Effects of Hormones and Epigenetic Regulation on the Callus and Adventitious Bud Induction of Fraxinus mandshurica Rupr.



Abstract
:1. Introduction
2. Materials and Methods
2.1. Disinfection and Inoculation of F. mandshurica Seeds
2.2. Adventitious Bud Induction of Hypocotyl, Root, Cotyledon and Stem Segments of F. Mandshurica
2.3. Extraction and Analysis of Endogenous Hormones in Plants
2.4. DNA Demethylation and Histone Acetylation
2.5. Quantitative Real-Time PCR Analyses
3. Results
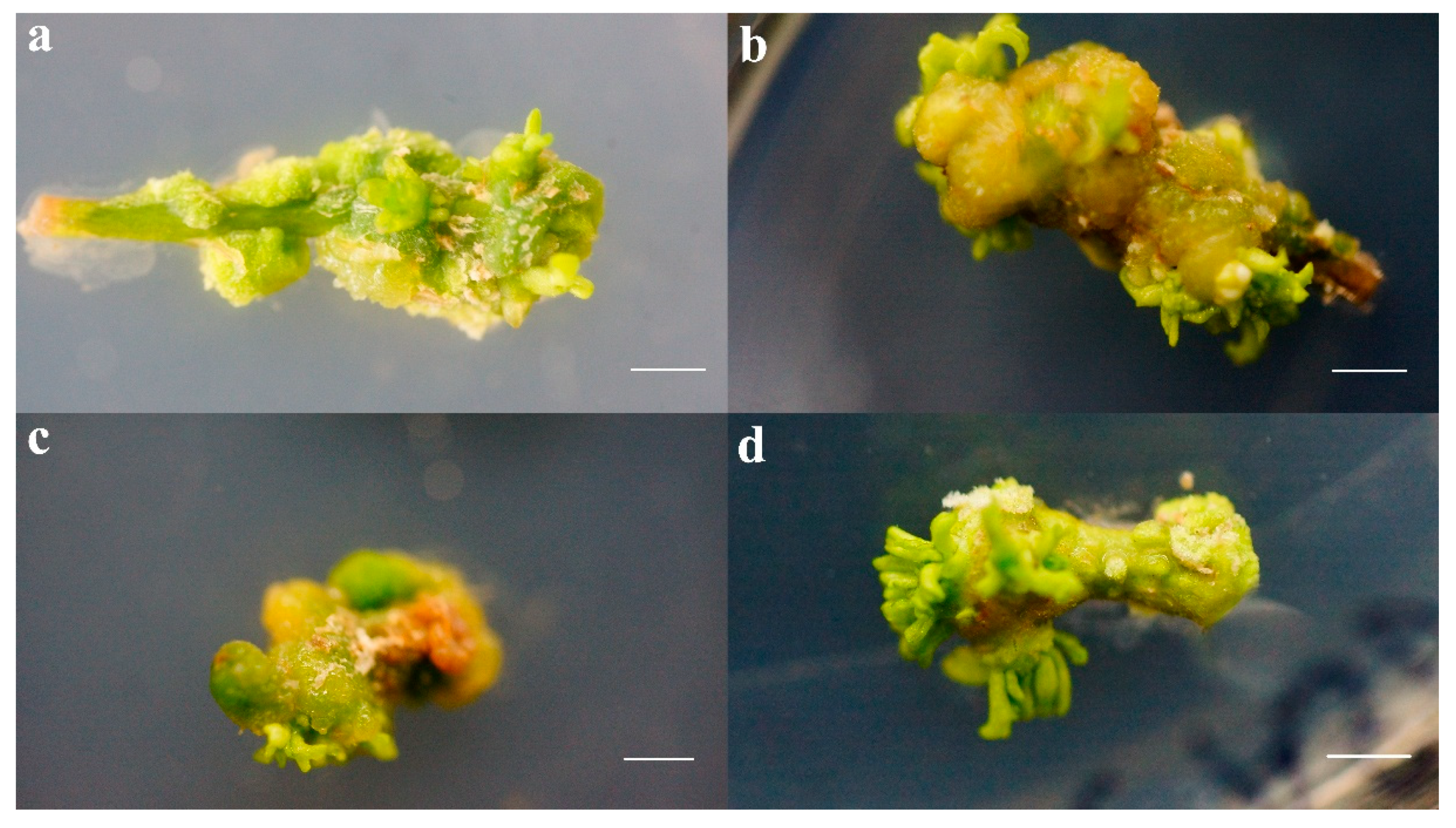
3.1. Effect of Exogenous 6-BA and TDZ Hormone Combination on Adventitious Bud Induction
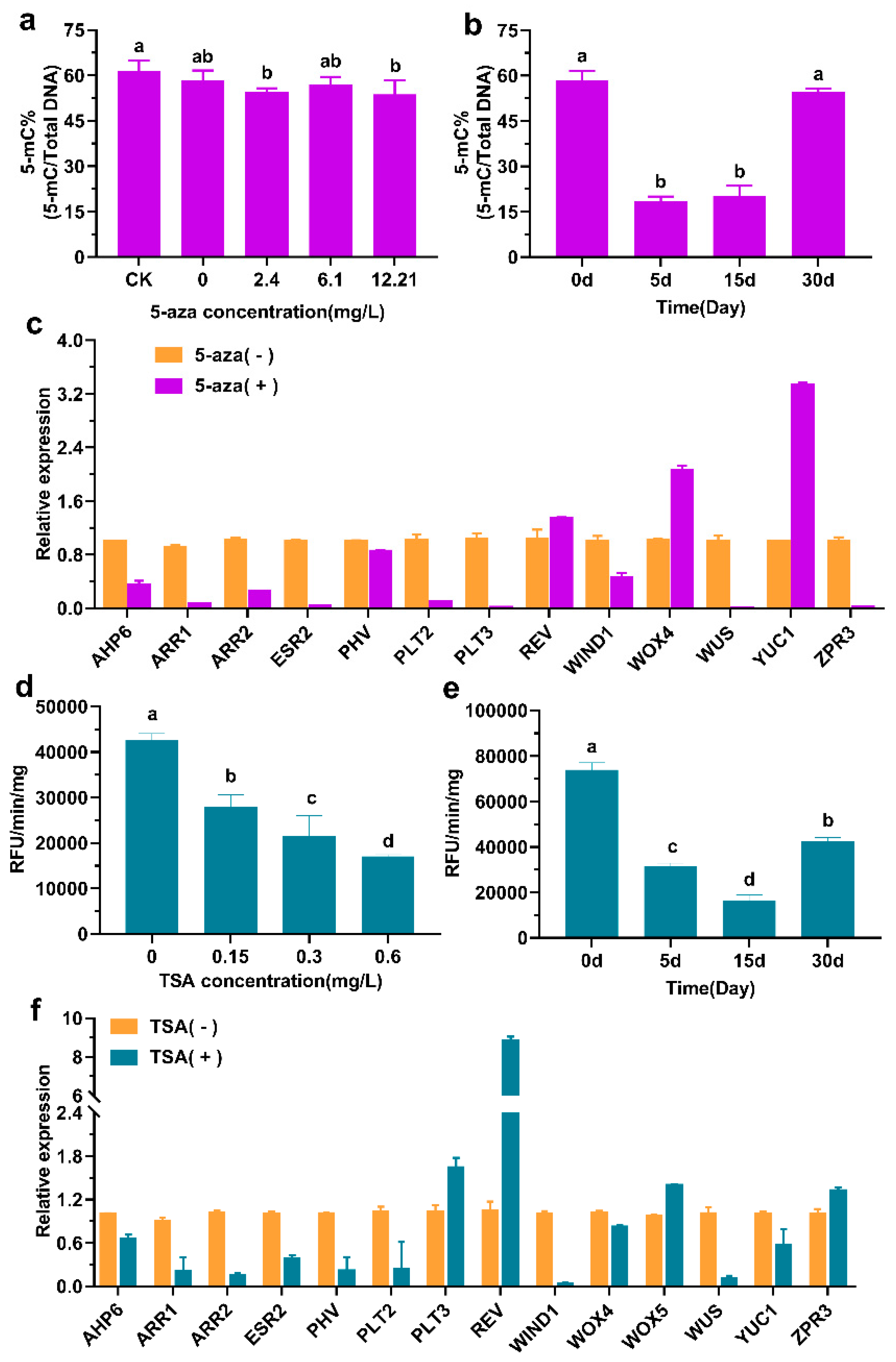
3.2. Effects of 5-aza and TSA on Adventitious Bud Differentiation of F. mandshurica
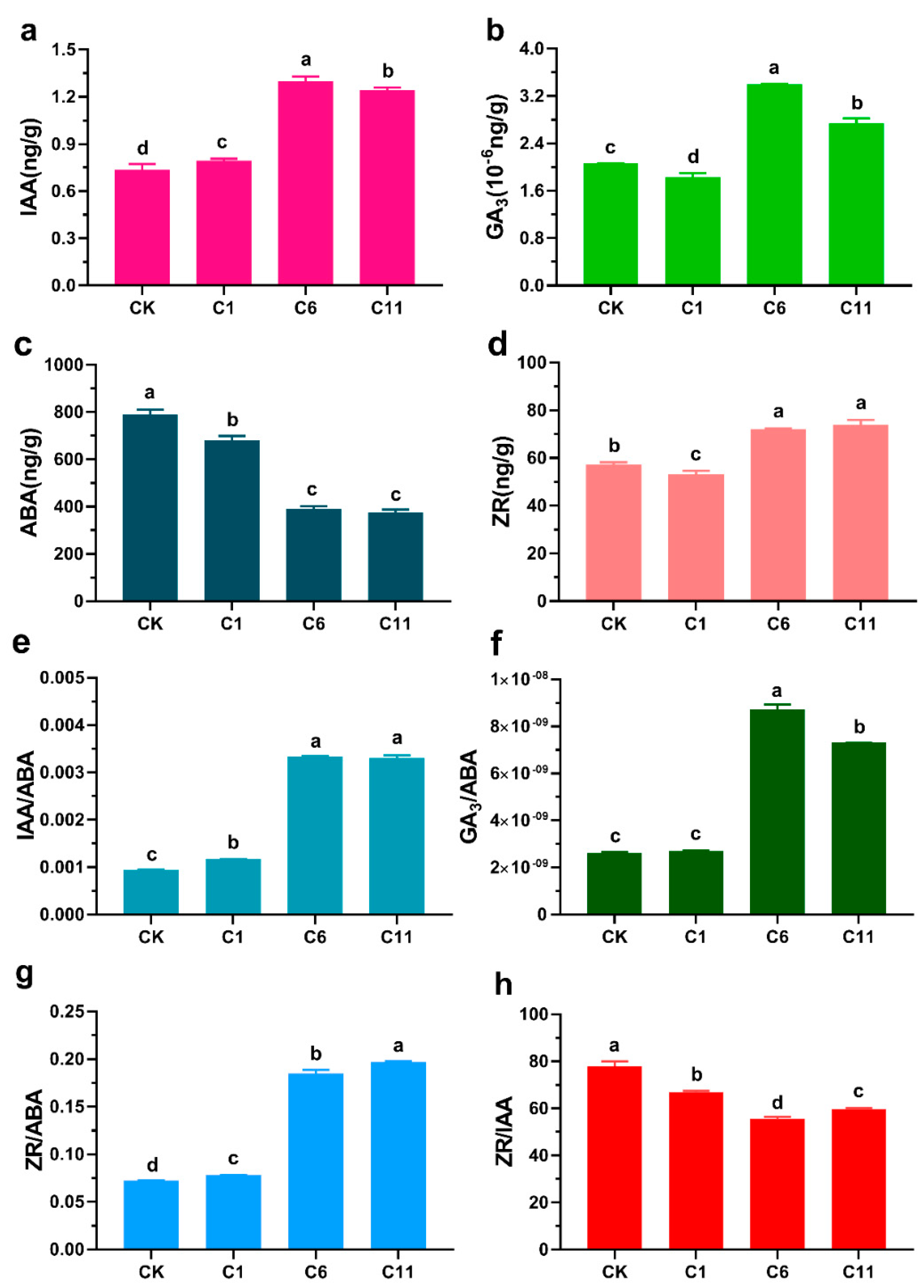
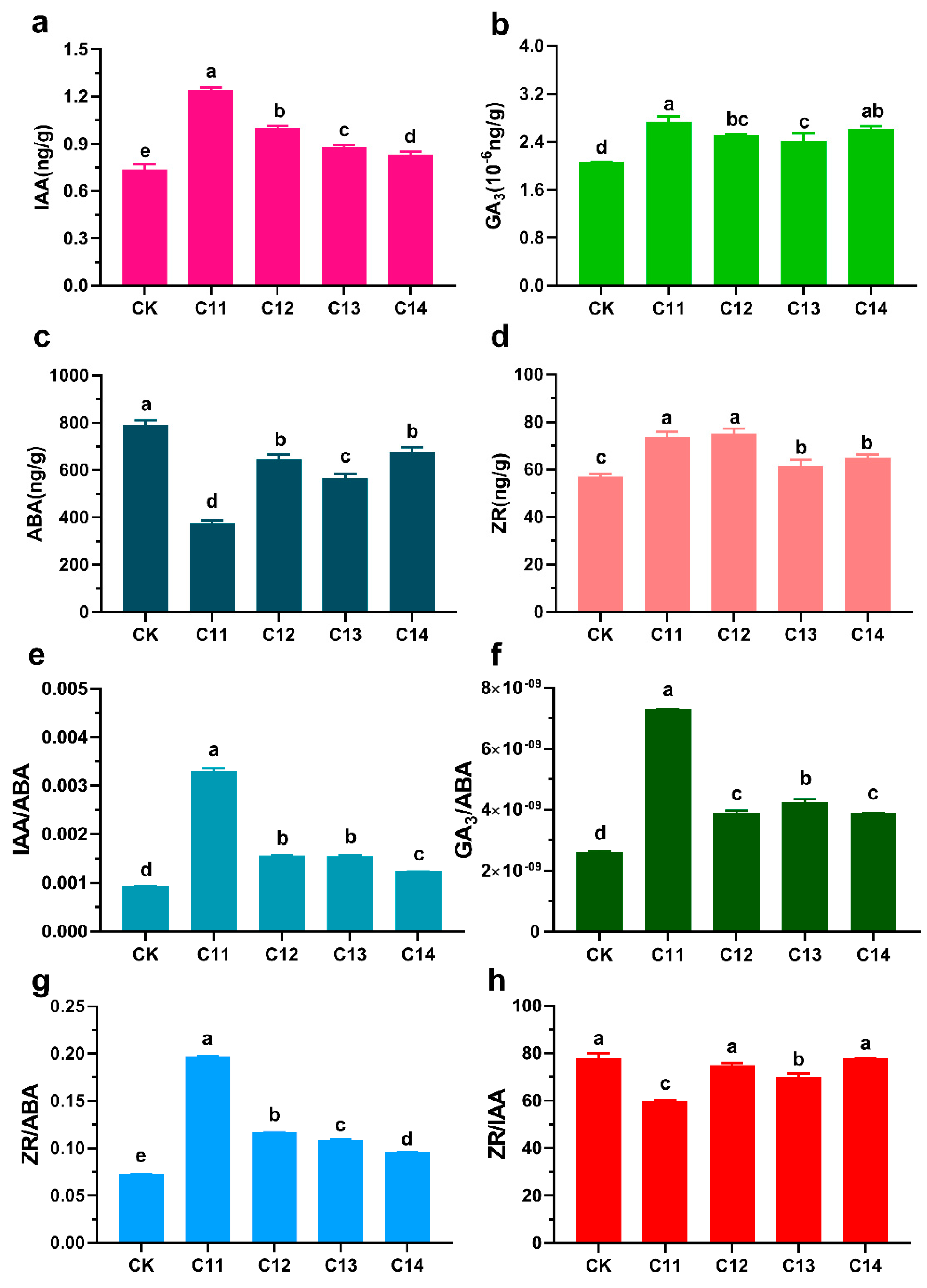
3.3. Effects of TDZ and 6-BA on Endogenous Hormone Levels in the Regeneration of F. mandshurica
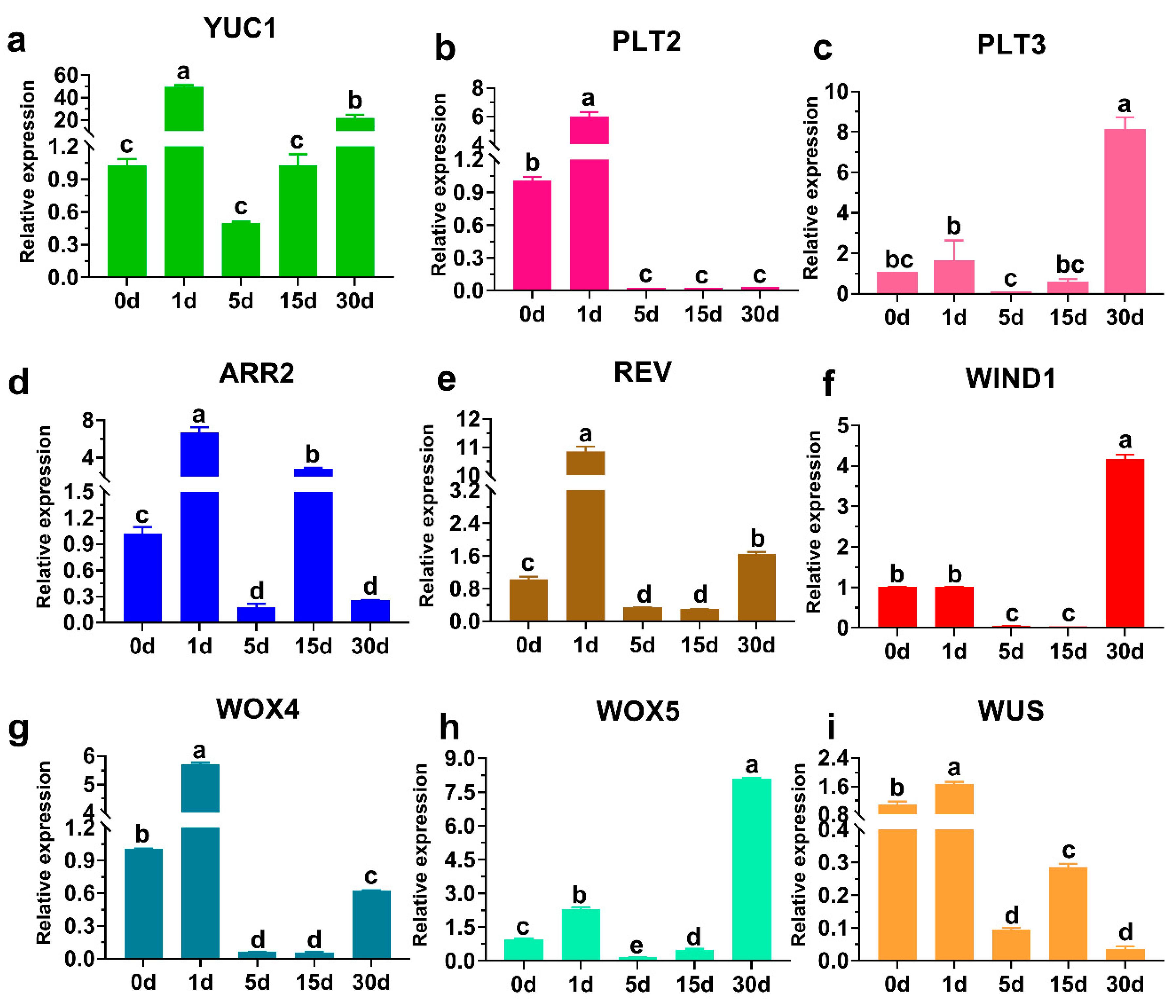
3.4. Analysis of Related Gene Expression Patterns during the Regeneration of F. mandshurica
3.5. Establishment of a Callus Regeneration System from Stem Segments of F. mandshurica
4. Discussion
4.1. Effect of the Exogenous Hormones TDZ and 6-BA on Induction of Adventitious Buds of the Callus
4.2. Wound Treatment Is Beneficial for Callus Re-Differentiation
4.3. 5-aza Promotes Callus Re-Differentiation
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gu, J.C.; Wei, X.; Wang, J.; Dong, X.Y.; Wang, Z.Q. Marked differences in standing biomass, length density, anatomy and physiological activity between white and brown roots in fraxinus mandshurica rupr. plantation. Plant Soil 2015, 392, 267–277. [Google Scholar] [CrossRef]
- Wang, J.S.; Wu, L.; Zhao, X.H.; Fan, J.; Zhang, C.Y.; von Gadow, K. Influence of ground flora on fraxinus mandshurica seedling growth on abandoned land and beneath forest canopy. Eur. J. For. Res. 2013, 132, 313–324. [Google Scholar] [CrossRef]
- Chen, X.H.; Wang, H.T.; Li, X.Y.; Ma, K.; Zhan, Y.G.; Zeng, F.S. Molecular cloning and functional analysis of 4-coumarate:Coa ligase 4(4cl-like 1)from fraxinus mandshurica and its role in abiotic stress tolerance and cell wall synthesis. BMC Plant Biol. 2019, 19, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.G.; Fan, X.H.; Wang, J.S.; Zhang, C.Y.; Lu, W.M.; Gadow, K.V. Spectral reflectance response of fraxinus mandshurica leaves to above- and belowground competition. Int. J. Remote Sens. 2012, 33, 5072–5086. [Google Scholar] [CrossRef]
