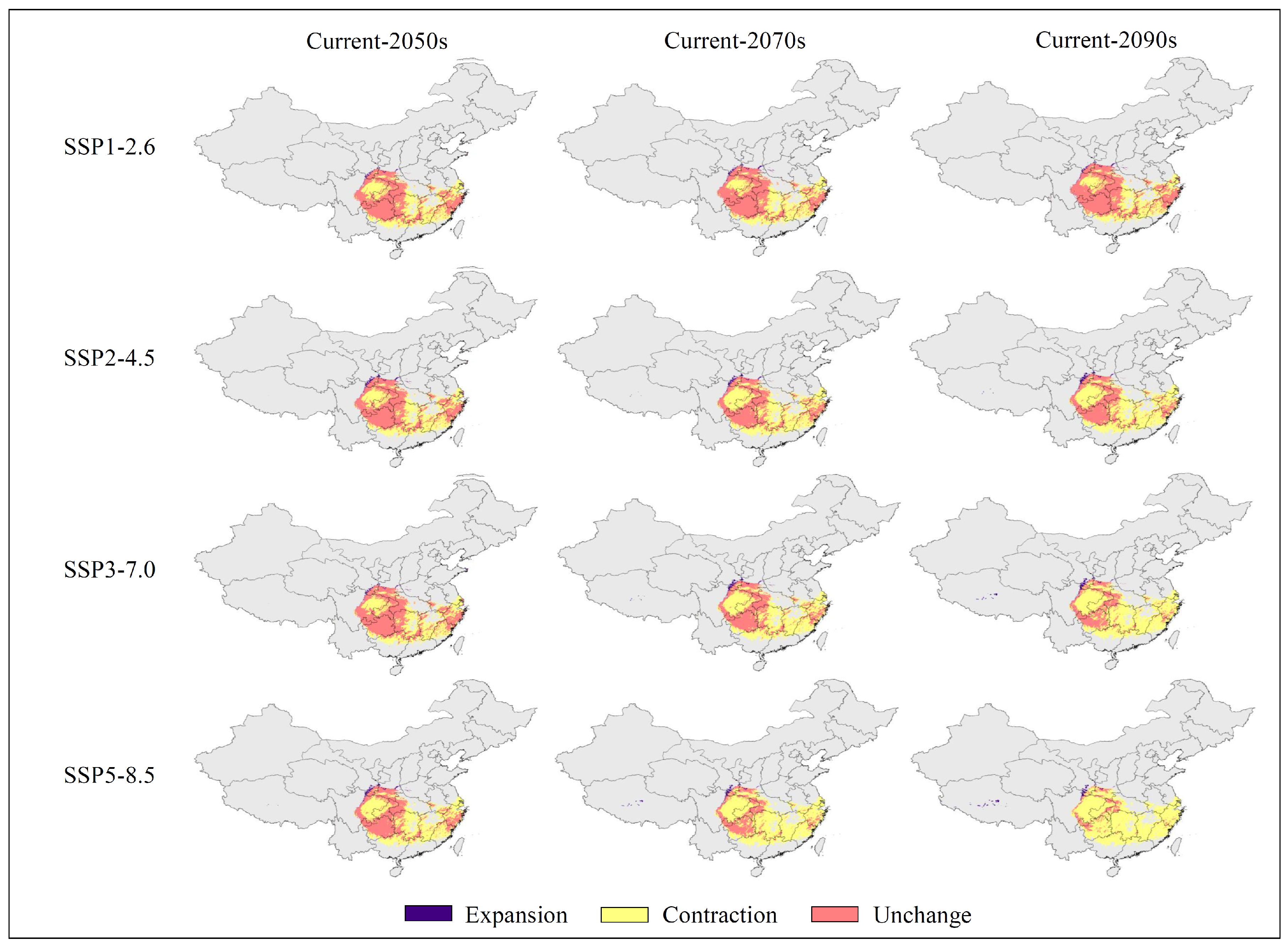
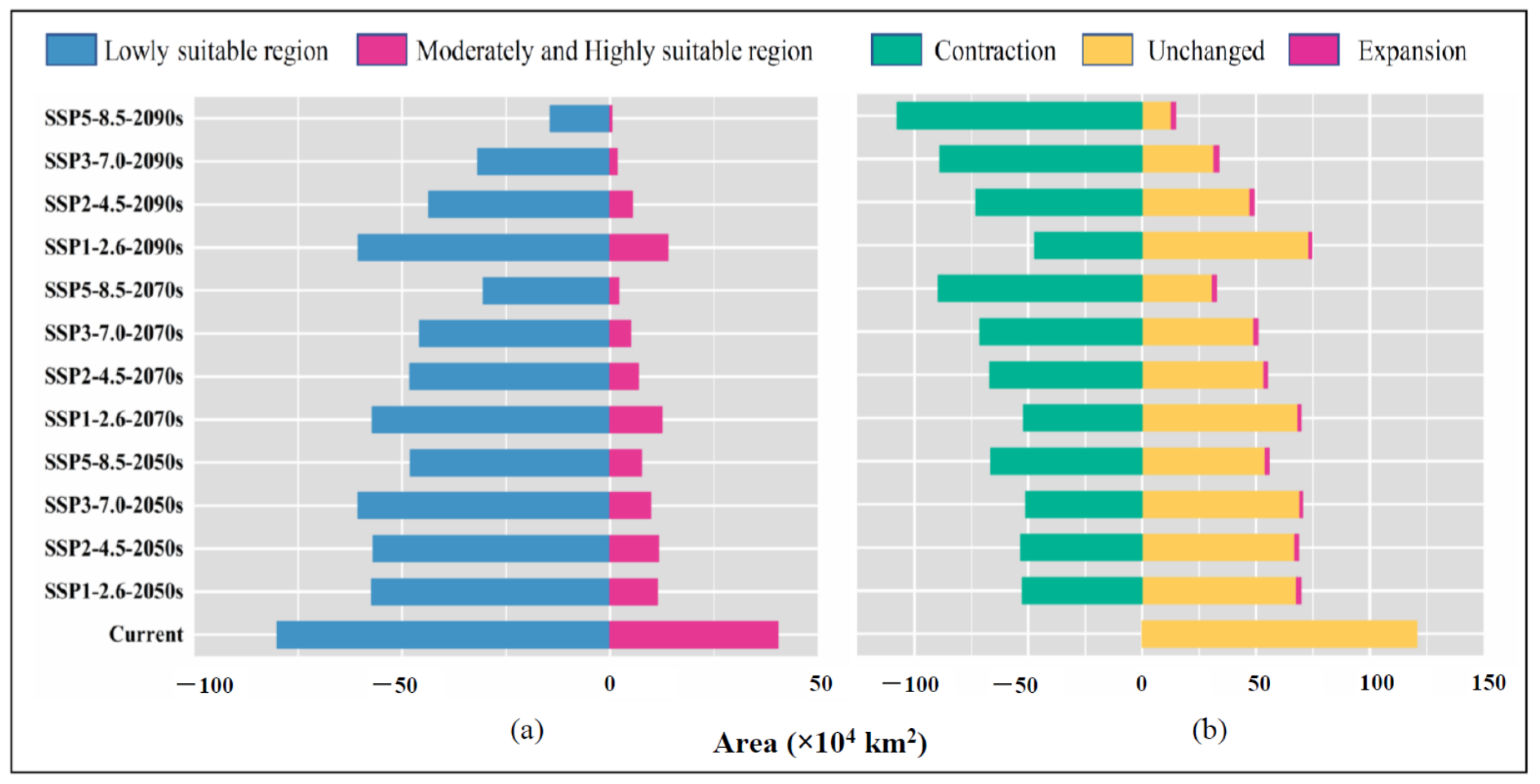
3.4. Low Impact Areas
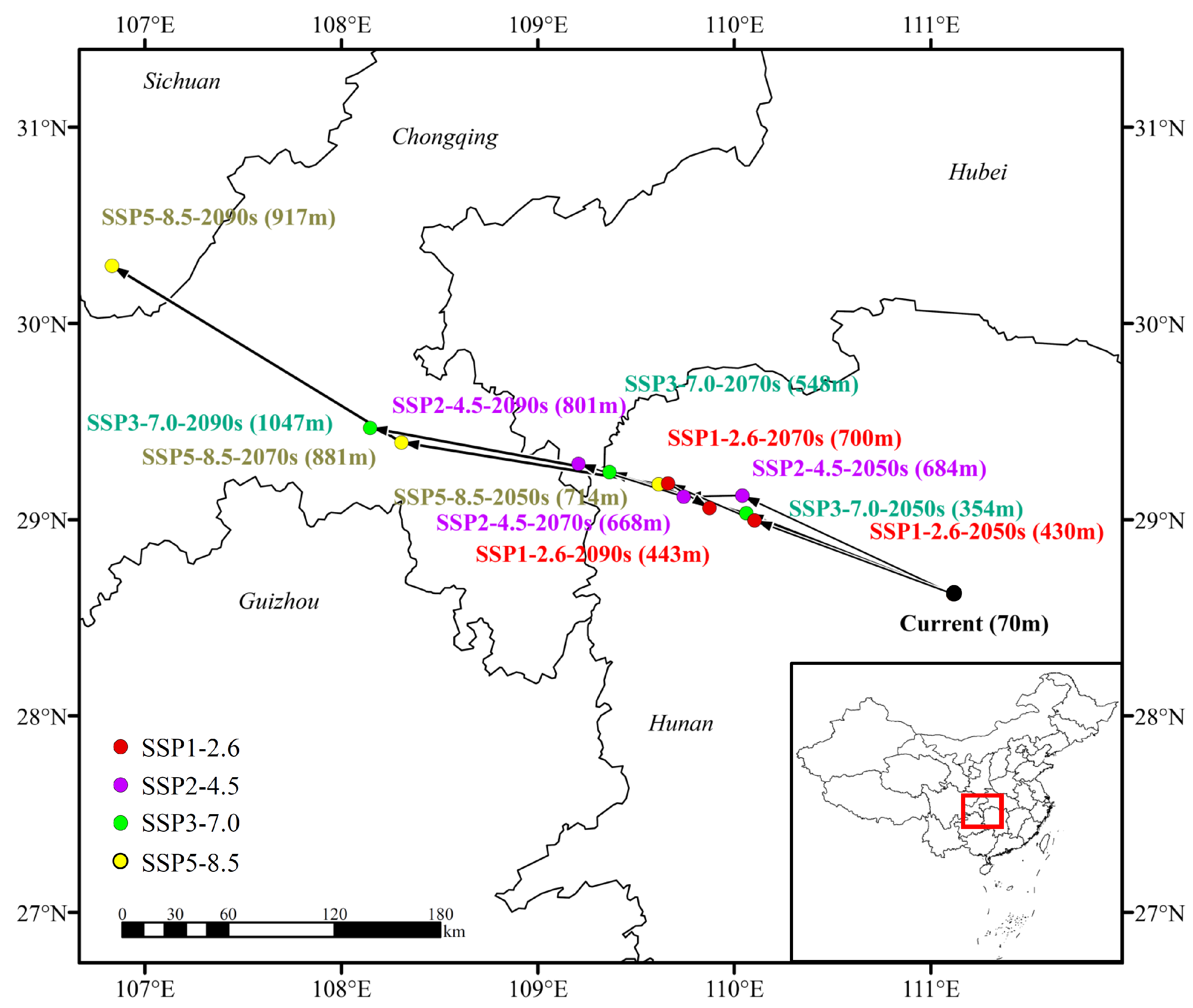
3.5. Shift in the Distribution Center of the Suitable Region
4. Discussion
4.1. Effects of Climate Change on Species Distribution and Migration Trends
4.2. Effects of Environmental Variables on Species Distribution
4.3. Protection of Genetic Resources
4.4. Rationality and Prediction Model Defects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Z.; Bai, Y.; Alatalo, J.M.; Huang, Z.; Yang, F.; Pu, X.; Wang, R.; Yang, W.; Guo, X. Spatio-temporal variation in potential habitats for rare and endangered plants and habitat conservation based on the maximum entropy model. Sci. Total Environ. 2021, 784, 147080. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.-Z.; Zhao, G.-H.; Zhang, M.-Z.; Cui, X.-Y.; Fan, H.-H.; Liu, B. Distribution pattern of endangered plant Semiliquidambar cathayensis (Hamamelidaceae) in response to climate change after the last interglacial period. Forests 2020, 11, 434. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Wu, Y.-M.; Shen, X.-L.; Tong, L.; Lei, F.-W.; Mu, X.-Y.; Zhang, Z.-X. Impact of past and future climate change on the potential distribution of an endangered montane shrub Lonicera oblata and its conservation implications. Forests 2021, 12, 125. [Google Scholar] [CrossRef]
- Hu, J.; Jiang, Z. Predicting the potential distribution of the endangered Przewalski’s gazelle. J. Zool. 2010, 282, 54–63. [Google Scholar] [CrossRef]
- Xu, W.; Sun, H.; Jin, J.; Cheng, J. Predicting the potential distribution of apple canker pathogen (Valsa mali) in China under climate change. Forests 2020, 11, 1126. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.; Shi, Y.; Wang, G.; Wang, T. Predicting suitable habitats of Camptotheca acuminata considering both climatic and soil variables. Forests 2020, 11, 891. [Google Scholar] [CrossRef]
- Santos-Hernández, A.F.; Monterroso-Rivas, A.I.; Granados-Sánchez, D.; Villanueva-Morales, A.; Santacruz-Carrillo, M. Projections for Mexico’s tropical rainforests considering ecological niche and climate change. Forests 2021, 12, 119. [Google Scholar] [CrossRef]
- Zhang, K.; Sun, L.; Tao, J. Impact of climate change on the distribution of Euscaphis japonica (Staphyleaceae) trees. Forests 2020, 11, 525. [Google Scholar] [CrossRef]
