Intake of Radionuclides in the Trees of Fukushima Forests 3. Removal of Radiocesium from Stem Wood, Cryptomeria Japonica (L.f.) D. Don. †

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Stems for Ponding Method
2.2. Determination of Radioactivity
2.3. Microscopy
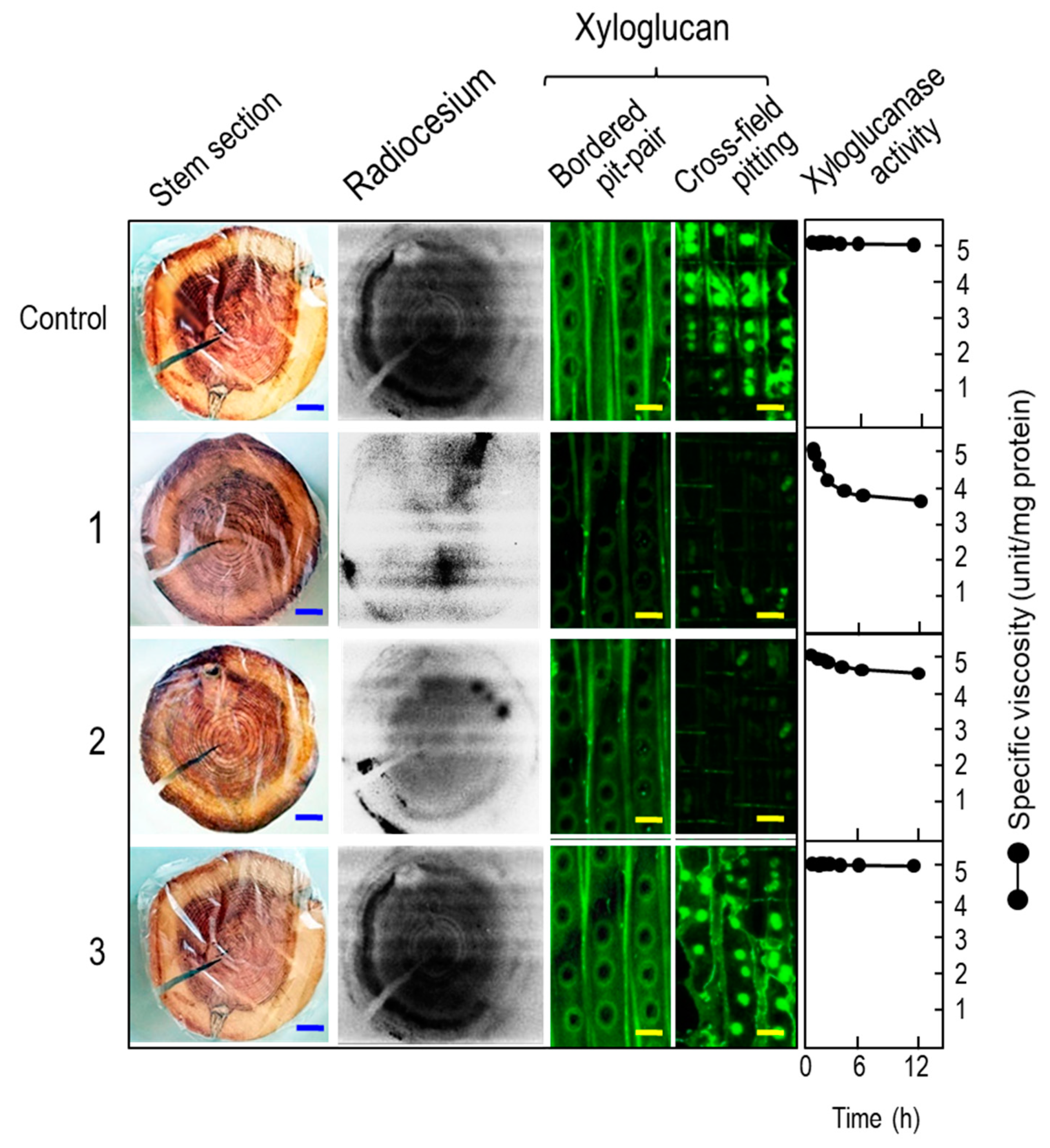
2.4. Determination of Xyloglucanase Activity
2.5. Potassium Fertilization
3. Results and Discussion
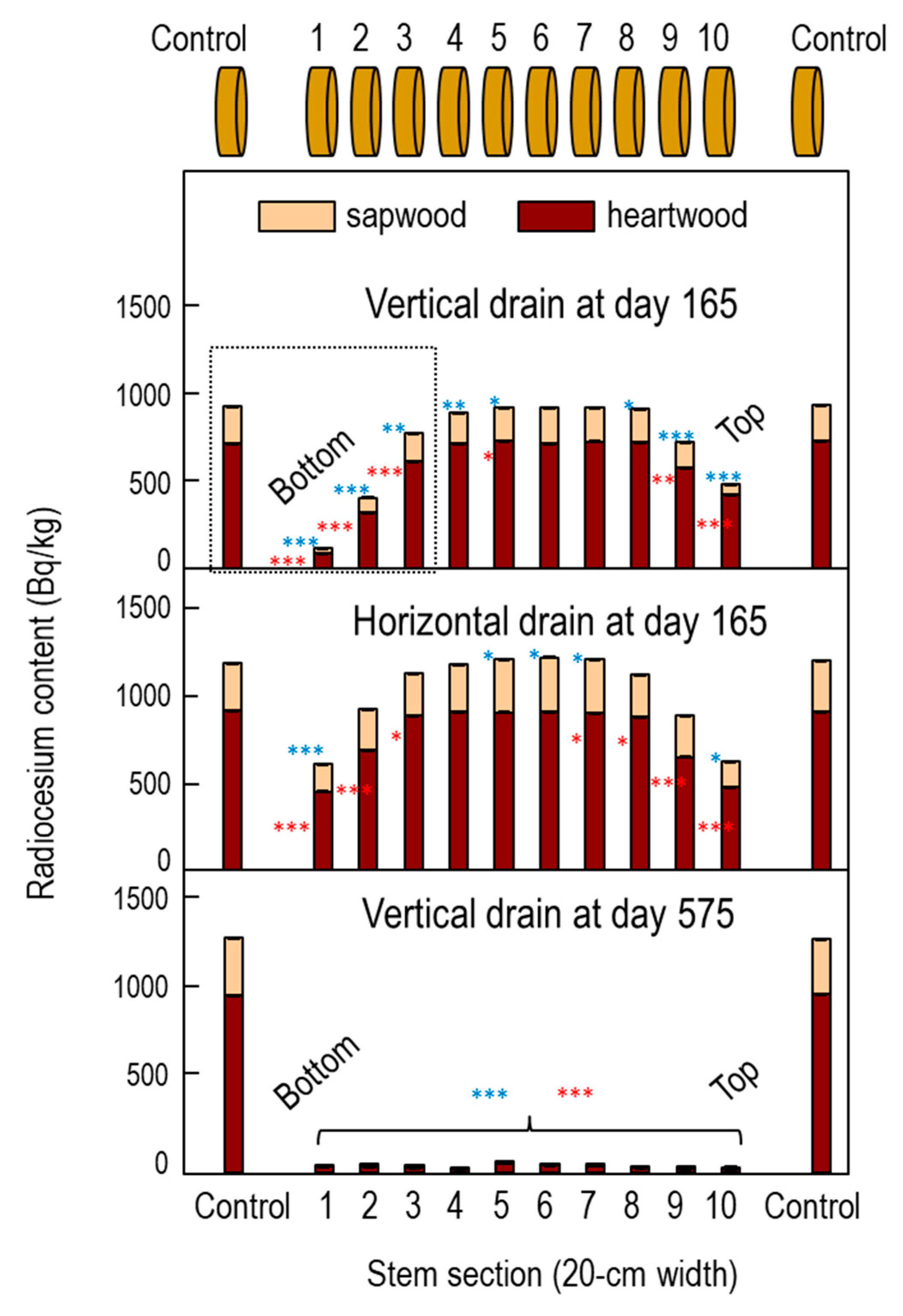
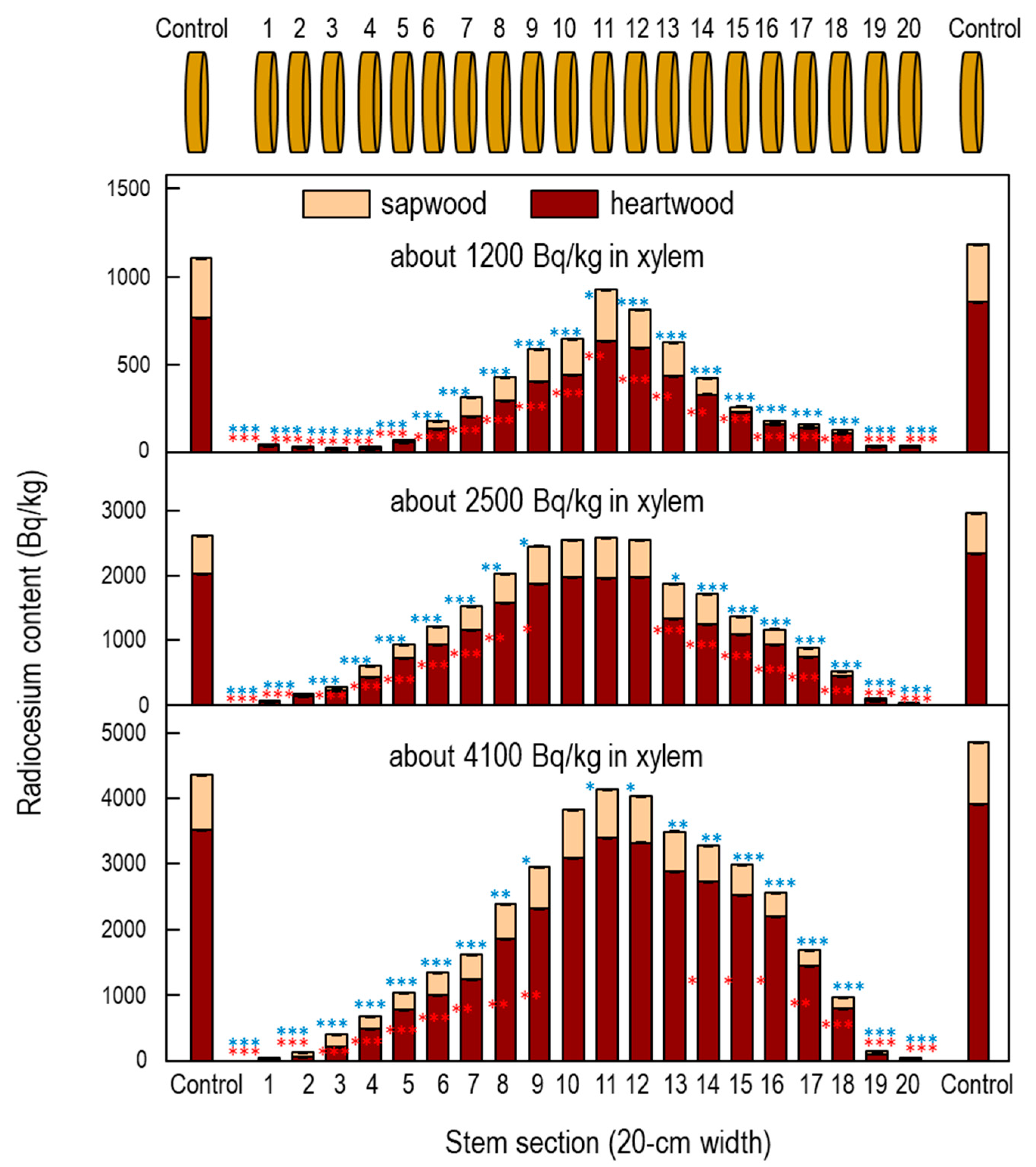