- Sun, X.D.; Wang, W.N.; Razaq, M.; Sun, H.L. Effects of stand density on soil organic carbon storage in the top and deep soil layers of fraxinus mandshurica plantations. Austrian J. For. Sci. 2019, 136, 27–44. [Google Scholar]
- He, L.M.; Zeng, F.S.; He, Z.L.; Li, Y.H.; Li, S.J.; Zhan, Y.G. Interspecific hybridizations of fraxinus l. (f. Mandshurica x f. Americana and f. Mandshurica x f. Velutina) and heterosis analysis and selection of f1 progenies. Can. J. For. Res. 2019, 49, 1265–1276. [Google Scholar] [CrossRef]
- Kong, D.M.; Preece, J.E.; Shen, H.L. Somatic embryogenesis in immature cotyledons of manchurian ash (fraxinus mandshurica rupr.). Plant Cell Tissue Organ. Cult. 2012, 108, 485–492. [Google Scholar] [CrossRef]
- Stener, L.G. Genetic evaluation of damage caused by ash dieback with emphasis on selection stability over time. For. Ecol. Manag. 2018, 409, 584–592. [Google Scholar] [CrossRef]
- Inoue, T.; Okane, I.; Ishiga, Y.; Degawa, Y.; Hosoya, T.; Yamaoka, Y. The life cycle of hymenoscyphus fraxineus on manchurian ash, fraxinus mandshurica, in Japan. Mycoscience 2019, 60, 89–94. [Google Scholar] [CrossRef]
- Du, X.Z.; Wei, X. Definition of fine roots on the basis of the root anatomy, diameter, and branch orders of one-year old fraxinus mandshurica seedlings. J. For. Res. 2018, 29, 1321–1327. [Google Scholar] [CrossRef]
- Lee, J.H.; Pijut, P.M. Adventitious shoot regeneration from in vitro leaf explants of fraxinus nigra. Plant Cell Tissue Organ. Cult. 2017, 130, 335–343. [Google Scholar] [CrossRef]
- Aida, M.; Beis, D.; Heidstra, R.; Willemsen, V.; Blilou, I.; Galinha, C.; Nussaume, L.; Noh, Y.S.; Amasino, R.; Scheres, B. The plethora genes mediate patterning of the arabidopsis root stem cell niche. Cell 2004, 119, 109–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwase, A.; Mitsuda, N.; Koyama, T.; Hiratsu, K.; Kojima, M.; Arai, T.; Inoue, Y.; Seki, M.; Sakakibara, H.; Sugimoto, K.; et al. The ap2/erf transcription factor wind1 controls cell dedifferentiation in arabidopsis. Curr. Biol. 2011, 21, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Guo, J.S.; Feng, Z.H.; Cui, X.H.; Zhu, J. Molecular characterization of a novel ap2 transcription factor thwind1-l from thellungiella halophila. Plant Cell Tissue Organ. Cult. 2012, 110, 423–433. [Google Scholar] [CrossRef]
- Chen, L.Q.; Sun, B.B.; Xu, L.; Liu, W. Wound signaling: The missing link in plant regeneration. Plant Signal. Behav. 2016, 11, 4273–4284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, W.; Wang, Z.C.; Liang, Y.; Wang, Y.H.; Hu, Y.X. Dynamic expression reveals a two-step patterning of wus and clv3 during axillary shoot meristem formation in arabidopsis. J. Plant Physiol. 2017, 214, 1–6. [Google Scholar] [CrossRef]