- Xu, D.; Zhuo, Z.; Wang, R.; Ye, M.; Pu, B. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling. Glob. Ecol. Conserv. 2019, 19, e00691. [Google Scholar] [CrossRef]
- Wei, J.; Li, X.; Lu, Y.; Zhao, L.; Zhang, H.; Zhao, Q. Modeling the potential global distribution of Phenacoccus madeirensis Green under various climate change scenarios. Forests 2019, 10, 773. [Google Scholar] [CrossRef] [Green Version]
- Solhjouy-Fard, S.; Sarafrazi, A.; Moeini, M.M.; Ahadiyat, A. Predicting habitat distribution of five heteropteran pest species in Iran. J. Insect Sci. 2013, 13, 116. [Google Scholar] [CrossRef] [Green Version]
- Peterman, W.E.; Crawford, J.A.; Kuhns, A.R. Using species distribution and occupancy modeling to guide survey efforts and assess species status. J. Nat. Conserv. 2013, 21, 114–121. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Zhao, P.; Zhou, H.-J.; Potter, D.; Hu, Y.-H.; Feng, X.-J.; Dang, M.; Feng, L.; Zulfiqar, S.; Liu, W.-Z.; Zhao, G.-F.; et al. Population genetics, phylogenomics and hybrid speciation of Juglans in China determined from whole chloroplast genomes, transcriptomes, and genotyping-by-sequencing (GBS). Mol. Phylogenet. Evol. 2018, 126, 250–265. [Google Scholar] [CrossRef]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Title, P.O.; Bemmels, J.B. ENVIREM: An expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 2018, 41, 291–307. [Google Scholar] [CrossRef] [Green Version]
- Sony, R.K.; Sen, S.; Kumar, S.; Sen, M.; Jayahari, K.M. Niche models inform the effects of climate change on the endangered Nilgiri Tahr (Nilgiritragus hylocrius) populations in the southern Western Ghats, India. Ecol. Eng. 2018, 120, 355–363. [Google Scholar] [CrossRef]
- Hoban, S. Integrative conservation genetics: Prioritizing populations using climate predictions, adaptive potential and habitat connectivity. Mol. Ecol. Resour. 2018, 18, 14–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Tao, J. Predicting the potential distribution of Paeonia veitchii (Paeoniaceae) in China by incorporating climate change into a maxent model. Forests 2019, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Kumar, S.; Stohlgren, T.J. Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar] [CrossRef]
