Microsite Influence on Woody Plant Regeneration in a Pinus palustris Woodland Following Catastrophic Disturbance


Abstract
:1. Introduction
2. Materials and Methods
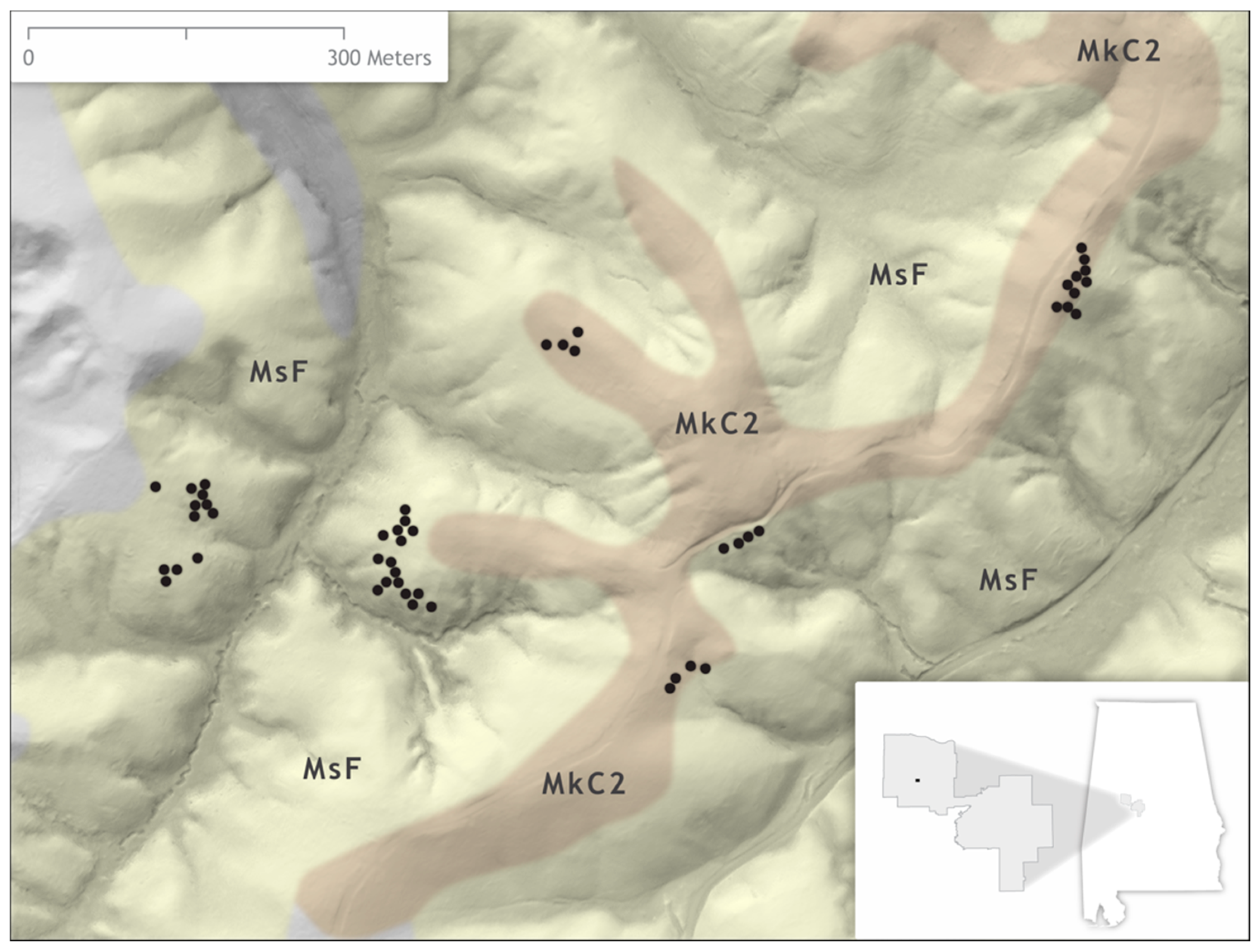
2.1. Study Site
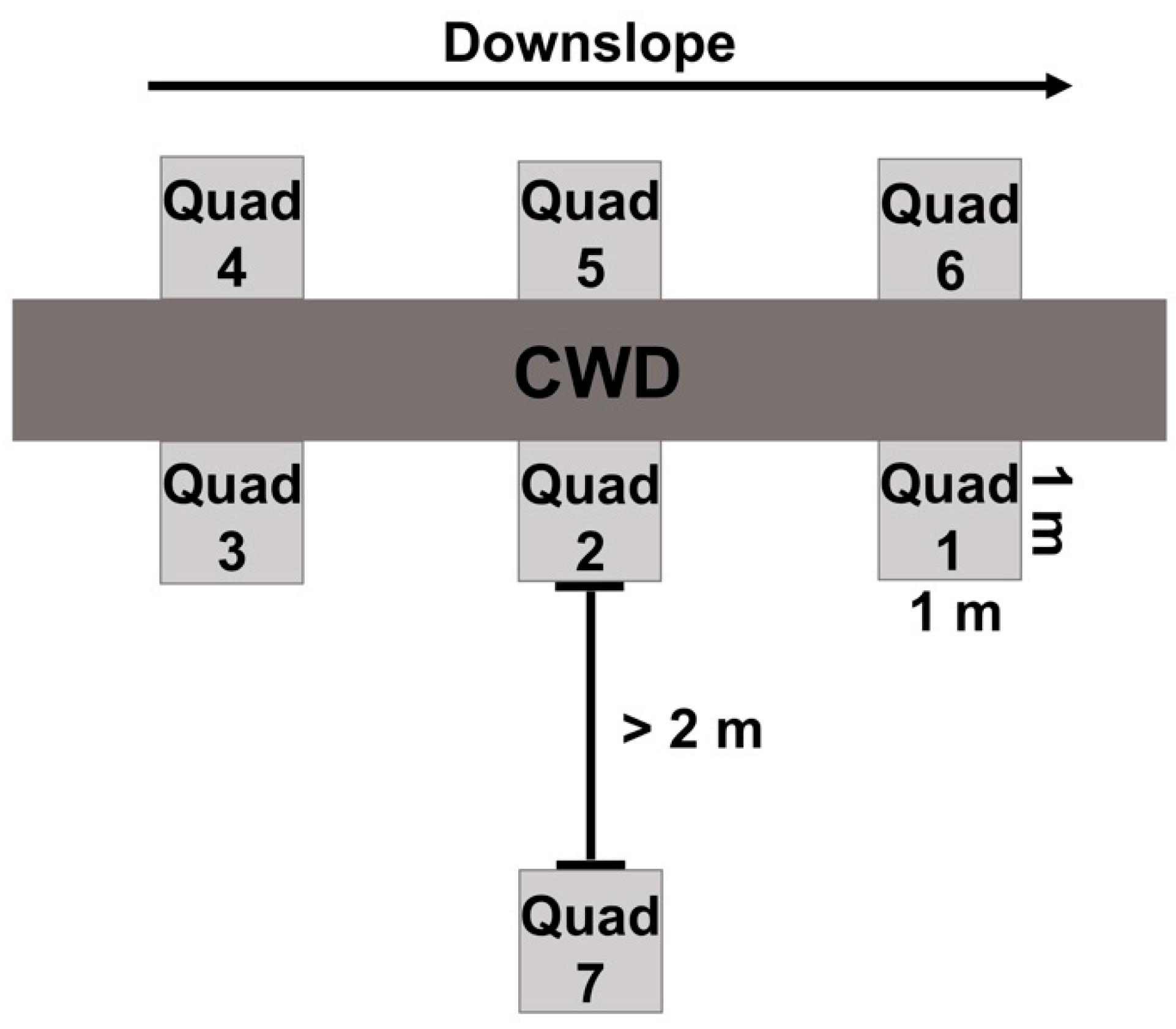
2.2. Field Methods
2.3. Laboratory and Analytical Methods
3. Results
3.1. Relative Density of Woody Plant Species
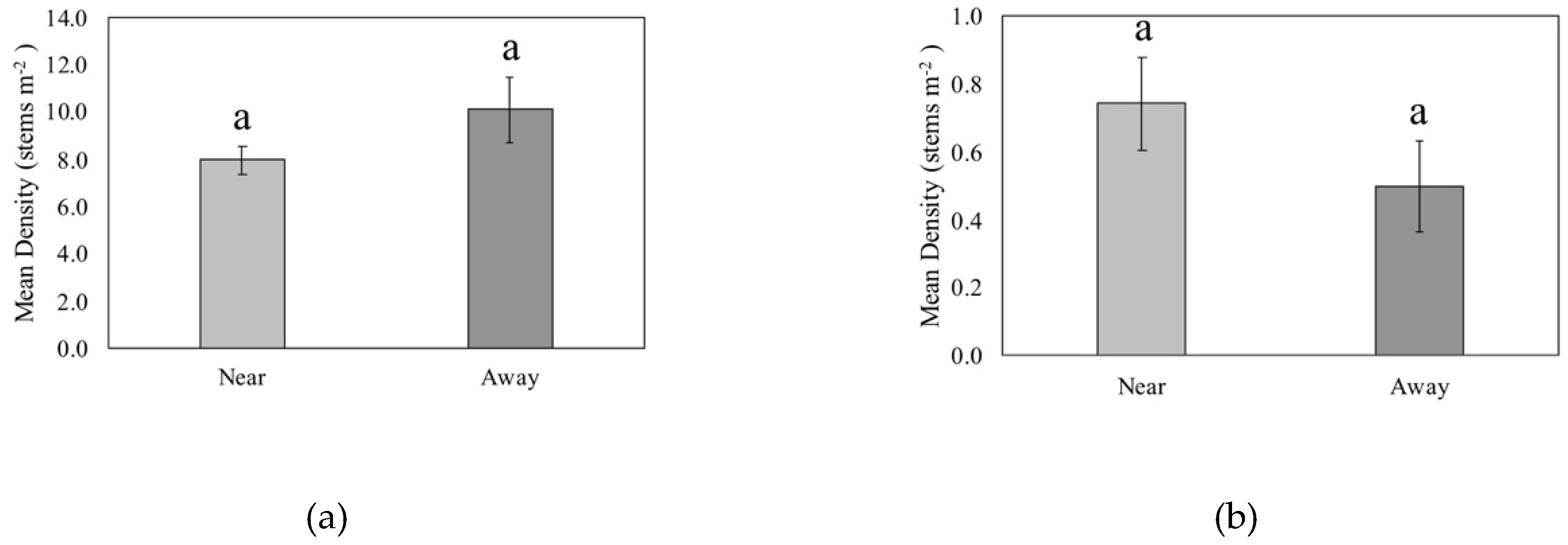
3.2. CWD Proximity and Woody Plant Density
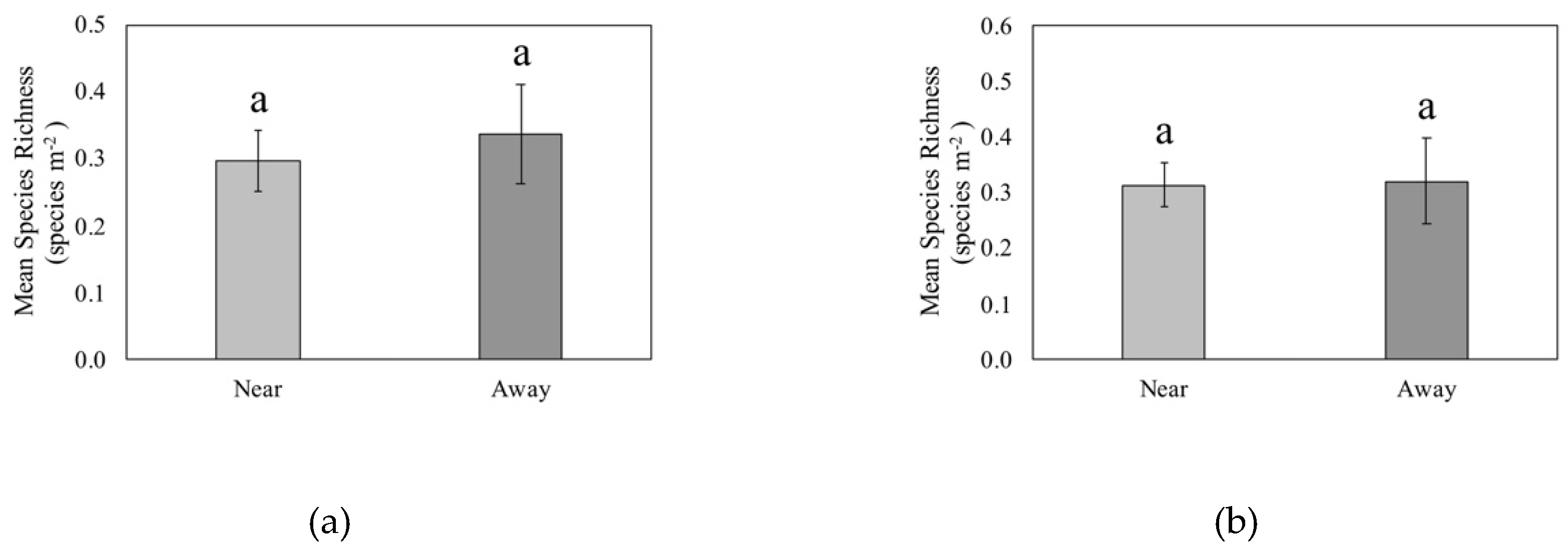
3.3. Proximity to CWD and Woody Plant Richness
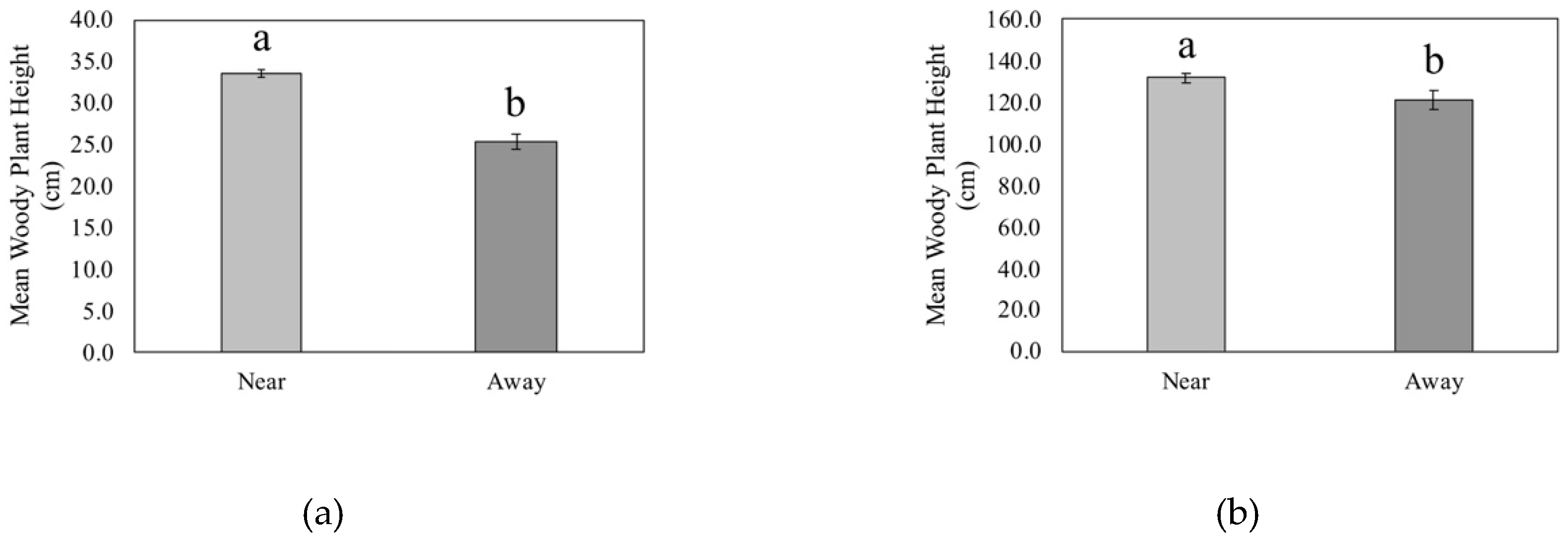
3.4. Proximity to CWD and Woody Plant Height
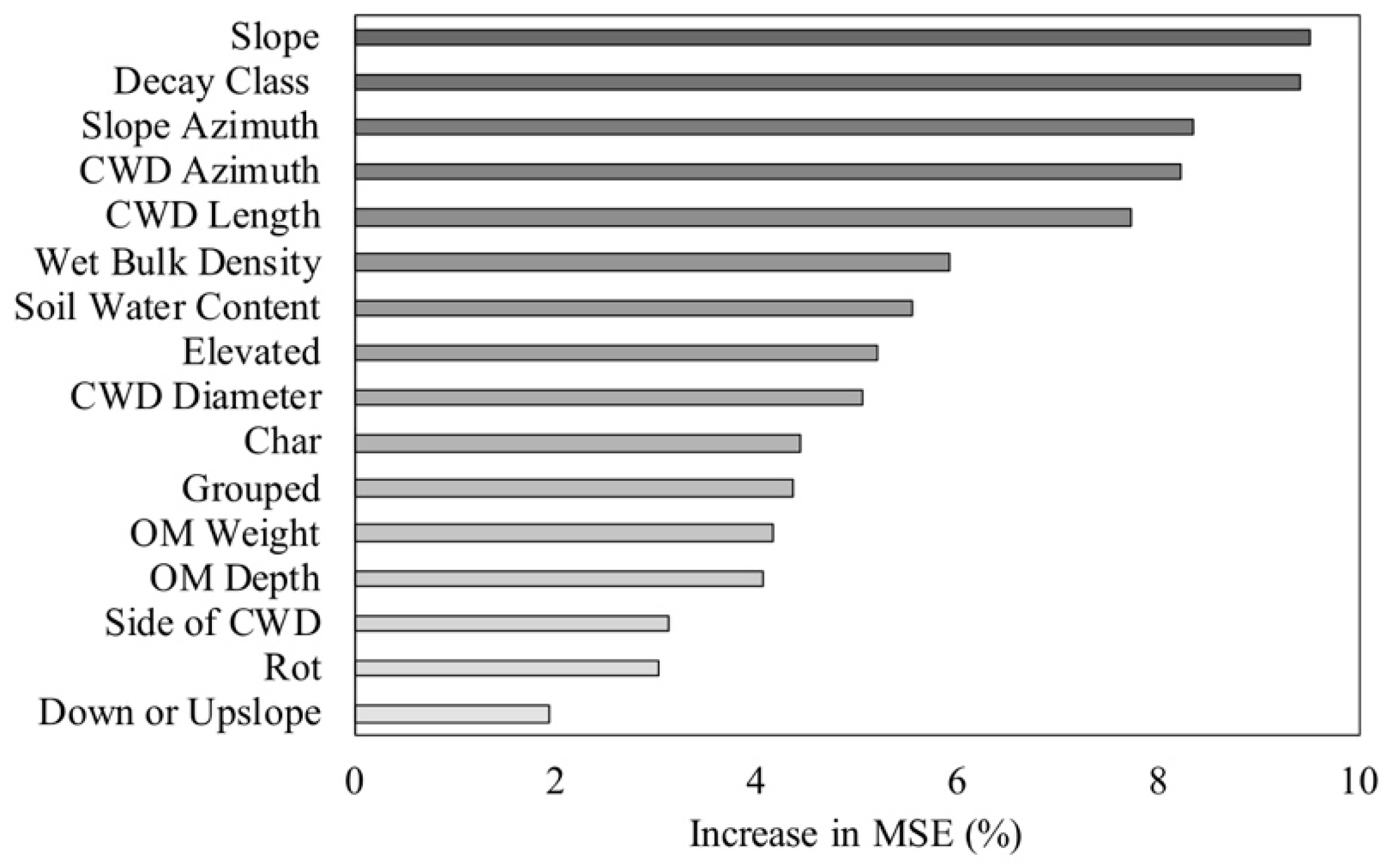
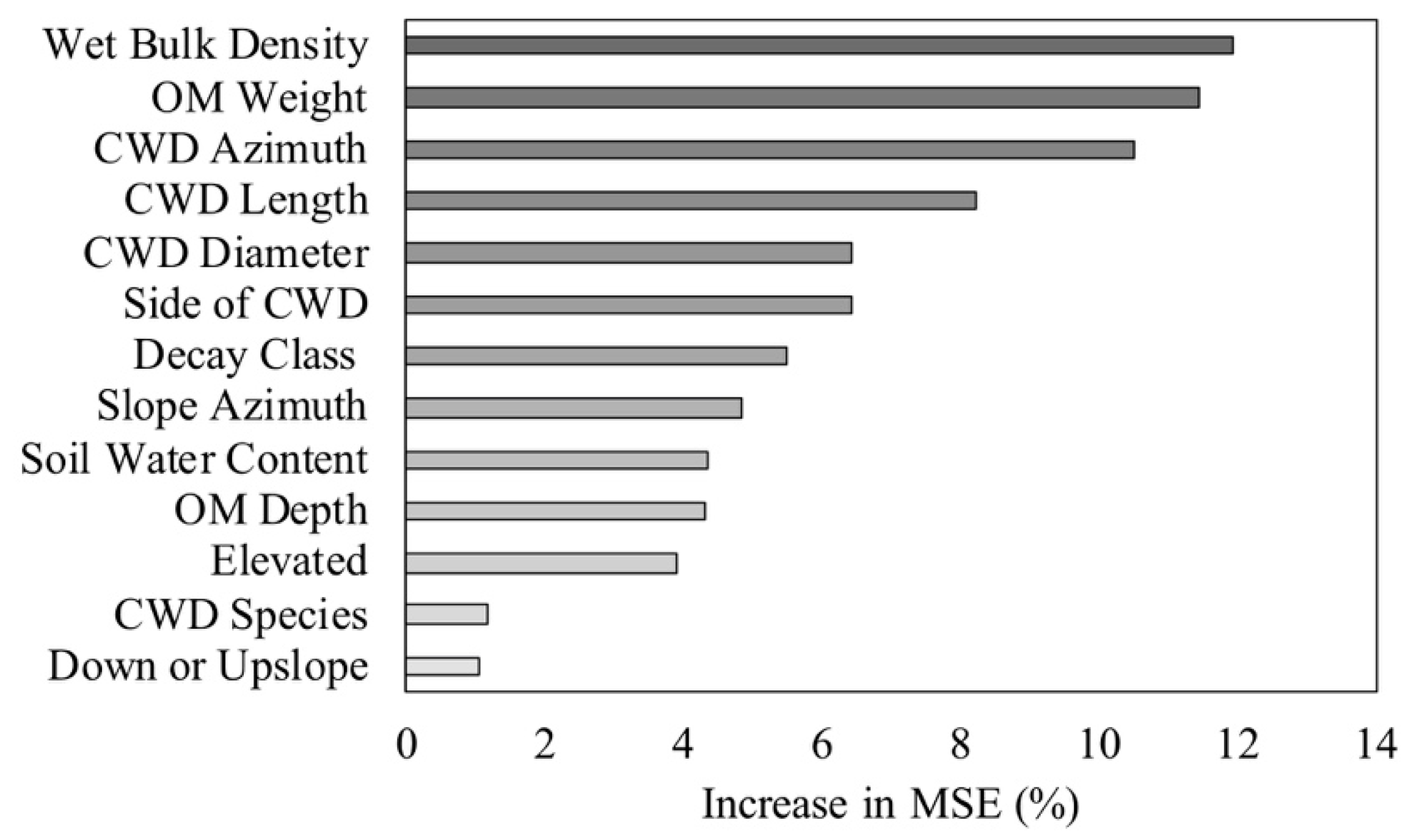
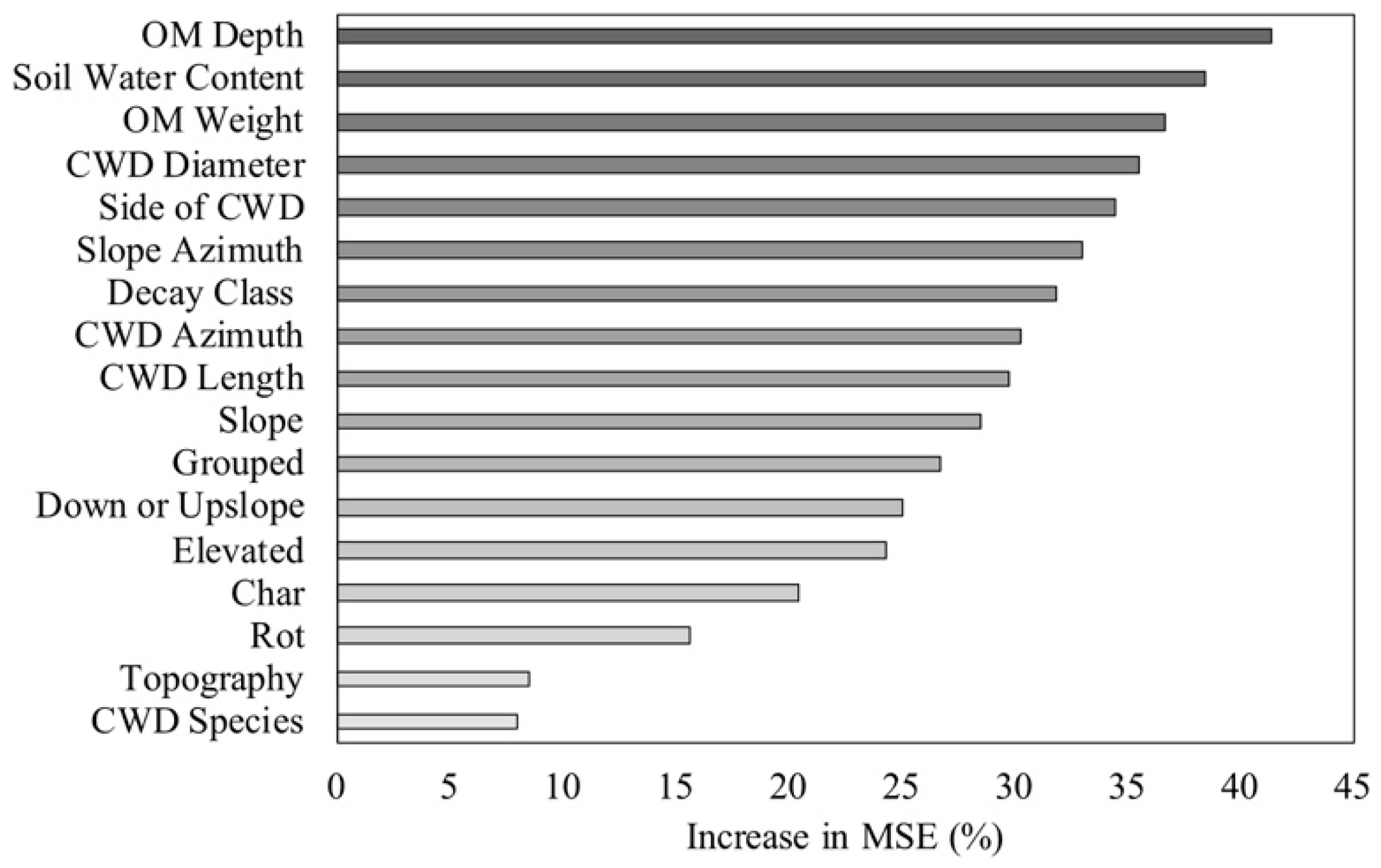
3.5. Influence of Microenvironmental Conditions on Woody Plant Characteristics
4. Discussion
4.1. Relative Density of Woody Plant Species
4.2. Proximity to CWD and Woody Plant Density, Richness, and Height
4.3. Influence of Microenvironmental Conditions on Woody Plant Characteristics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley and Sons: Hoboken, NJ, USA, 1996. [Google Scholar]
- Foster, D.R.; Knight, D.H.; Franklin, J.F. Landscape patterns and legacies resulting from large, infrequent forest disturbances. Ecosystems 1998, 1, 497–510. [Google Scholar] [CrossRef]