- Kareem, A.; Durgaprasad, K.; Sugimoto, K.; Du, Y.J.; Pulianmackal, A.J.; Trivedi, Z.B.; Abhayadev, P.V.; Pinon, V.; Meyerowitz, E.M.; Scheres, B.; et al. Plethora genes control regeneration by a two-step mechanism. Curr. Biol. 2015, 25, 1017–1030. [Google Scholar] [CrossRef] [Green Version]
- Wojcikowska, B.; Botor, M.; Moronczyk, J.; Wojcik, A.M.; Nodzynski, T.; Karcz, J.; Gaj, M.D. Trichostatin a triggers an embryogenic transition in arabidopsis explants via an auxin-related pathway. Front. Plant Sci. 2018, 9, 1353. [Google Scholar] [CrossRef]
- Zhao, Q.; Du, Y.L.; Wang, H.T.; Rogers, H.J.; Yu, C.M.; Liu, W.; Zhao, M.Z.; Xie, F.T. 5-azacytidine promotes shoot regeneration during agrobacterium-mediated soybean transformation. Plant Physiol. Biochem. 2019, 141, 40–50. [Google Scholar] [CrossRef]
- Shemer, O.; Landau, U.; Candela, H.; Zemach, A.; Williams, L.E. Competency for shoot regeneration from arabidopsis root explants is regulated by DNA methylation. Plant Sci. 2015, 238, 251–261. [Google Scholar] [CrossRef]
- Liang, N.S.; Zhan, Y.G.; Yu, L.; Wang, Z.Q.; Zeng, F.S. Characteristics and expression analysis of fmtcp15 under abiotic stresses and hormones and interact with della protein in fraxinus mandshurica rupr. Forests 2019, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Liang, N.S.; Yu, L.; Liu, C.H.; Wang, Z.Q.; Zhao, X.T.; Zhan, Y.G. Molecular cloning and expression under abiotic stresses and hormones of the ethylene response factor vii gene fmrap2.12 from fraxinus mandshurica. J. For. Res. 2019, 30, 1289–1300. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yang, X.; Guo, T.; Gao, K.; Zhao, T.; Chen, Z.; An, X. High-efficiency somatic embryogenesis from seedlings of koelreuteria paniculata laxm. Forests 2018, 9, 769. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.Q.; Han, Z.Q.; Kang, X.Y. Adventitious shoot regeneration from leaf, petiole and root explants in triploid (populus albaxp.Glandulosa)xp.Tomentosa. Plant Cell Tissue Organ. Cult. 2019, 138, 121–130. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Shibata, M.; Rymen, B.; Iwase, A.; Bagman, A.M.; Watt, L.; Coleman, D.; Favero, D.S.; Takahashi, T.; Ahnert, S.E.; et al. A gene regulatory network for cellular reprogramming in plant regeneration. Plant Cell Physiol. 2018, 59, 770–782. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, S.G.; Lee, M.; Lee, I.; Park, H.Y.; Seo, P.J.; Jung, J.H.; Kwon, E.J.; Suh, S.W.; Paek, K.H.; et al. Hd-zip iii activity is modulated by competitive inhibitors via a feedback loop in arabidopsis shoot apical meristem development. Plant Cell 2008, 20, 920–933. [Google Scholar] [CrossRef] [Green Version]