- Alami, M.M.; Xue, J.; Ma, Y.; Zhu, D.; Abbas, A.; Gong, Z.; Wang, X. Structure, function, diversity, and composition of fungal communities in rhizospheric soil of Coptis chinensis Franch under a successive cropping system. Plants 2020, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Luo, Y.; Deng, D.; Su, S.; Li, S.; Xiang, L.; Hu, Y.; Wang, P.; Meng, X. Coptisine from Coptis chinensis exerts diverse beneficial properties: A concise review. J. Cell. Mol. Med. 2019, 23, 7946–7960. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Xiao, H.; Deng, C.; Fan, G.; Qin, S.; Peng, C. Complete chloroplast genome sequence of Coptis chinensis Franch. and its evolutionary history. Biomed Res. Int. 2017, 2017, 8201836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, J.; Fu, B.; Jiang, L.; Sun, T. Biosynthesis, characterization, and anticancer effect of plant-mediated silver nanoparticles using Coptis chinensis. Int. J. Nanomed. 2019, 14, 1969–1978. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Yun, X.; Yang, X.; Jia, S.; Jiao, C.; Shao, R.; Hao, J.; Chang, Y.; Fan, G.; Zhang, J.; et al. An inhibitory effect of Berberine from herbal Coptis chinensis Franch on rat detrusor contraction in benign prostatic hyperplasia associated with lower urinary tract symptoms. J. Ethnopharmacol. 2021, 268, 113666. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, B.; Shu, S.; Li, Z.; Song, C.; Liu, D.; Niu, Y.; Liu, J.; Zhang, J.; Liu, H.; et al. Analysis of the Coptis chinensis genome reveals the diversification of protoberberine-type alkaloids. Nat. Commun. 2021, 12, 3276. [Google Scholar] [CrossRef]
- Chen, D.X.; Pan, Y.; Wang, Y.; Cui, Y.Z.; Zhang, Y.J.; Mo, R.Y.; Wu, X.L.; Tan, J.; Zhang, J.; Guo, L.A.; et al. The chromosome-level reference genome of Coptis chinensis provides insights into genomic evolution and berberine biosynthesis. Hortic. Res. 2021, 8, 121. [Google Scholar] [CrossRef]
- Li, J.; Meng, X.; Wang, C.; Zhang, H.; Chen, H.; Deng, P.; Liu, J.; Huandike, M.; Wei, J.; Chai, L. Coptidis alkaloids extracted from Coptis chinensis Franch attenuate IFN-gamma-induced destruction of bone marrow cells. PLoS ONE 2020, 15, e0236433. [Google Scholar] [CrossRef]