- Hart, J.L.; Kleinman, J.S. What are intermediate-severity forest disturbances and why are they important? Forests 2018, 9, 579. [Google Scholar] [CrossRef] [Green Version]
- Clements, F.E. Plant Succession: An Analysis of the Development of Vegetation; No. 242; Carnegie Institution of Washington Publication: Washington, DC, USA, 1916; pp. 1–512. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Loehle, C. Strategy space and the disturbance spectrum: A life-history model for tree species coexistence. Am. Nat. 2000, 156, 14–33. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Bush, M.B.; Richards, K.J.E.M. Plant recolonization and vegetation succession on the Krakatau Islands, Indonesia. Ecol. Monogr. 1989, 59, 59–123. [Google Scholar] [CrossRef]
- Webster, C.R.; Dickinson, Y.L.; Burton, J.I.; Frelich, L.E.; Jenkins, M.A.; Kern, C.C.; Raymond, P.; Saunders, M.R.; Walters, M.B.; Willis, J.L. Promoting and maintaining diversity in contemporary hardwood forests: Confronting contemporary drivers of change and the loss of ecological memory. For. Ecol. Manag. 2018, 421, 98–108. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Franklin, J.F.; Lindenmayer, D.; MacMahon, J.A.; McKee, A.; Magnuson, J.; Perry, D.A.; Waide, R.; Foster, D. Threads of continuity. Conserv. Biol. 2000, 1, 8–16. [Google Scholar] [CrossRef]