- Shull, T.E.; Kurepa, J.; Smalle, J.A. Cytokinin signaling promotes differential stability of type-b arrs. Plant Signal. Behav. 2016, 11, e1169354. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, H.L.; Guo, H.R.; Xie, L.; Zeng, R.Z.; Zhang, X.Q.; Zhang, Z.S. Transcriptomic and hormonal analyses reveal that yuc-mediated auxin biogenesis is involved in shoot regeneration from rhizome in cymbidium. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Ohme-Takagi, M. Tcps, wuss, and winds: Families of transcription factors that regulate shoot meristem formation, stem cell maintenance, and somatic cell differentiation. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Iwase, A.; Mita, K.; Favero, D.S.; Mitsuda, N.; Sasaki, R.; Kobayshi, M.; Takebayashi, Y.; Kojima, M.; Kusano, M.; Oikawa, A.; et al. Wind1 induces dynamic metabolomic reprogramming during regeneration in brassica napus. Dev. Biol. 2018, 442, 40–52. [Google Scholar] [CrossRef]
- Orlowska, A.; Kepczynska, E. Identification of polycomb repressive complex1, trithorax group genes and their simultaneous expression with wuschel, wuschel-related homeobox5 and shoot meristemless during the induction phase of somatic embryogenesis in medicago truncatula gaertn. Plant Cell Tissue Organ. Cult. 2018, 134, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Iwase, A.; Mitsuda, N.; Ikeuchi, M.; Ohnuma, M.; Koizuka, C.; Kawamoto, K.; Imamura, J.; Ezura, H.; Sugimoto, K. Arabidopsis wind1 induces callus formation in rapeseed, tomato, and tobacco. Plant Signal. Behav. 2013, 8, e27432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwase, A.; Mita, K.; Nonaka, S.; Ikeuchi, M.; Koizuka, C.; Ohnuma, M.; Ezura, H.; Imamura, J.; Sugimoto, K. Wind1-based acquisition of regeneration competency in arabidopsis and rapeseed. J. Plant Res. 2015, 128, 389–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwase, A.; Harashima, H.; Ikeuchi, M.; Rymen, B.; Ohnuma, M.; Komaki, S.; Morohashi, K.; Kurata, T.; Nakata, M.; Ohme-Takagi, M.; et al. Wind1 promotes shoot regeneration through transcriptional activation of enhancer of shoot regeneration1 in arabidopsis. Plant Cell 2017, 29, 54–69. [Google Scholar] [CrossRef] [Green Version]
