The Economy of Canopy Space Occupation and Shade Production in Early- to Late-Successional Temperate Tree Species and Their Relation to Productivity

Abstract
:1. Introduction
2. Material and Methods
2.1. Tree Species and Study Sites
2.2. Determination of Aboveground Biomass, Biomass per Crown Volume and Biomass Nutrient Content
2.3. The Cost of Canopy Space Occupation and Shade Production
2.4. Determination of Aboveground Productivity
2.5. Statistical Analysis
3. Results
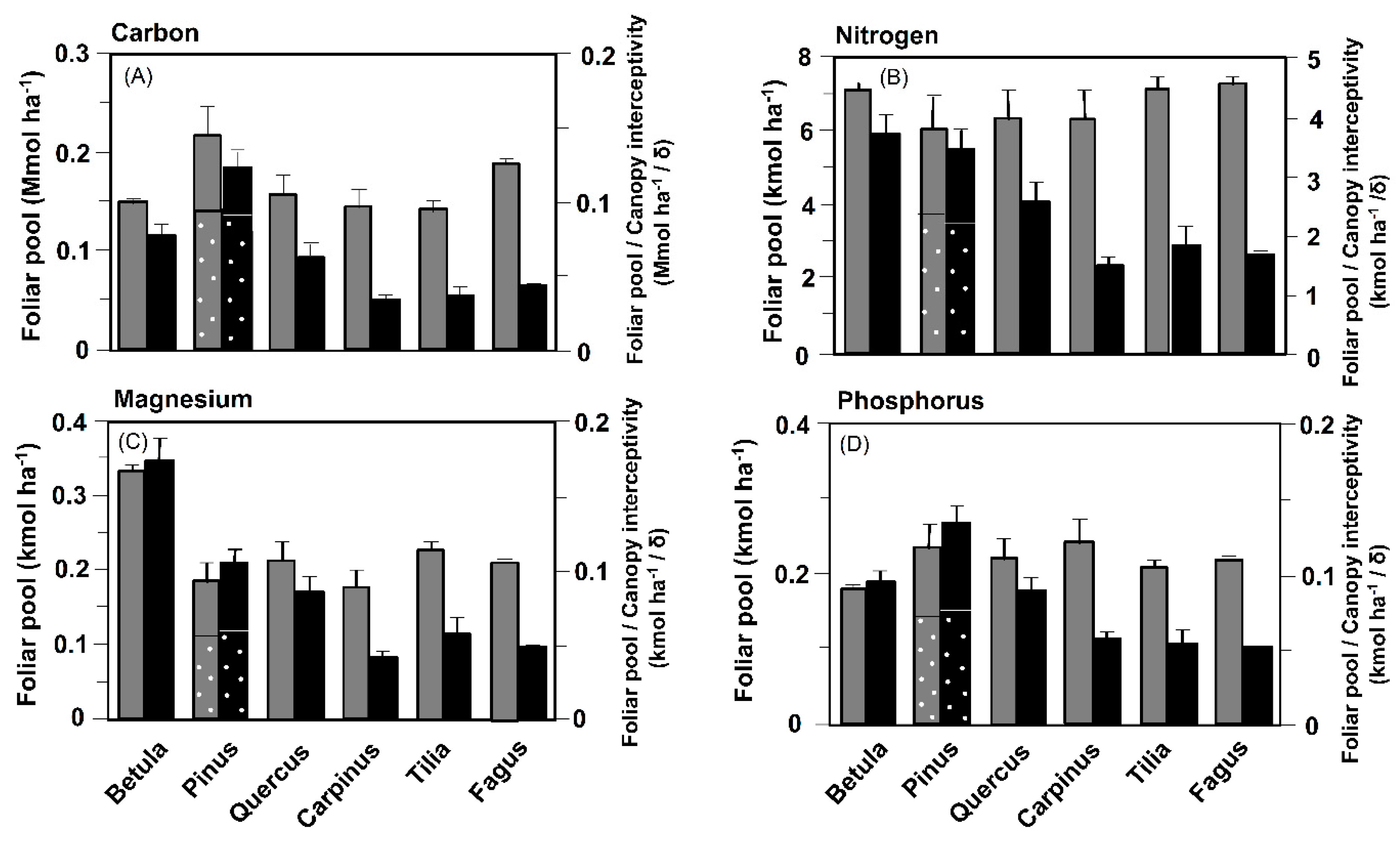
3.1. Aboveground Biomass and Stocks of Carbon and Nutrient Elements in the Crown
3.2. Carbon and Nutrient Costs of Space Occupation and Shade Production
3.3. Aboveground Productivity
4. Discussion
4.1. Do the Costs of Space Occupation and Shade Production Vary with the Species’ seral Status?
4.2. Are Crown Expansion and Shade Production Related to Productivity and Nutrient Acquisition?
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Valladares, F.; Niinemets, Ü. The architecture of plant crowns: From design rules to light capture and performance. In Functional Plant Ecology; Pugnaire, F.I., Valladares, F., Eds.; CRC Press: Boca Raton, London; New York, NY, USA, 2007; pp. 101–149. [Google Scholar]
- Landsberg, J.J. Forest canopies. In Encyclopedia of Environmental Biology; Nierenberg, W.A., Ed.; Academic Press: San Diego, CA, USA, 1995; Volume 3, pp. 81–94. [Google Scholar]
- Bazzaz, F. Plants in Changing Environments; Cambridge Univ. Press: Cambridge, UK, 1996. [Google Scholar]
- Valladares, F. Architecture, ecology, and evolution of plant crowns. In Handbook of Functional Plant Ecology; Pugnaire, F.I., Valladares, F., Eds.; Marcel Dekker: New York, NY, USA, 1999; pp. 121–194. [Google Scholar]
- Grams, T.E.E.; Lüttge, U. Space as a resource. Prog. Bot. 2011, 72, 349–370. [Google Scholar]
- Rudnicki, M.; Lieffers, V.J.; Silins, U. Stand structure governs the crown collisions of lodgepole pine. Can. J. For. Res. 2003, 33, 1238–1244. [Google Scholar] [CrossRef] [Green Version]
- Hajek, P.; Seidel, D.; Leuschner, C. Mechanical abrasion, and not competition for light, is the dominant canopy interaction in a temperate mixed forest. For. Ecol. Manag. 2015, 348, 108–116. [Google Scholar] [CrossRef]
- Niinemets, Ü. Photosynthesis and resource distribution through plant canopies. Plant Cell Environ. 2007, 30, 1052–1071. [Google Scholar] [CrossRef]
- Oldham, A.R.; Sillett, S.C.; Tomescu, A.M.F.; Koch, G.W. The hydrostatic gradient, not light availability, drives height-related variation in Sequoia sempervirens (Cupressaceae) leaf anatomy. Am. J. Bot. 2010, 97, 1087–1097. [Google Scholar] [CrossRef] [Green Version]