- Yang, S.B.; Kim, E.H.; Kim, S.H.; Kim, Y.H.; Oh, W.; Lee, J.T.; Jang, Y.A.; Sabina, Y.; Ji, B.C.; Yeum, J.H. Electrospinning fabrication of poly(vinyl alcohol)/Coptis chinensis extract nanofibers for antimicrobial exploits. Nanomaterials 2018, 8, 734. [Google Scholar] [CrossRef] [Green Version]
- Azareh, A.; Rahmati, O.; Rafiei-Sardooi, E.; Sankey, J.B.; Lee, S.; Shahabi, H.; Ahmad, B.B. Modelling gully-erosion susceptibility in a semi-arid region, Iran: Investigation of applicability of certainty factor and maximum entropy models. Sci. Total Environ. 2019, 655, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Cui, B.; Duan, S.; Chen, J.; Fan, H.; Lu, B.; Zheng, J. Moving north in China: The habitat of Pedicularis kansuensis in the context of climate change. Sci. Total. Environ. 2019, 697, 133979. [Google Scholar] [CrossRef]
- Poirazidis, K.; Bontzorlos, V.; Xofis, P.; Zakkak, S.; Xirouchakis, S.; Grigoriadou, E.; Kechagioglou, S.; Gasteratos, I.; Alivizatos, H.; Panagiotopoulou, M. Bioclimatic and environmental suitability models for capercaillie (Tetrao urogallus) conservation: Identification of optimal and marginal areas in Rodopi Mountain-Range National Park (Northern Greece). Glob. Ecol. Conserv. 2019, 17, e00526. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Wu, D.; Zhu, C. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and MaxEnt. Glob. Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, L.; Wang, J.; Wang, W.; Niyati, N.; Guo, Y.; Wang, X. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci. Total Environ. 2021, 755, 142548. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Zhu, C.; Qu, Y.; Wang, H. Predicting the potential distribution of an invasive species, Erigeron canadensis L., in China with a maximum entropy model. Glob. Ecol. Conserv. 2020, 21, e00822. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, Y.; Huang, D.; Wang, H.; Cao, Q.; Fan, P.; Yang, N.; Zheng, P.; Wang, R. The effect of climate change on the richness distribution pattern of oaks (Quercus L.) in China. Sci. Total Environ. 2020, 744, 140786. [Google Scholar] [CrossRef] [PubMed]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas-Castro, S.; Goncalves, J.F.; Moreno, M.; Villar, R. Projected climate changes are expected to decrease the suitability and production of olive varieties in southern Spain. Sci. Total Environ. 2020, 709, 136161. [Google Scholar] [CrossRef]