- Salaj, T.; Klubicova, K.; Matusova, R.; Salaj, J. Somatic embryogenesis in selected conifer trees pinus nigra arn. And abies hybrids. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Ree, J.F.; Polesi, L.G.; Back, F.; Bertolazi, A.A.; Silveira, V.; Guerra, M.P. Aging peach palm (bactris gasipaes kunth) cultures lose embryogenic potential and metabolic cellular function due to continuous culture in hypoxic environments. Plant Cell Tissue Organ. Cult. 2019, 140, 49–67. [Google Scholar] [CrossRef]
- Muniandy, K.; Tan, M.H.; Song, B.K.; Ayub, Q.; Rahman, S. Comparative sequence and methylation analysis of chloroplast and amyloplast genomes from rice. Plant Mol. Biol. 2019, 100, 33–46. [Google Scholar] [CrossRef]
- Li, J.Y.; Wang, M.J.; Li, Y.J.; Zhang, Q.H.; Lindsey, K.; Daniell, H.; Jin, S.X.; Zhang, X.L. Multi-omics analyses reveal epigenomics basis for cotton somatic embryogenesis through successive regeneration acclimation process. Plant Biotechnol. J. 2019, 17, 435–450. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.J.; Li, N.; Zhang, Z.B.; Meng, X.C.; Dong, Q.L.; Xu, C.M.; Gong, L.; Liu, B. Cg hypomethylation leads to complex changes in DNA methylation and transpositional burst of diverse transposable elements in callus cultures of rice. Plant J. 2019, 101, 188–203. [Google Scholar] [CrossRef]
- Gulyas, A.; Dobranszki, J.; Kiss, E.; da Silva, J.A.T.; Posta, K.; Hidvegi, N. Changes in DNA methylation pattern of apple long-term in vitro shoot culture and acclimatized plants. J. Plant Physiol. 2019, 239, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Osorio-Montalvo, P.; Saenz-Carbonell, L.; De-la-Pena, C. 5-azacytidine: A promoter of epigenetic changes in the quest to improve plant somatic embryogenesis. Int. J. Mol. Sci. 2018, 19, 3182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.J.; Ma, L.L.; Yang, X.R.; Zhang, L.; Zeng, X.; Xie, S.P.; Peng, H.W.; Gao, S.B.; Lin, H.J.; Pan, G.T.; et al. Integrative analysis of DNA methylation, mrnas, and small rnas during maize embryo dedifferentiation. BMC Plant Biol. 2017, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Media Number | Basic Medium Type | Sucrose (g/L) | Agar (g/L) | 6-BA (mg/L) | TDZ (mg/L) | IBA (mg/L) | Coconut (v/v) | TSA (mg/L) | 5-aza (mg/L) |
|---|---|---|---|---|---|---|---|---|---|
| c1 | MSB5 | 30 | 7 | 0 | 6 | 0.1 | 5% | 0 | 0 |
| c2 | MSB5 | 30 | 7 | 5 | 6 | 0.1 | 5% | 0 | 0 |
| c3 | MSB5 | 30 | 7 | 6 | 6 | 0.1 | 5% | 0 | 0 |
| c4 | MSB5 | 30 | 7 | 7 | 6 | 0.1 | 5% | 0 | 0 |