- Chin, A.R.O.; Sillett, S.C. Within-crown plasticity in leaf traits among the tallest conifers. Am. J. Bot. 2017, 106, 1–13. [Google Scholar] [CrossRef]
- Reiter, I.M.; Häberle, K.-H.; Nunn, A.J.; Heerdt, C.; Reitmayer, H.; Grote, R.; Matyssek, R. Competitive strategies in adult beech and spruce: Space-related foliar carbon investment versus carbon gain. Oecologia 2005, 146, 337–349. [Google Scholar] [CrossRef]
- Farnsworth, K.D.; Niklas, K.J. Theories of optimization, form and function in branching architecture in plants. Funct. Ecol. 1995, 9, 355–363. [Google Scholar] [CrossRef]
- Chen, S.G.; Ceulemans, R.; Impens, I.A. A fractal-based Populus canopy structure model for the calculation of light interception. For. Ecol. Manag. 1994, 69, 97–110. [Google Scholar] [CrossRef]
- Fleck, S. Integrated Analysis of Relationships between 3D-Structure, Leaf Photosynthesis and Branch Transpiration of Mature Fagus sylvatica and Quercus petraea Trees in a Mixed Forest Stand; Bayreuther Forum für Ökologie 97; University of Bayreuth: Bayreuth, Germany, 2001. [Google Scholar]
- Küppers, M. Kohlenstoffhaushalt, Wasserhaushalt, Wachstum und Wuchsform von Holzgewächsen im Konkurrenzgefüge eines Heckenstandortes. Ph.D. Thesis, University of Bayreuth, Bayreuth, Germany, 1982. [Google Scholar]
- Küppers, M. Carbon relations and competition between woody species in a Central European hedgerow. IV. Growth from and partitioning. Oecologia 1985, 66, 343–352. [Google Scholar] [CrossRef]
- Kozovits, A.R.; Matyssek, R.; Winkler, J.B.; Göttlein, A.; Blaschke, H.; Grams, T.E.E. Aboveground space sequestration determines competitive success in juvenile beech and spruce trees. New Phytol. 2005, 167, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Kramer, R.D.; Sillett, S.C.; van Pelt, R. Quantifying aboveground components of Picea sitchensis for allometric comparisons among tall conifers in North American rainforests. For. Ecol. Manag. 2018, 430, 59–77. [Google Scholar] [CrossRef]
- Sillett, S.C.; van Pelt, R.; Carroll, A.L.; Campbell-Spickler, J. Structure and dynamics of forests dominated by Sequoiadendron giganteum. For. Ecol. Manag. 2019, 448, 218–239. [Google Scholar] [CrossRef]
- Cermak, J.; Riguzzi, F.; Ceulemans, R. Scaling up from the individual tree to the stand level in Scots pine. I. Needle distribution, overall crown and root geometry. Ann. Sci. For. 1997, 55, 63–88. [Google Scholar] [CrossRef] [Green Version]
- Ford, E.D. Branching, crown structure, and the control of timber production. In Attributes of Trees as Crop Plants; Cannell, M.G.R., Jackson, J.E., Eds.; Institute of Terrestrial Ecology: Huntingdon, UK, 1985; pp. 228–252. [Google Scholar]