- Franklin, J.F.; Spies, T.A.; Van Pelt, R.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C.; et al. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The forgotten stage of forest succession: Early-successional ecosystems on forest sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.G.; Baker, W.L.; Peterson, C.J.; Peet, R.K. Factors influencing succession: Lessons from large, infrequent natural disturbances. Ecosystems 1998, 1, 511–523. [Google Scholar] [CrossRef]
- Franklin, J.F.; MacMahon, J.A. Messages from a mountain. Science 2000, 288, 1183–1184. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.G.; Davidson, N.J.; Close, D.C. Understanding the regeneration niche: Microsite attributes and recruitment of eucalypts in dry forests. For. Ecol. Manag. 2012, 269, 229–238. [Google Scholar] [CrossRef]
- Baskin, C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Henry, J.D.; Swan, J.M.A. Reconstructing Forest History from Live and Dead Plant Material—An Approach to the Study of Forest Succession in Southwest New Hampshire. Ecology 1974, 55, 772–783. [Google Scholar] [CrossRef]
- Lorimer, C.G. Age structure and disturbance history of a southern Appalachian virgin forest. Ecology 1980, 61, 1169–1184. [Google Scholar] [CrossRef]
- Zielonka, T. When does dead wood turn into a substrate for spruce replacement? J. Veg. Sci. 2006, 17, 739–746. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Pulliam, W.M.; Lodge, D.J.; Quinones-Orfila, V.; Fetcher, N.; Guzman-Grajales, S.; Parrotta, J.A.; Asbury, C.E.; Walker, L.R.; Waide, R.B. Nitrogen immobilization by decomposing woody debris and the recovery of tropical wet forest from hurricane damage. Oikos 1995, 72, 314–322. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F. Tree seedlings on logs in Picea-Tsuga forests of Oregon and Washington. Ecology 1989, 70, 48–59. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Miyamaoto, T. Racodium snow blight in Japan. For. Pathol. 2005, 35, 1–7. [Google Scholar] [CrossRef]
- Whyte, H.D.; Lusk, C.H. Woody debris in treefall gaps shelters palatable plant species from deer browsing, in an old-growth temperate forest. For. Ecol. Manag. 2019, 448, 198–207. [Google Scholar] [CrossRef]
- Waldron, K.; Ruel, J.C.; Gauthier, S. Forest structural attributes after windthrow and consequences of salvage logging. For. Ecol. Manag. 2013, 289, 28–37. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. In Advances in Ecological Research; MacFadyn, A., Ford, E.D., Eds.; Academic Press: Orlando, FL, USA, 1986; Volume 15, pp. 133–302. [Google Scholar]