| c5 | MSB5 | 30 | 7 | 8 | 6 | 0.1 | 5% | 0 | 0 |
| c6 | MSB5 | 30 | 7 | 0 | 7 | 0.1 | 5% | 0 | 0 |
| c7 | MSB5 | 30 | 7 | 5 | 7 | 0.1 | 5% | 0 | 0 |
| c8 | MSB5 | 30 | 7 | 6 | 7 | 0.1 | 5% | 0 | 0 |
| c9 | MSB5 | 30 | 7 | 7 | 7 | 0.1 | 5% | 0 | 0 |
| c10 | MSB5 | 30 | 7 | 8 | 7 | 0.1 | 5% | 0 | 0 |
| c11 | MSB5 | 30 | 7 | 0 | 8 | 0.1 | 5% | 0 | 0 |
| c12 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 0 | 0 |
| c13 | MSB5 | 30 | 7 | 6 | 8 | 0.1 | 5% | 0 | 0 |
| c14 | MSB5 | 30 | 7 | 7 | 8 | 0.1 | 5% | 0 | 0 |
| c15 | MSB5 | 30 | 7 | 8 | 8 | 0.1 | 5% | 0 | 0 |
| b12 | MS | 30 | 7 | 5 | 8 | 0.1 | 5% | 0 | 0 |
| d12 | WPM | 30 | 7 | 5 | 8 | 0.1 | 5% | 0 | 0 |
| T5 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 0.15 | 0 |
| T6 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 0.3 | 0 |
| T7 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 0.6 | 0 |
| T8 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 1.2 | 0 |
| A5 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 0 | 2.4 |
| A6 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 0 | 6.1 |
| A7 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 0 | 12.21 |
| A8 | MSB5 | 30 | 7 | 5 | 8 | 0.1 | 5% | 0 | 24.42 |
| Primer Name | Primer Sequence |
|---|---|
| qFmTU-F | 5′-AGGACGCTGCCAACAACTTT-3′ |
| qFmTU-R | 5′-TTGAGGGGAAGGGTAAATAGTG-3′ |
| qFmAHP6-F | 5′-GTAAGTCACCACCAATGCCCA-3′ |
| qFmAHP6-R | 5′-TCAGAGTTACAGAGGGTGATGCC-3′ |
| qFmYUC1-F | 5′-CCAGCATCACCACCTTCAACAT-3′ |
| qFmYUC1-R | 5′-CGAACAGACCCTCGGCAAGTA-3′ |
| qFmPLT2-F | 5′-TGACACTCCGCTTCATAACCA-3′ |
| qFmPLT2-R | 5′-AACAGATAATCGGAACCACTCG-3′ |
| qFmPLT3-F | 5′-GGAACATTTGCTACTGAGGAGG-3′ |
| qFmPLT3-R | 5′-GAGATTATGATGATAGAATGGGGC-3′ |
| qFmARR1-F | 5′- GGGTTTGATGAGGAGAGTGGG -3′ |
| qFmARR1-R | 5′- GTCCCATTTAGTCCCCGTTTG -3′ |
| qFmARR2-F | 5′-CTATGCCTGGGATGACTGGA-3′ |
| qFmARR2-R | 5′-GGAAAGCCATGATCGTGAAA-3′ |
| qFmESR2-F | 5′-GGCTTGGAACTTTTGACACG-3′ |
| qFmESR2-R | 5′-TGAAGCTATACGAACGGACAAG-3′ |
| qFmPHV-F | 5′-CGGATAAGCAAAACTGGGGGT-3′ |
| qFmPHV-R | 5′-AGGGTGGTGGCTCCATTATTCA-3′ |
| qFmREV-F | 5′-CGGTTACTTTCGCCAGCATAC-3′ |
| qFmREV-R | 5′-GGCTTCATTCCAGGCATTTG-3′ |
| qFmWIND1-F | 5′-TGCCAATCACAGGATCTAAA-3′ |
| qFmWIND1-R | 5′-CTGACCAAACTGACACGAAA-3′ |
| qFmWOX4-F | 5′-TGGATTCTGGGAGCAACATA-3′ |
| qFmWOX4-R | 5′-AGCCGATTCCTTCTTGTGAT-3′ |
| qFmWOX5-F | 5′-GGCAATAATGGCGTCAGTGG-3′ |
| qFmWOX5-R | 5′-CCCTGAATGAGTTGAGTGGAAATA-3′ |
| qFmWUS-F | 5′- GGAGGAAGTCTTTGTGGTGC -3′ |
| qFmWUS-R | 5′- GAACACTTTGATTGGAGGTCG -3′ |
| qFmZPR3-F | 5′-GCAGTGCTTGATTCTCCTGGTT-3′ |