- Leuschner, C. Changes in forest ecosystem function with succession in the Lüneburger Heide. In Ecosystem Properties and Landscape Function in Central Europe; Tenhunen, J., Hantschel, R., Lenz, R., Eds.; Ecological Studies 147; Springer: Berlin, Germany, 2001; pp. 517–567. [Google Scholar]
- Hallé, F.; Oldeman, R.A.A.; Tomlinson, P.B. Tropical Trees and Forests. An Architectural Analysis; Springer: Heidelberg/Berlin, Germany, 1978. [Google Scholar]
- Turner, I.M. The Ecology of Trees in the Tropical Rain Forest; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Bazzaz, F. The physiological ecology of plant succession. Ann. Rev. Ecol. Syst. 1979, 10, 351–371. [Google Scholar] [CrossRef] [Green Version]
- Huston, M.; Smith, T. Plant succession, life history and competition. Am. Nat. 1987, 130, 168–198. [Google Scholar] [CrossRef]
- Küppers, M. Canopy gaps: Competitive light interception and economic space filling–A matter of whole plant allocation. In Exploitation of Environmental Heterogeneity by Plant Ecophysiological Processes Above and Below Ground; Caldwell, M., Pearcy, R.W., Eds.; Academic Press: San Diego, CA, USA, 1994; pp. 111–114. [Google Scholar]
- Franklin, J.F.; Spies, T.A.; van Pelt, R.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.S. Disturbance and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Grace, J.B.; Tilman, D. Perspectives on Plant Competition; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Hagemeier, M.; Leuschner, C. Functional crown architecture of five temperate broadleaf tree species: Vertical gradients in leaf morphology, leaf angle, and leaf area density. Forests 2019, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Stoutjesdijk, P.; Barkman, J.J. Microclimate, Vegetation and Fauna; Opulus Press: Uppsala, Sweden, 1987. [Google Scholar]
- Hagemeier, M.; Leuschner, C. Leaf and crown optical properties of five early-, mid- and late-successional temperate tree species and their relation to sapling light demand. Forests 2019, 10, 925. [Google Scholar] [CrossRef] [Green Version]
- Ellenberg, H. Vegetation Mitteleuropas mit den Alpen, 6th ed.; Ulmer Verlag: Stuttgart, Germany, 1996. [Google Scholar]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests. In Vegetation Ecology of Central Europe; Springer Nature: Cham, Switzerland, 2017; Volume 1. [Google Scholar]
- Leuschner, C.; Meier, I.C. The ecology of Central European tree species: Trait spectra, functional trade-offs, and ecological classification of adult trees. Perspec. Plant Ecol. Evol. Syst. 2018, 33, 89–103. [Google Scholar] [CrossRef]
- Schober, R. Ertragstafeln wichtiger Baumarten; Sauerländer, J.D., Frankfurt, M., Eds.; Germany, 1995; Available online: https://www.lehmanns.de/shop/weitere-fachgebiete/5012551-9783793907305-ertragstafeln-wichtiger-baumarten-bei-verschiedener-durchforstung (accessed on 11 March 2020).