3.1. Ponding Experiments in Fukushima Water Pool
3.2. Analysis at the Cross-Cut Edge of Stem
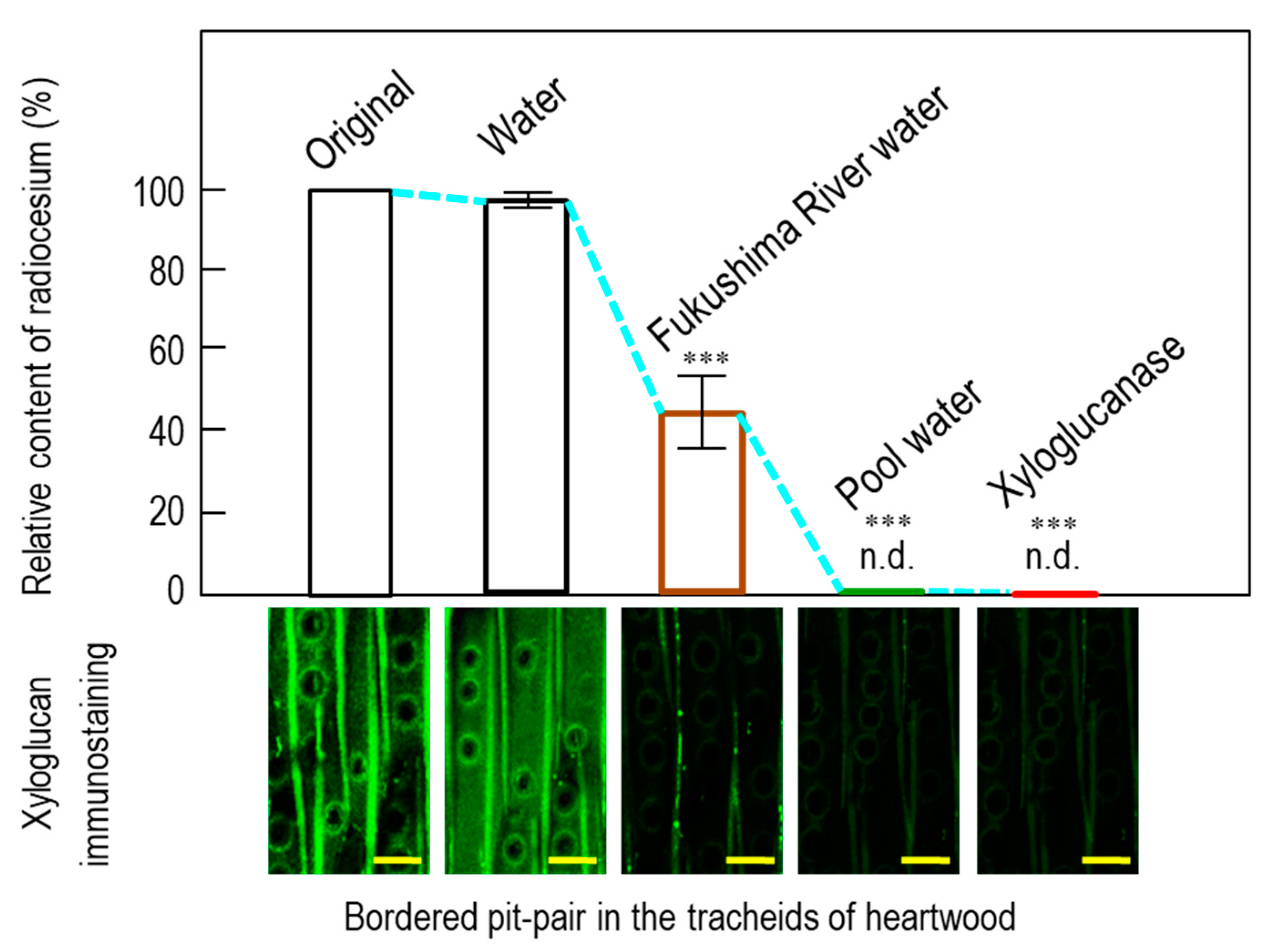
3.3. Lab-scale Ponding Experiments
3.4. Distribution of Radiocesium in Water Pool
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- The Fukushima Daiichi Accident. Reported by the Director General. IAEA. 2015. Available online: https://www-pub.iaea.org/MTCD/Publications/PDF/Pub1710-ReportByTheDG-Web.pdf (accessed on 8 May 2020).