| qFmZPR3-R | 5′-ATGGAGAGGCTAAACTCAAAGCTG-3′ |
| Media Number | 6-BA (mg/L) | TDZ (mg/L) | Cotyledon Survival Rate (%) | Cotyledon Callus Induction Rate (%) | Root Survival Rate (%) | Root Callus Induction Rate (%) | Adventitious Bud Induction Rate (%) |
|---|---|---|---|---|---|---|---|
| a1 | 0 | 0 | 17.5 ± 5.46hi | 55.00 ± 7.07d | 25.42 ± 4.28jkl | 0g | 0 |
| a2 | 0 | 0.1 | 50 ± 1.57f | 0g | 28.48 ± 5.07ijk | 14.58 ± 7.92e | 0 |
| a3 | 0 | 0.5 | 59.52 ± 0.49ef | 41.67 ± 3.27e | 54.94 ± 1.42bcd | 23.33 ± 2.35d | 0 |
| a4 | 0 | 1 | 100a | 100a | 57.71 ± 2.96abc | 48.49 ± 1.44a | 0 |
| a5 | 1.5 | 0 | 12.5 ± 8.92i | 0g | 18.06 ± 0.39l | 0g | 0 |
| a6 | 1.5 | 0.1 | 88.89 ± 8.17b | 0g | 47.17 ± 4.59def | 10.42 ± 7.5ef | 0 |
| a7 | 1.5 | 0.5 | 55.56 ± 6.29f | 44.44 ± 1.02e | 51.39 ± 8.58cde | 33.68 ± 4.15c | 0 |
| a8 | 1.5 | 1 | 78.42 ± 3.75cd | 92.71 ± 3.15b | 61.86 ± 0.69ab | 27.42 ± 5.54cd | 0 |
| a9 | 2 | 0 | 27.27 ± 1.31gh | 18.18 ± 6.96f | 64.44 ± 6.81a | 0g | 0 |
| a10 | 2 | 0.1 | 57.14 ± 9.24f | 0g | 36.31 ± 2.2ghi | 4.17 ± 7.48fg | 0 |
| a11 | 2 | 0.5 | 75.25 ± 6.75d | 63.89 ± 8.46c | 48.33 ± 7.63def | 14.17 ± 2.68e | 0 |
| a12 | 2 | 1 | 87.5 ± 6.5bc | 91.67 ± 6.29b | 44.55 ± 6.8efg | 40.97 ± 0.02b | 0 |
| a13 | 3 | 0 | 50 ± 8.05f | 0g | 28.65 ± 3.5ijk | 0g | 0 |
| a14 | 3 | 0.1 | 20 ± 5.19hi | 12.5 ± 3.33f | 24.64 ± 0.83jkl | 0g | 0 |
| a15 | 3 | 0.5 | 10 ± 6.44i | 0g | 33.93 ± 0.16hij | 0g | 0 |
| a16 | 3 | 1 | 70.00 ± 3.57d | 70 ± 5.11c | 33.22 ± 0.06hij | 29.64 ± 0.85cd | 0 |
| a17 | 5 | 0 | 32.14 ± 8.89g | 0g | 20.35 ± 6.81kl | 0g | 0 |
| a18 | 5 | 0.1 | 33.18 ± 3.79g | 15.48 ± 4.83f | 40.24 ± 0.08fgh | 3.33 ± 9.79fg | 0 |
| a19 | 5 | 0.5 | 68.75 ± 7.25de | 48.75 ± 7.91de | 27.56 ± 8.77ijk | 10 ± 7.42ef | 0 |
| a20 | 5 | 1 | 50 ± 3.17f | 16.67 ± 4.99f | 58.47 ± 8.43abc | 50.71 ± 0.05a | 0 |
| Media Number | 6-BA (mg/L) | TDZ (mg/L) | Survival Rate (%) | Callus Induction Rate (%) | Adventitious Bud Induction Rate (%) |
|---|---|---|---|---|---|
| a1 | 0 | 0 | 26.39 ± 7.01de | 10 ± 1.2ij | 0c |
| a2 | 0 | 0.1 | 30.95 ± 3.58de | 25 ± 8.41gh | 0c |
| a3 | 0 | 0.5 | 82.59 ± 8.48b | 77.38 ± 4.81b | 0c |
| a4 | 0 | 1 | 96.43 ± 1.39a | 71.43 ± 9.92b | 3.13 ± 1.25b |
| a5 | 1.5 | 0 | 0f | 0j | 0c |
| a6 | 1.5 | 0.1 | 25 ± 6.83e | 25 ± 6.36gh | 0c |
| a7 | 1.5 | 0.5 | 58.33 ± 3.05c | 53.33 ± 3.1cd | 0c |
| a8 | 1.5 | 1 | 73.86 ± 7.51b | 87.5 ± 6.24a | 0c |
| a9 | 2 | 0 | 0f | 0j | 0c |
| a10 | 2 | 0.1 | 36.36 ± 6.73d | 18.18 ± 4.42hi | 0c |
| a11 | 2 | 0.5 | 78.47 ± 9.12b | 72.69 ± 2.8b | 0c |