- Böckmann, F. Wachstum und Ertrag der Winterlinde (Tilia cordata Mill.) in Niedersachsen und Nordhessen. Ph.D. Thesis, University of Göttingen, Göttingen, Germany, 1990. [Google Scholar]
- Roloff, A. Kronenentwicklung und Vitalitätsbeurteilung ausgewählter Baumarten der gemäßigten Breiten. Schr. Forstl. Fak. Univ. Göttingen Niedersächs. Forstl. Vers. anst. 1989, 93, 1–258. [Google Scholar]
- Lovy, D. WinDIG. University of Geneva, Switzerland. 1996. Available online: http://www.unige.ch/sciences/chifi/cpb/windig.html (accessed on 1 February 2002).
- Ceulemans, R.; Saugier, B. Photosynthesis. In Physiology of Trees; Raghavendra, A.S., Ed.; Wiley: New York, NY, USA, 1991; pp. 21–50. [Google Scholar]
- Kutsch, W.; Wirth, C.; Kattge, J.; Nöllert, S.; Herbst, M.; Kappen, L. Ecophysiological characteristics of mature trees and stands–Consequences for old-growth forest productivity. In Old-Growth Forests. Function, Fate and Value; Wirth, C., Gleixner, G., Heimann, M., Eds.; Ecol. Stud. 127; Springer: Berlin/Heidelberg, Germany, 2009; pp. 57–80. [Google Scholar]
- Art, H.W.; Marks, P.L. A summary table of biomass and net annual primary production in forest ecosystems of the world. In Proceedings of the XVth IUFRO Congress, Gainsville, Florida, USA, 15–20 March 1971; Working Group on Forest Biomass Studies, Section 25, Growth and Yield. University of Florida: Gainsville, FL, USA; IUFRO: Vienna, Austria; pp. 3–32. [Google Scholar]
- De Angeles, D.L.; Gardner, R.H.; Shugart, H.H. Productivity of forest ecosystems studied during the IPB: The woodland data set. In Dynamic Properties of Forest Ecosystems; Reichle, D.E., Ed.; Cambridge University Press: Cambridge, UK; London, UK; New York, NY, USA; Melbourne, Australia, 1981; pp. 567–672. [Google Scholar]
- Cannell, M.G.R. World Forest Biomass and Production Data; Academic Press: London, UK, 1982. [Google Scholar]
- Scarascia-Mugnozza, G.; Bauer, G.A.; Persson, H.; Matteucci, G.; Masci, A. Tree biomass, growth and nutrient pools. In Carbon and Nitrogen Cycling in European Forest Ecosystems; Schulze, E.D., Ed.; Ecol. Stud. 142; Springer: Berlin, Germany, 2000; pp. 49–62. [Google Scholar]
- Ellenberg, H.; Mayer, R.; Schauermann, J. Ökosystemforschung–Ergebnisse des Sollingprojekts 1966–1986; Ulmer: Stuttgart, Germany, 1986. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Betula pendula | Pinus sylvestris | Quercus petraea | Carpinus betulus | Tilia cordata | Fagus sylvatica | |
|---|---|---|---|---|---|---|
| Family | Betulaceae | Pinaceae | Fagaceae | Betulaceae | Malvaceae | Fagaceae |
| Successional status | early | early | mid/late | mid/late | late | late |
| Maximum age (yr) | 120 | 450 | 500 | 250 | 400 | 450 |
| Age of peak volume increment 1 | 65 | 75 | 125 | ? | 65 | 145 |
| Maximum height (m) | 35 | 50 | 50 | 30 | 45 | 56 |
| Wood density (g cm−3) | 610 | 490 | 650 | 790 | 490 | 680 |
| Crown architectural type 2 | Ra/Sc/Tr | Rauh | Rauh | Troll | Troll | Troll |
| Shade production of adults 3 | V | V | III | I | II | I |
| Light demand of saplings 4 | 21 | 18 | 11 | 3 | 3.5 | 3.5 |
| Light demand of shade leaves 5 | 21 | 25 | 6 | 2.5 | 3 | <1 |
| Species | Site # | Location | Coordi-Nates | Slope | Elevation m a.s.l. | Soil Substrate | Admixed Species |
|---|---|---|---|---|---|---|---|
| Betula pendula | 1 | Lehrer Wald (Helmstedt county), Lower Saxony | 10°42’E 52°19’N | level | 95 | Loamy sand | Alnus glutinosa |