- Cornett, M.W.; Reich, P.B.; Puettmann, K.J.; Frelich, L.E. Seedbed and moisture availability determine safe sites for early Thuja occidentalis (Cupressaceae) regeneration. Am. J. Bot. 2000, 87, 1807–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskell, D.E.; Flaspohler, D.J.; Webster, C.R.; Meyer, M.W. Variation in soil temperature, moisture, and plant growth with the addition of downed woody material on lakeshore restoration sites. Restor. Ecol. 2012, 20, 113–121. [Google Scholar] [CrossRef]
- Stroheker, S.; Weiss, M.; Sieber, T.N.; Bugmann, H. Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest. Forests 2018, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Frost, C. History and future of the longleaf pine ecosystem. In The Longleaf Pine Ecosystem; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer: New York, NY, USA, 2007; pp. 9–48. [Google Scholar] [CrossRef]
- Kirkman, L.K.; Coffey, K.L.; Mitchell, R.J.; Moser, E.B. Ground cover recovery patterns and life-history traits: Implications for restoration obstacles and opportunities in a species-rich savanna. J. Ecol. 2004, 92, 409–421. [Google Scholar] [CrossRef]
- Noss, R.F.; LaRoe, E.T.; Scott, J.M. Endangered Ecosystems of the United States: A Preliminary Assessment of Loss and Degradation; U.S. Department of the Interior, National Biological Service: Washington, DC, USA, 1995.