- Akpoti, K.; Kabo-Bah, A.T.; Dossou-Yovo, E.R.; Groen, T.A.; Zwart, S.J. Mapping suitability for rice production in inland valley landscapes in Benin and Togo using environmental niche modeling. Sci. Total Environ. 2020, 709, 136165. [Google Scholar] [CrossRef]
- Liu, B.; Gao, X.; Ma, J.; Jiao, Z.; Xiao, J.; Hayat, M.A.; Wang, H. Modeling the present and future distribution of arbovirus vectors Aedes aegypti and Aedes albopictus under climate change scenarios in Mainland China. Sci. Total Environ. 2019, 664, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, X.; Zhao, Z.; Nawaz, Z. Predicting the impacts of climate change, soils and vegetation types on the geographic distribution of Polyporus umbellatus in China. Sci. Total Environ. 2019, 648, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.; Yang, Y.; Ohsawa, M.; Luu, H.T.; et al. Identifying long-term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Yu, X.; Yu, C.; Tayibazhaer, A.; Xu, F.; Skidmore, A.K.; Wang, T. Impacts of future climate and land cover changes on threatened mammals in the semi-arid Chinese Altai Mountains. Sci. Total Environ. 2018, 612, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Fan, X.; Luo, S.; Zhang, Y.; Yao, S.; Guo, Q.; Qian, Z. Predicting the potential distribution of two varieties of Litsea coreana (leopard-skin camphor) in China under climate change. Forests 2020, 11, 1159. [Google Scholar] [CrossRef]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Václavík, T.; Meentemeyer, R.K. Invasive species distribution modeling (iSDM): Are absence data and dispersal constraints needed to predict actual distributions? Ecol. Model. 2009, 220, 3248–3258. [Google Scholar] [CrossRef]
- Smith, A.B.; Godsoe, W.; Rodríguez-Sánchez, F.; Wang, H.-H.; Warren, D. Niche estimation above and below the species level. Trends Ecol. Evol. 2019, 34, 260–273. [Google Scholar] [CrossRef]
- Guo, Y.; Lu, Y.; El-Kassaby, Y.A.; Feng, L.; Wang, G.; Wang, T. Predicting growth and habitat responses of Ginkgo biloba L. to climate change. Ann. Forest Sci. 2019, 76, 101. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Feng, L.; Tang, X.; Bentley, Y.; Höök, M. The implications of fossil fuel supply constraints on climate change projections: A supply-side analysis. Futures 2017, 86, 58–72. [Google Scholar] [CrossRef]