- Kagawa, A.; Aoki, T.; Okada, N.; Katayama, Y. Tree-ring strontium-90 andcesium-137 as potentialindicatorsofradioactive pollution. J. Environ. Q. 2002, 31, 2001–2007. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Morino, Y.; Tanaka, A. Atmospheric behavior of radioactive materials from Fukushima Daiichi nuclear power plant. J. Nad. Inst. Public Health 2011, 60, 292–299. [Google Scholar]
- Jost, D.T.; Gaggeler, H.W.; Baltensperger, U. Chernobyl fallout in size-fractionated aerosol. Nature 1986, 324, 22–23. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, C.; Aoki, S.; Nonaka, M.; Itakura, M.; Tsubokura, M.; Baba, K.; Ohbayashi, H.; Sugawara, I.; Seyama, T.; Uehara, I.; et al. Intake of radionuclides in the trees of Fukushima forests. 1. Field study. Forests 2019, 10, 652. [Google Scholar] [CrossRef] [Green Version]
- Aoki, S.; Nonaka, M.; Yasukawa, C.; Itakura, M.; Tsubokura, M.; Baba, K.; Ohbayashi, H.; Seyama, T.; Uehara, I.; Kaida, R.; et al. Intake of radionuclides in the trees of Fukushima forests. 2. Study of radiocesium flow to poplar seedlings as a model tree. Forests 2019, 10, 736. [Google Scholar] [CrossRef] [Green Version]
- Kato, N.; Kihou, N.; Fujimura, S.; Ikeba, M.; Miyazaki, N.; Saito, Y.; Eguchi, T.; Itoh, S. Potassium fertilizer and other materials as countermeasures to reduce radiocesium levels in rice: Results of urgent experiments in 2011 responding to the Fukushima Daiichi nuclear power plant accident. Soil Sci. Plant Nut. 2015, 61, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Fukushima Mokuren. Available online: http://www.aburin.net/PDF_24/241113 (accessed on 8 May 2020). (In Japanese).
- McCormick, K.; Kaberger, T. Exploring a pioneering bioenergy system: The case of Enkoping in Sweden. J. Clean. Prod. 2005, 13, 1003–1014. [Google Scholar] [CrossRef]
- Liese, W.; Karnop, G. On the attack of coniferous wood by bacteria. Holz. RohWerkstoff 1968, 26, 202–208. [Google Scholar] [CrossRef]
- Unligil, H. Penetrability and strength of white spruce after ponding. Forest Prod. J. 1972, 22, 92–97. [Google Scholar]
- Kobayashi, Y. Drying characteristics of sugi wood and an attempt of its improvement. Wood Sci. Technol. 1991, 5, 43–44. [Google Scholar]
- Klaassen, R.K. Effect of Ponding on the Wood Quality of Scots Pine. In Proceedings of the COST-Action IE-0610 Workshop in Hamburg; Available online: https://www.shr.nl/uploads/pdf-files/2010-10-00-cost-action-0601-ponded-pine-full-paper.pdf (accessed on 8 May 2020).