- Dirzo, R.; Raven, P.H. Global state of biodiversity and loss. Annu. Rev. Env. Resour. 2003, 28, 137–167. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.C.; Stehman, S.V.; Potapov, P.V.; Loveland, T.R.; Townshend, J.R.; De Fries, R.S.; Pittman, K.W.; Arunarwati, B.; Stolle, F.; Steininger, M.K.; et al. Humid tropical forest clearing from 2000 to 2005 quantified by using multitemporal and multiresolution remotely sensed data. Proc. Natl. Acad. Sci. USA 2008, 105, 9439–9444. [Google Scholar] [CrossRef] [Green Version]
- Seibold, S.; Bässler, C.; Brandl, R.; Gossner, M.M.; Thorn, S.; Ulyshen, M.D.; Müller, J. Experimental studies of dead-wood biodiversity—A review identifying global gaps in knowledge. Biol. Conserv. 2015, 191, 139–149. [Google Scholar] [CrossRef]
- Gensini, V.A.; Brooks, H.E. Spatial trends in United States tornado frequency. NPJ Clim. Atmos. Sci. 2018, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Brais, S.; Paré, D.; Lierman, C. Tree bole mineralization rates of four species of the Canadian eastern boreal forest: Implications for nutrient dynamics following stand- replacing disturbances. Can. J. For. Res. 2006, 36, 2331–2340. [Google Scholar] [CrossRef]
- Pinno, B.; Das Gupta, S. Coarse Woody Debris as a Land Reclamation Amendment at an Oil Sands Mining Operation in Boreal Alberta, Canada. Sustainability 2018, 10, 1640. [Google Scholar] [CrossRef] [Green Version]
- White, P.J.; McGill, B.J.; Lechowicz, M.J. Detecting changes in forest floor habitat after canopy disturbance. Ecol. Res. 2012, 27, 397–406. [Google Scholar] [CrossRef]
- Han, A.R.; Kim, H.J.; Jung, J.B.; Park, P.S. Seed germination and initial seedling survival of the subalpine tree species, Picea jezoensis, on different forest floor substrates under elevated temperature. For. Ecol. Manag. 2018, 429, 579–588. [Google Scholar] [CrossRef]
- Vítková, L.; Bače, R.; Kjučukov, P.; Svoboda, M. Deadwood management in Central European forests: Key considerations for practical implementation. For. Ecol. Manag. 2018, 429, 394–405. [Google Scholar] [CrossRef]
- Fenneman, N.M. Physiography of Eastern United States; McGraw-Hill: New York, NY, USA, 1938. [Google Scholar]
- Shankman, D.; Hart, J.L. The fall line: A physiographic forest-vegetation boundary. Geogr. Rev. 2007, 97, 502–519. [Google Scholar] [CrossRef]
- USDA NRCS (United States Department of Agriculture, Natural Resources Conservation Service). Web Soil Survey. Available online: https://websoilsurvey.nrcs.usda.gov/ (accessed on 23 April 2020).
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- PRISM Climate Group. Northwest Alliance for Computational Science and Engineering. Available online: http://www.prism.oregonstate.edu/ (accessed on 23 April 2020).
- USDA NOAA (United States Department of Agriculture, National Oceanic and Atmospheric Administration). United States Drought Monitor. Available online: https://data.nal.usda.gov/dataset/united-states-drought-monitor (accessed on 15 May 2020).
- Harper, R.M. Forests of Alabama Geological Survey of Alabama; Monograph 10; Wetumpka Printing Company: Wetumpka, AL, USA, 1943. [Google Scholar]
- Braun, E.L. Deciduous Forests of Eastern North America; The Blackburn Press: Caldwell, NJ, USA, 1950. [Google Scholar]
- Beckett, S.; Golden, M.S. Forest vegetation and vascular flora of Reed Brake Research Natural Area, Alabama. Castanea 1982, 47, 368–392. [Google Scholar]
- Cox, L.E.; Hart, J.L. Two centuries of forest compositional and structural changes in the Alabama Fall Line Hills. Am. Midl. Nat. 2015, 174, 218–238. [Google Scholar] [CrossRef]
- Kleinman, J.S.; Hart, J.L. Vascular flora of longleaf pine woodlands after wind disturbance and salvage harvesting in the Alabama Fall Line Hills. Castanea 2018, 83, 183–195. [Google Scholar] [CrossRef]
- Walker, J.L.; Silletti, A.M. Restoring the ground layer of longleaf pine ecosystems. In The Longleaf Pine Ecosystem; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer: New York, NY, USA, 2007; pp. 297–333. [Google Scholar] [CrossRef]
- Van Lear, D.H.; Carroll, W.D.; Kapeluck, P.R.; Johnson, R. History and restoration of the longleaf pine-grassland ecosystem: Implications for species at risk. For. Ecol. Manag. 2005, 211, 150–165. [Google Scholar] [CrossRef]
- Platt, W.J.; Evans, G.W.; Rathbun, S.L. The population dynamics of a long-lived conifer (Pinus palustris). Am. Nat. 1988, 131, 491–525. [Google Scholar] [CrossRef]
- Reed, F.W. A Working Plan for Forest Lands in Central Alabama; US Department of Agriculture, Forest Service: Washington, DC, USA, 1905.
- NCDC (National Climatic Data Center). Storm Events Database. Available online: https://www.ncdc.noaa.gov/stormevents/ (accessed on 25 April 2020).
- NWS (National Weather Service). Sawyerville-Eoline (Greene, Hale and Bibb Counties) EF-3 Tornado 27 April 2011. Available online: https://www.weather.gov/bmx/event_04272011sawyerville (accessed on 25 April 2020).
- Goode, J.D.; Kleinman, J.S.; Hart, J.L.; Bhuta, A.A. Edge influence on composition and structure of a Pinus palustris woodland following catastrophic wind disturbance. Can. J. For. Res. 2020, 50, 332–341. [Google Scholar] [CrossRef]
- Chećko, E.; Jaroszewicz, B.; Olejniczak, K.; Kwiatkowska-Falińska, A.J. The importance of coarse woody debris for vascular plants in temperate mixed deciduous forests. Can. J. For. Res. 2015, 45, 1154–1163. [Google Scholar] [CrossRef]
- Donato, D.C.; Campbell, J.L.; Fontaine, J.B.; Law, B.E. Quantifying char in postfire woody detritus inventories. Fire Ecol. 2009, 5, 104–115. [Google Scholar] [CrossRef]
- Knapp, E.E.; Keeley, J.E.; Ballenger, E.A.; Brennan, T.J. Fuel reduction and coarse woody debris dynamics with early season and late season prescribed fire in a Sierra Nevada mixed conifer forest. For. Ecol. Manag. 2005, 208, 383–397. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Horn, S.; Pokswinski, S.; McHugh, J.V.; Hiers, J.K. A comparison of coarse woody debris volume and variety between old-growth and secondary longleaf pine forests in the southeastern United States. For. Ecol. Manag. 2018, 429, 124–132. [Google Scholar] [CrossRef]
- Schowalter, T.D.; Zhang, Y.L.; Sabin, T.E. Decomposition and nutrient dynamics of oak Quercus spp. logs after five years of decomposition. Ecography 1998, 21, 3–10. [Google Scholar] [CrossRef]
- LaFevor, M.C. Restoration of degraded agricultural terraces: Rebuilding landscape structure and process. J. Environ. Manag. 2014, 138, 32–42. [Google Scholar] [CrossRef]
- Dorau, K.; Mansfeldt, T. Comparison of redox potential dynamics in a diked marsh soil: 1990 to 1993 versus 2011 to 2014. J. Plant Nutr. Soil Sci. 2016, 179, 641–651. [Google Scholar] [CrossRef]
- Tome, J.B., Jr.; Dechen, A.R.; Atkinson, R.J. Effects of moist storage and different drying temperatures on the extractability of iron, copper, manganese, and zinc in soil samples. Commun. Soil Sci. Plan. 1996, 27, 2591–2611. [Google Scholar] [CrossRef]
- Davies, G.M.; Gray, A. Don’t let spurious accusations of pseudoreplication limit our ability to learn from natural experiments (and other messy kinds of ecological monitoring). Ecol. Evol. 2015, 5, 5295–5304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org/ (accessed on 25 April 2020).