| 2 | Lappwald (Helmstedt county), Lower Saxony | 11°01’E 52°16’N | level | 147 | Loamy sand | Pinus sylvestris | |
| Pinus sylvestris | 3 | Unterlüss, Lüneburger Heide, Lower Saxony | 52°51’N 10°16’E | level | 103 | Loamy sand | Betula pendula |
| 4 | Unterlüss, Lüneburger Heide, Lower Saxony | 52°50’N 10°15’E | level | 112 | Loamy sand | Betula pend., Picea abies | |
| Quercus petraea | 5 | Breitenhees, Lüneburg Heath, Lower Saxony | 10°30’E 52°49’N | level | 120 | Loamy sand | Picea abies |
| 6 | Breitenhees, Lüneburg Heath, Lower Saxony | 10°30’E 52°49’N | level | 123 | Loamy sand | Picea abies | |
| Carpinus betulus | 7 | Ziegelrodaer Forst (near Mansfeld), Thuringia | 11°32’E 51°26’N | 5% NW | 225 | Sandy loam | Tilia cordata, Acer ssp. |
| 8 | Ziegelrodaer Forst (near Mansfeld), Thuringia | 11°32’E 51°26’N | 11% N | 235 | Sandy loam | Fraxinus exc. Tilia cordata | |
| Tilia cordata | 9 | Colbitz-Letzlinger Heide (Wolmirstedt), Sax.-Anhalt | 11°32’E 52°21’N | level | 85 | Loamy sand | Quercus rob., Fraxinus exc. |
| 10 | Colbitz-Letzlinger Heide (Wolmirstedt), Sax.-Anhalt | 11°33’E 52°20’N | level | 78 | Loamy sand | Quercus rob., Fraxinus exc. | |
| Fagus sylvatica | 11 | Unterlüss, Lüneburger Heide, Lower Saxony | 10°16’E 52°50’N | level | 115 | Sand | Quercus petraea |
| 12 | Unterlüss, Lüneburger Heide, Lower Saxony | 10°19’E 52°49’N | level | 113 | Loamy sand | - |
| Crown Volume Occupation | Shade Production | |||||
|---|---|---|---|---|---|---|
| Initial C costs | Annual C costs | Initial N costs | Annual N costs | C costs | N costs | |
| r | 0.77 | 0.75 | 0.76 | 0.48 | 0.40 | 0.07 |
| p | 0.04 | 0.04 | 0.04 | 0.16 | 0.21 | 0.44 |
| Tree species | Age | Basal area | ANPP | Source |
|---|---|---|---|---|
| years | m2 ha−1 | Mg ha−1 year−1 | ||
| Betula pendula | 42–55 | 21.6–26 | 7.7–10.8 | [42,43,44] |
| 45–67 | 25.2–28.5 | 9.2–10.2 | This study | |
| Pinus sylvestris | 35–55 | 30.8–35 | 7.9–22.0 | [42,43,44] |
| 48–50 | 29.5–33.4 | 14.5–8.1 | This study | |
| 65 ± 6 | 28.1 ± 4.8 | 6.8 | Förster et al., submitted | |
| Quercus petraea | 117 | - | 12.6 | [42,43,44] |
| 149-154 | 23.8–27.5 | 11.4–11.8 | This study | |
| Carpinus betulus | 95–116 | 31.5–37 | 15.7–16.1 | This study |
| Tilia cordata | 74 | - | 7.0 | [42,43,44] |
| 65–83 | 35.9–37.7 | 8.2–8.7 | This study | |
| Fagus sylvatica | 80–122 | 25.2–31 | 10.1–17.3 | [42,43,44] |
| 95–115 | 24.2–30.8 | 10.1–12.4 | This study | |
| 146 ± 31 | 29.1 ± 6.4 | 9.0 | Förster et al., submitted | |
| 118 | 24.6 | 8.9 | [45] | |
| 71–134 | 32–40 | 9.4–10.9 | [46] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leuschner, C.; Hagemeier, M. The Economy of Canopy Space Occupation and Shade Production in Early- to Late-Successional Temperate Tree Species and Their Relation to Productivity. Forests 2020, 11, 317. https://doi.org/10.3390/f11030317
Leuschner C, Hagemeier M. The Economy of Canopy Space Occupation and Shade Production in Early- to Late-Successional Temperate Tree Species and Their Relation to Productivity. Forests. 2020; 11(3):317. https://doi.org/10.3390/f11030317
Chicago/Turabian StyleLeuschner, Christoph, and Marc Hagemeier. 2020. "The Economy of Canopy Space Occupation and Shade Production in Early- to Late-Successional Temperate Tree Species and Their Relation to Productivity" Forests 11, no. 3: 317. https://doi.org/10.3390/f11030317