- Li, J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef] [PubMed]
- Puchalka, R.; Dyderski, M.K.; Vitkova, M.; Sadlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.Y.; Sun, M.F.; Li, T.C.; Lin, C.T. Effect of Coptis chinensis on biofilm formation and antibiotic susceptibility in Mycobacterium abscessus. Evid-Based Compl. Alt. 2020, 2020, 9754357. [Google Scholar] [CrossRef]
- Velásquez-Tibatá, J.; Salaman, P.; Graham, C. Effects of climate change on species distribution, community structure, and conservation of birds in protected areas in Colombia. Reg. Environ. Change 2013, 13, 235–248. [Google Scholar] [CrossRef]
- Record, S.; Fitzpatrick, M.C.; Finley, A.O.; Veloz, S.; Ellison, A.M. Should species distribution models account for spatial autocorrelation? A test of model projections across eight millennia of climate change. Global Ecol. Biogeogr. 2013, 22, 760–771. [Google Scholar] [CrossRef] [Green Version]
- Maiorano, L.; Cheddadi, R.; Zimmermann, N.E.; Pellissier, L.; Petitpierre, B.; Pottier, J.; Laborde, H.; Hurdu, B.I.; Pearman, P.B.; Psomas, A.; et al. Building the niche through time: Using 13,000 years of data to predict the effects of climate change on three tree species in Europe. Global Ecol. Biogeogr. 2013, 22, 302–317. [Google Scholar] [CrossRef]
- Li, X.; Tian, H.; Wang, Y.; Li, R.; Song, Z.; Zhang, F.; Xu, M.; Li, D. Vulnerability of 208 endemic or endangered species in China to the effects of climate change. Reg. Environ. Change 2013, 13, 843–852. [Google Scholar] [CrossRef]
- Blank, L.; Blaustein, L. Using ecological niche modeling to predict the distributions of two endangered amphibian species in aquatic breeding sites. Hydrobiologia 2012, 693, 157–167. [Google Scholar] [CrossRef]
- Lee, J.W.; Kang, Y.J.; Choi, H.K.; Yoon, Y.G. Fractionated Coptis chinensis extract and its bioactive component suppress Propionibacterium acnes-stimulated inflammation in human keratinocytes. J. Microbiol. Biotechnol. 2018, 28, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Choi, J.S.; Jeong, C.S. Inhibitory Activities of palmatine from Coptis chinensis against Helicobactor pylori and gastric damage. Toxicol. Res. 2014, 30, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Friedemann, T.; Otto, B.; Klatschke, K.; Schumacher, U.; Tao, Y.; Leung, A.K.; Efferth, T.; Schroder, S. Coptis chinensis Franch. exhibits neuroprotective properties against oxidative stress in human neuroblastoma cells. J. Ethnopharmacol. 2014, 155, 607–615. [Google Scholar] [CrossRef]