- Puhlmann, J.; Bucheli, E.; Swain, M.J.; Dunning, N.; Albersheim, P.; Darvill, A.G.; Hahn, M.G. Generation of monoclonal antibodies against plant cell-wall polysaccharides. Plant Physiol. 1994, 104, 699–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Wong, Y.-S.; Maclachlan, G. Pea xyloglucan and cellulose. II. Hydrolysis by pea endo-1,4-β-glucanases. Plant Physiol. 1984, 75, 605–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, Y.; Iida, I.; Imamura, Y.; Watanabe, U. Drying and anatomical characteristics of sugi wood attacked by bacteria during pond storage. J. Wood Sci. 1998, 44, 432–437. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Iida, I.; Imamura, Y.; Watanabe, U. Improvement of penetrability of sugi wood by impregnation of bacteria using sap-flow method. J. Wood Sci. 1998, 44, 482–485. [Google Scholar] [CrossRef]
- Klepsch, M.M.; Schmitt1, M.; Knox, J.P.; Jansen, S. The Chemical Identity of Intervessel Pit Membranes in Acer Challenges Hydrogel Control of Xylem Hydraulic Conductivity. AoBPlants. Available online: https://academic.oup.com/aobpla/article-abstract/doi/10.1093/aobpla/plw052/2609612 (accessed on 8 May 2020).
- Mimura, H.; Akiba, K. Adsorption behavior of cesium and strontium on synthetic zeolite P. J. Nucl. Sci. Technol. 1993, 30, 436–443. [Google Scholar] [CrossRef]
- Valent, B.S.; Albersheim, P. The structure of plant cell walls. V. On the binding of xyloglucan to cellulose fibers. Plant Physiol. 1974, 54, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Ogawa, K.; Mitsuishi, Y. Characterization of the adsorption of xyloglucan to cellulose. Plant Cell Physiol. 1994, 35, 1199–1205. [Google Scholar] [CrossRef]
- Zhao, Z.; Crespi, V.H.; Kubicki, J.D.; Cosgrove, D.J.; Zhonget, L. Molecular dynamics simulation study of xyloglucan adsorption on cellulose surfaces: Effects of surface hydrophobicity and side-chain variation. Cellulose 2014, 21, 1025–1039. [Google Scholar] [CrossRef]
- Yildiz, S.; Canakci, S.; Yieldiz, U.C.; Ozgenc, O.; Tomak, E.D. Improving of the impregnability of refractory spruce wood by Bacillus licheniformis pretreatment. BioResources 2012, 7, 565–577. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Radiocesium Level | Level of Adsorption | |
|---|---|---|---|
| Day 0 | Day 575 | ||
| kBq | % | ||
| Wood | 5100 | 2340 | – |
| Bark | 12,000 | 1 | – |
| Zeolite | 0 | 5400 | 31.5 |
| Water | 0 | 5200 | 30.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seyama, T.; Arakawa, R.; Machida, S.; Yoshida, S.; Maru, A.; Baba, K.; Kobayashi, Y.; Kaida, R.; Taji, T.; Sakata, Y.; et al. Intake of Radionuclides in the Trees of Fukushima Forests 3. Removal of Radiocesium from Stem Wood, Cryptomeria Japonica (L.f.) D. Don. Forests 2020, 11, 589. https://doi.org/10.3390/f11050589
Seyama T, Arakawa R, Machida S, Yoshida S, Maru A, Baba K, Kobayashi Y, Kaida R, Taji T, Sakata Y, et al. Intake of Radionuclides in the Trees of Fukushima Forests 3. Removal of Radiocesium from Stem Wood, Cryptomeria Japonica (L.f.) D. Don. Forests. 2020; 11(5):589. https://doi.org/10.3390/f11050589
Chicago/Turabian StyleSeyama, Tomoko, Ryohei Arakawa, Shogo Machida, Sota Yoshida, Akihiko Maru, Kei’ichi Baba, Yoshinori Kobayashi, Rumi Kaida, Teruaki Taji, Yoichi Sakata, and et al. 2020. "Intake of Radionuclides in the Trees of Fukushima Forests 3. Removal of Radiocesium from Stem Wood, Cryptomeria Japonica (L.f.) D. Don." Forests 11, no. 5: 589. https://doi.org/10.3390/f11050589