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Cutler, D.R.; Edwards, T.C., Jr.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Liaw, A.; Wiener, M. Classification and Regression by random Forest. R News 2002, 2, 18–22. [Google Scholar]
- Kleinman, J.S.; Goode, J.D.; Hart, J.L.; Dey, D.C. Prescribed fire effects on Pinus palustris woodland development after catastrophic wind disturbance and salvage logging. For. Ecol. Manag. 2020, in press. [Google Scholar] [CrossRef]
- Clendenin, M.; Ross, W.G. Effects of cool season prescribed fire on understory vegetation in a mixed pine hardwood forest of east Texas. Tex. J. Sci. 2001, 53, 65–78. [Google Scholar]
- Dey, D.C.; Hartman, G. Returning fire to Ozark Highland forest ecosystems: Effects on advance regeneration. For. Ecol. Manag. 2005, 217, 37–53. [Google Scholar] [CrossRef]
- Fan, Z.; Ma, Z.; Dey, D.C.; Roberts, S.D. Response of advance reproduction of oaks and associated species to repeated prescribed fires in upland oak-hickory forests, Missouri. For. Ecol. Manag. 2012, 266, 160–169. [Google Scholar] [CrossRef]
- Olson, D.F.; Boyce, S.G. Factors affecting acorn production and germination and early growth of seedlings and seedling sprouts. In Oak Symposium Proceedings; US Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Upper Darby, PA, USA, 1971; pp. 44–48. [Google Scholar]
- Whelan, A.W.; Bigelow, S.W.; Nieminen, M.F.; Jack, S.B. Fire season, overstory density and groundcover composition affect understory hardwood sprout demography in longleaf pine woodlands. Forests 2018, 9, 423. [Google Scholar] [CrossRef] [Green Version]
- Kuddes-Fischer, L.M.; Arthur, M.A. Response of understory vegetation and tree regeneration to a single prescribed fire in oak-pine forests. Nat. Area J. 2002, 22, 43–52. [Google Scholar]
- Kleinman, J.S.; Ford, S.A.; Hart, J.L. Catastrophic wind and salvage harvesting effects on woodland plants. For. Ecol. Manag. 2017, 403, 112–125. [Google Scholar] [CrossRef]
- Lyrene, P.M. First report of Vaccinium arboreum hybrids with cultivated highbush blueberry. HortScience 2011, 46, 563–566. [Google Scholar] [CrossRef] [Green Version]
- Freeman, D.C.; Brown, M.L.; Duda, J.J.; Graham, J.H.; Emlen, J.M.; Krzysik, A.J.; Balbach, H.E.; Kovacic, D.A.; Zak, J.C. Photosynthesis and fluctuating asymmetry as indicators of plant response to soil disturbance in the fall-line sandhills of Georgia: A case study using Rhus copallinum and Ipomoea pandurata. Int. J. Plant Sci. 2004, 165, 805–816. [Google Scholar] [CrossRef]
- Kleinman, J.S.; Hart, J.L. Response by vertical strata to catastrophic wind in restored Pinus palustris stands. J. Torrey Bot. Soc. 2017, 144, 423–438. [Google Scholar] [CrossRef]
- Perry, D.A.; Amaranthus, M.P.; Borchers, J.G.; Borchers, S.L.; Brainerd, R.E. Bootstrapping in ecosystems. Bioscience 1989, 39, 230–237. [Google Scholar] [CrossRef]
- Orman, O.; Adamus, M.; Szewczyk, J. Regeneration processes on coarse woody debris in mixed forests: Do tree germinants and seedlings have species-specific responses when grown on coarse woody debris? J. Ecol. 2016, 104, 1809–1818. [Google Scholar] [CrossRef]
- Kennedy, P.G.; Quinn, T. Understory plant establishment on old-growth stumps and the forest floor in western Washington. For. Ecol. Manag. 2001, 154, 193–200. [Google Scholar] [CrossRef]
- Sleeper, B.E.; Ficklin, R.L. Edaphic and vegetative responses to forested wetland restoration with created microtopography in Arkansas. Ecol. Restor. 2016, 34, 117–123. [Google Scholar] [CrossRef]
- Titus, J.H. Microtopography and woody plant regeneration in a hardwood floodplain swamp in Florida. Bull. Torrey Bot. Club 1990, 117, 429–437. [Google Scholar] [CrossRef]
- Russell, K.N.; Beauchamp, V.B. Plant species diversity in restored and created Delmarva Bay wetlands. Wetlands 2017, 37, 1119–1133. [Google Scholar] [CrossRef]
- Åström, M.; Dynesius, M.; Hylander, K.; Nilsson, C. Effects of slash harvest on bryophytes and vascular plants in southern boreal forest clear-cuts. J. Appl. Ecol. 2005, 42, 1194–1202. [Google Scholar] [CrossRef]
- Goldin, S.R.; Brookhouse, M.T. Effects of coarse woody debris on understorey plants in a temperate Australian woodland. Appl. Veg. Sci. 2015, 18, 134–142. [Google Scholar] [CrossRef]
- Bace, R.; Svoboda, M.; Janda, P. Density and height structure of seedlings in subalpine spruce forests of Central Europe: Logs vs. stumps as a favourable substrate. Silva Fenn. 2011, 45, 1065–1078. [Google Scholar] [CrossRef] [Green Version]
- Holeksa, J.; Saniga, M.; Szwagrzyk, J.; Dziedzic, T.; Ferenc, S.; Wodka, M. Altitudinal variability of stand structure and regeneration in the subalpine spruce forests of the Pol’ana biosphere reserve, Central Slovakia. Eur. J. For. Res. 2007, 126, 303–313. [Google Scholar] [CrossRef]
- Bird, P.R.; Jackson, T.T.; Kearney, G.A.; Roache, A. Effects of windbreak structure on shelter characteristics. Aust. J. Exp. Agric. 2007, 47, 727–737. [Google Scholar] [CrossRef]
- Cleugh, H.A.; Hughes, D.E. Impact of shelter on crop microclimates: A synthesis of results from wind tunnel and field experiments. Aust. J. Exp. Agric. 2002, 42, 679–701. [Google Scholar] [CrossRef]
- Ulyshen, M.D. Saproxylic Insects: Diversity, Ecology and Conservation; Springer International Publishing: New York, NY, USA, 2018. [Google Scholar]
- Davis, J.C.; Castleberry, S.B.; Kilgo, J.C. Influence of coarse woody debris on herpetofaunal communities in upland pine stands of the southeastern Coastal Plain. For. Ecol. Manag. 2010, 259, 1111–1117. [Google Scholar] [CrossRef]
- White, D.L. Land-use history. In Ecology and Management of a Forested Landscape: Fifty Years of Natural Resource Stewardship on the Savannah River Site; Kilgo, J.C., Blake, J.I., Eds.; Island Press: Washington, DC, USA, 2005; pp. 2–12. [Google Scholar]