- Yu, Q.; Tong, S.; Yan, J.; Hong, C.; Zhai, W.; Li, Y. Preparative separation of quaternary ammonium alkaloids from Coptis chinensis Franch by pH-zone-refining counter-current chromatography. J. Sep. Sci. 2011, 34, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Tu, D.; Ye, X.; Wu, J. Hypoglycemic and hypocholesterolemic effects of Coptis chinensis Franch inflorescence. Plant Food. Hum. Nutr. 2006, 61, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Luoto, M. The importance of biotic interactions for modelling species distributions under climate change. Global Ecol. Biogeogr. 2007, 16, 743–753. [Google Scholar] [CrossRef]
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species–climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- García-Valdés, R.; Zavala, M.A.; Araújo, M.B.; Purves, D.W. Chasing a moving target: Projecting climate change-induced shifts in non-equilibrial tree species distributions. J. Ecol. 2013, 101, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Low, B.W.; Yeo, D.C.J. Novel methods to select environmental variables in MaxEnt: A case study using invasive crayfish. Ecol. Model. 2016, 341, 5–13. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Variable | Percent Contribution (%) |
|---|---|---|
| BIO02 | Mean diurnal range (Mean of monthly (max temp–min temp)) (°C) | 36.2 |
| BIO04 | Temperature seasonality (standard deviation × 100) | 5.1 |
| BIO06 | Min temperature of coldest month (°C) | 41.4 |
| BIO10 | Mean temperature of warmest quarter (°C) | 2.1 |
| BIO11 | Mean temperature of coldest quarter (°C) | 12.2 |
| BIO14 | Precipitation of driest month (mm) | 3.0 |
| Decades | Predicted Area (× 104 km2) and % of the Corresponding Current Area | ||||
|---|---|---|---|---|---|
| Total Suitable Region | Lowly Suitable Region | Moderately Suitable Region | Highly Suitable Region | ||
| 1970–2000 | 120.47 | 80.10 | 37.16 | 3.21 | |
| SSP1-2.6 | 2050s | 68.66 | 57.30 | 11.05 | 0.30 |
| (56.99%) | (71.53%) | (29.74%) | (9.51%) | ||