| a12 | 2 | 1 | 95.83 ± 1.77a | 93.56 ± 2.39a | 0c |
| a13 | 3 | 0 | 28.57 ± 0.72de | 37.5 ± 4.71ef | 0c |
| a14 | 3 | 0.1 | 52.08 ± 9.97c | 31.94 ± 1.81fg | 0c |
| a15 | 3 | 0.5 | 30.16 ± 9.87de | 0j | 0c |
| a16 | 3 | 1 | 62.5 ± 2.03c | 59.29 ± 0.84c | 0c |
| a17 | 5 | 0 | 30 ± 7.14de | 20 ± 7.48h | 0c |
| a18 | 5 | 0.1 | 52.05 ± 2.08c | 27.56 ± 7.24gh | 0c |
| a19 | 5 | 0.5 | 78.61 ± 3.54b | 61.67 ± 9.79c | 0c |
| a20 | 5 | 1 | 81.75 ± 5.06b | 88.69 ± 9.64a | 10.52 ± 2.12a |
| a21 | 5 | 2 | 28.64 ± 6.22c | 46.35 ± 1.67de | 0c |
| a22 | 5 | 2 | 60.34 ± 7.93c | 40.72 ± 8.65ef | 0c |
| Media Number | 6-BA (mg/L) | TDZ (mg/L) | Survival Rate (%) | Callus Induction Rate (%) | Adventitious Bud Induction Rate (%) |
|---|---|---|---|---|---|
| c1 | 0 | 6 | 35.75 ± 1.02f | 67.98 ± 0.33k | 0g |
| c2 | 5 | 6 | 65.74 ± 3.08cd | 87.45 ± 0.22h | 0g |
| c3 | 6 | 6 | 78.68 ± 6.79a | 90.41 ± 0.07f | 0g |
| c4 | 7 | 6 | 67.48 ± 0.44bcd | 91.34 ± 0.45e | 4.55 ± 0.15ef |
| c5 | 8 | 6 | 79.64 ± 4.3a | 96.54 ± 0.7c | 3.33 ± 1.23f |
| c6 | 0 | 7 | 45.67 ± 6.27ef | 74.12 ± 0.84j | 6.25 ± 2.14de |
| c7 | 5 | 7 | 78.14 ± 7.54ab | 98.54 ± 0.41a | 3.7 ± 1.33f |
| c8 | 6 | 7 | 77.64 ± 5.69ab | 97.35 ± 0.09b | 0g |
| c9 | 7 | 7 | 64.12 ± 4cd | 88.97 ± 0.85g | 11.81 ± 1.49c |
| c10 | 8 | 7 | 61.47 ± 1.59d | 98.74 ± 0.37a | 16.21 ± 1.19b |
| c11 | 0 | 8 | 41.24 ± 8.43ef | 75.64 ± 0.03i | 6.94 ± 1.5d |
| c12 | 5 | 8 | 74.65 ± 8.32abc | 99.15 ± 0.27a | 33.33 ± 1.17a |
| c13 | 6 | 8 | 74.51 ± 3.69abc | 97.64 ± 0.36b | 10.48 ± 1.45c |
| c14 | 7 | 8 | 66.66 ± 9.88cd | 98.42 ± 0.03a | 2.86 ± 1.6f |
| c15 | 8 | 8 | 50.79 ± 5.99e | 95.11 ± 0.36d | 9.72 ± 1.69c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Li, X.; Tian, H.; Liu, H.; Xiao, Y.; Liang, N.; Zhao, X.; Zhan, Y. Effects of Hormones and Epigenetic Regulation on the Callus and Adventitious Bud Induction of Fraxinus mandshurica Rupr. Forests 2020, 11, 590. https://doi.org/10.3390/f11050590
Yu L, Li X, Tian H, Liu H, Xiao Y, Liang N, Zhao X, Zhan Y. Effects of Hormones and Epigenetic Regulation on the Callus and Adventitious Bud Induction of Fraxinus mandshurica Rupr. Forests. 2020; 11(5):590. https://doi.org/10.3390/f11050590
Chicago/Turabian StyleYu, Lei, Xinyu Li, Hongmei Tian, Hualing Liu, Ying Xiao, Nansong Liang, Xingtang Zhao, and Yaguang Zhan. 2020. "Effects of Hormones and Epigenetic Regulation on the Callus and Adventitious Bud Induction of Fraxinus mandshurica Rupr." Forests 11, no. 5: 590. https://doi.org/10.3390/f11050590