- Błońska, E.; Kacprzyk, M.; Spolnik, A. Effect of deadwood of different tree species in various stages of decomposition on biochemical soil properties and carbon storage. Ecol. Res. 2017, 32, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Stutz, K.P.; Lang, F. Potentials and unknowns in managing coarse woody debris for soil functioning. Forests 2017, 8, 37. [Google Scholar] [CrossRef]
- Laiho, R.; Prescott, C.E. Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis. Can. J. For. Res. 2004, 34, 763–777. [Google Scholar] [CrossRef]
- Ford, S.A.; Kleinman, J.S.; Hart, J.L. Effects of wind disturbance and salvage harvesting on macrofungal communities in a Pinus woodland. For. Ecol. Manag. 2018, 407, 31–46. [Google Scholar] [CrossRef]
- McIntyre, S.; Cunningham, R.B.; Donnelly, C.F.; Manning, A.D. Restoration of eucalypt grassy woodland: Effects of experimental interventions on ground-layer vegetation. Aust. J. Bot. 2015, 62, 570–579. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Gustafsson, L.; Benayas, J.M.R.; Castro, J. Does post-disturbance salvage logging affect the provision of ecosystem services? A systematic review protocol. Environ. Evid. 2015, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Beschta, R.L.; Rhodes, J.J.; Kauffman, J.B.; Gresswell, R.E.; Minshall, G.W.; Karr, J.R.; Perry, D.A.; Hauer, F.R.; Frissell, C.A. Postfire management on forested public lands of the western United States. Conserv. Biol. 2004, 18, 957–967. [Google Scholar] [CrossRef]
- DellaSala, D.A.; Karr, J.R.; Schoennagel, T.; Perry, D.; Noss, R.F.; Lindenmayer, D.; Beschta, R.; Hutto, R.L.; Swanson, M.E.; Evans, J. Post-fire logging debate ignores many issues. Science 2006, 314, 51–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindenmayer, D.B.; Noss, R.F. Salvage logging, ecosystem processes, and biodiversity conservation. Conserv. Biol. 2006, 20, 949–958. [Google Scholar] [CrossRef]
- Kroll, A.J.; Duke, S.D.; Hane, M.E.; Johnson, J.R.; Rochelle, M.; Betts, M.G.; Arnett, E.B. Landscape composition influences avian colonization of experimentally created snags. Biol. Conserv. 2012, 152, 145–151. [Google Scholar] [CrossRef]
- Rost, J.; Clavero, M.; Brotons, L.; Pons, P. The effect of postfire salvage logging on bird communities in Mediterranean pine forests: The benefits for declining species. J. Appl. Ecol. 2012, 49, 644–651. [Google Scholar] [CrossRef]
- Kleinman, J.S.; Goode, J.D.; Fries, A.C.; Hart, J.L. Ecological consequences of compound disturbances in forest ecosystems: A systematic review. Ecosphere 2019, 10, e02962. [Google Scholar] [CrossRef] [Green Version]
- Trumbore, S.; Brando, P.; Hartmann, H. Forest health and global change. Science 2015, 349, 814–818. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.W.; Jackson, S.T. Novel climates, no-analog communities, and ecological surprises. Front. Ecol. Environ. 2007, 5, 475–482. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Relative Density |
|---|---|
| Vaccinium arboreum Marshall | 29.02 |
| Rhus copallinum L. | 18.39 |
| Pinus palustris Mill. | 9.90 |
| Nyssa sylvatica Marshall | 5.90 |
| Symplocos tinctoria (L.) L’Hér. | 5.69 |
| Oxydendrum arboreum (L.) DC. | 3.21 |
| Quercus falcata Michx. | 3.14 |
| Diospyros virginiana L. | 2.97 |
| Callicarpa americana L. | 2.80 |
| Quercus nigra L. | 2.14 |
| Vaccinium stamineum L. | 1.93 |
| Sassafras albidum (Nutt.) Nees | 1.90 |
| Carya tomentosa (Lam.) Nutt. | 1.59 |
| Quercus coccinea Münchh. | 1.38 |
| Acer rubrum L. | 1.35 |
| Quercus alba L. | 1.07 |
| Quercus margarettae W.W. Ashe ex Small | 1.04 |
| Quercus stellata Wangenh. | 0.83 |
| Carya glabra (Mill.) Sweet | 0.76 |
| Liquidambar styraciflua L. | 0.72 |
| Asimina parviflora (Michx.) Dunal | 0.55 |
| Quercus velutina Lam. | 0.55 |
| Rhododendron canescens (Michx.) Sweet | 0.52 |
| Quercus rubra L. | 0.48 |
| Vaccinium elliottii Chapm. | 0.48 |
| Quercus hemisphaerica Bartram ex Willd. | 0.41 |
| Prunus serotina Ehrh. var. serotina | 0.31 |
| Pinus taeda L. | 0.21 |
| Nestronia umbellula Raf. (V10) | 0.17 |
| Quercus marilandica Münchh. var. marilandica | 0.14 |
| Rhus glabra L. | 0.14 |
| Quercus montana Willd. | 0.10 |
| Cornus florida L. | 0.07 |
| Acer floridanum (Chapm.) Pax | 0.03 |
| Hamamelis virginiana L. | 0.03 |
| Liriodendron tulipifera L. | 0.03 |
| Quercus laevis Walter | 0.03 |
| Total | 100.00 |
| Species | Relative Density |
|---|---|
| Rhus copallinum L. | 26.09 |
| Oxydendrum arboreum (L.) DC. | 16.52 |
| Vaccinium arboreum Marshall | 10.43 |
| Quercus nigra L. | 8.26 |
| Quercus falcata Michx. | 6.09 |
| Quercus coccinea Münchh. | 5.22 |
| Acer rubrum L. | 4.78 |
| Symplocos tinctoria (L.) L’Hér. | 3.48 |
| Nyssa sylvatica Marshall | 3.04 |
| Carya glabra (Mill.) Sweet | 2.61 |
| Diospyros virginiana L. | 2.61 |
| Liquidambar styraciflua L. | 2.17 |
| Carya tomentosa (Lam.) Nutt. | 1.74 |
| Callicarpa americana L. | 1.30 |
| Quercus velutina Lam. | 1.30 |
| Hamamelis virginiana L. | 0.87 |
| Quercus alba L. | 0.87 |
| Quercus marilandica Münchh. var. marilandica | 0.87 |
| Asimina parviflora (Michx.) Dunal | 0.43 |
| Pinus taeda L. | 0.43 |
| Quercus stellata Wangenh. | 0.43 |
| Rhus glabra L. | 0.43 |
| Total | 100.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Logan, A.T.; Goode, J.D.; Keellings, D.J.; Hart, J.L. Microsite Influence on Woody Plant Regeneration in a Pinus palustris Woodland Following Catastrophic Disturbance. Forests 2020, 11, 588. https://doi.org/10.3390/f11050588
Logan AT, Goode JD, Keellings DJ, Hart JL. Microsite Influence on Woody Plant Regeneration in a Pinus palustris Woodland Following Catastrophic Disturbance. Forests. 2020; 11(5):588. https://doi.org/10.3390/f11050588
Chicago/Turabian StyleLogan, Alexandra T., Jonathan D. Goode, David J. Keellings, and Justin L. Hart. 2020. "Microsite Influence on Woody Plant Regeneration in a Pinus palustris Woodland Following Catastrophic Disturbance" Forests 11, no. 5: 588. https://doi.org/10.3390/f11050588