| 2070s | 69.64 | 57.17 | 12.02 | 0.45 | |
| (57.81%) | (71.38%) | (32.34%) | (14.00%) | ||
| 2090s | 74.47 | 60.53 | 13.50 | 0.44 | |
| (61.82%) | (75.57%) | (36.33%) | (13.78%) | ||
| SSP2-4.5 | 2050s | 68.72 | 56.98 | 11.40 | 0.35 |
| (57.05%) | (71.14%) | (30.66%) | (10.92%) | ||
| 2070s | 54.98 | 48.17 | 6.68 | 0.14 | |
| (45.64%) | (60.14%) | (17.96%) | (4.22%) | ||
| 2090s | 49.08 | 43.64 | 5.36 | 0.07 | |
| (40.74%) | (54.49%) | (14.42%) | (2.27%) | ||
| SSP3-7.0 | 2050s | 70.40 | 60.58 | 9.65 | 0.18 |
| (58.44%) | (75.63%) | (25.97%) | (5.51%) | ||
| 2070s | 50.80 | 45.84 | 4.90 | 0.06 | |
| (42.17%) | (57.23%) | (13.20%) | (1.73%) | ||
| 2090s | 33.52 | 31.79 | 1.73 | 0.00 | |
| (27.83%) | (39.69%) | (4.65%) | (0.11%) | ||
| SSP5-8.5 | 2050s | 55.66 | 48.08 | 7.38 | 0.20 |
| (46.21%) | (60.02%) | (19.86%) | (6.38%) | ||
| 2070s | 32.63 | 30.50 | 2.12 | 0.02 | |
| (27.09%) | (38.07%) | (5.69%) | (0.49%) | ||
| 2090s | 14.69 | 14.28 | 0.41 | 0.00 | |
| (12.19%) | (17.83%) | (1.10%) | (0.00%) | ||
| LIA Statistics | Shared Socio-Economic Pathways (SSPs) | |||
|---|---|---|---|---|
| SSP1-2.6 | SSP2-4.5 | SSP3-7.0 | SSP5-8.5 | |
| Geographic area (×104 km2) | 64.22 | 47.33 | 31.29 | 12.86 |
| Percentage of current suitable area (%) | 53.31 | 39.29 | 25.97 | 10.67 |
| Percentage of SSP1-2.6 area (%) | 100.00 | 73.70 | 48.72 | 20.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Q.; Zhang, Y.; Li, W.-N.; Hu, B.-W.; Zou, J.-B.; Wang, S.-Q.; Niu, J.-F.; Wang, Z.-Z. Predicting the Potential Distribution of Perennial Plant Coptis chinensis Franch. in China under Multiple Climate Change Scenarios. Forests 2021, 12, 1464. https://doi.org/10.3390/f12111464
Zhao Q, Zhang Y, Li W-N, Hu B-W, Zou J-B, Wang S-Q, Niu J-F, Wang Z-Z. Predicting the Potential Distribution of Perennial Plant Coptis chinensis Franch. in China under Multiple Climate Change Scenarios. Forests. 2021; 12(11):1464. https://doi.org/10.3390/f12111464
Chicago/Turabian StyleZhao, Qian, Yuan Zhang, Wen-Na Li, Bang-Wen Hu, Jia-Bin Zou, Shi-Qiang Wang, Jun-Feng Niu, and Zhe-Zhi Wang. 2021. "Predicting the Potential Distribution of Perennial Plant Coptis chinensis Franch. in China under Multiple Climate Change Scenarios" Forests 12, no. 11: 1464. https://doi.org/10.3390/f12111464